







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
塔里木盆地苏盖特布拉克剖面寒武系第二统第三阶微生物岩结构类型*
[李莹1, 2, 3  , 潘文庆
, 潘文庆4 , 吴亚生1, 2, 3 , 杨果4 , 孙崇浩4 , 姜红霞5 ]
, 潘文庆, 杨果|
|

第一作者简介 李莹,女,1997年生,硕士研究生,研究方向为生物沉积学。E-mail: lilynut@163.com。
塔里木盆地阿克苏地区寒武系第二统第三阶肖尔布拉克组主要由微生物白云岩组成,代表性的剖面在阿克苏市西南90 km处的苏盖特布拉克。由于成岩作用改造严重,对这套微生物碳酸盐岩的特征和形成环境的认识还存在很多分歧,制约了勘探工作。通过详细野外勘察和室内研究,将肖尔布拉克组微生物岩分为 4种结构类型: 凝块结构、纹层结构、砂屑结构和骨架结构。首次对凝块结构和纹层结构进行了亚类型的划分,其中凝块结构可划分为蠕虫状、网状和斑点状 3种亚类型,纹层结构可划分为致密纹层、短薄纹层、颗粒纹层以及单纹层、纹层组、复合纹层。肖 Ⅰ段—肖 Ⅲ段发育凝块石白云岩、凝块—层纹石白云岩和层纹石白云岩,肖 Ⅳ段发育凝块石微生物丘,肖 V段下部发育网状结构凝块石白云岩,肖 V段上部发育砂屑白云岩和肾形菌骨架岩。白云石化作用、溶蚀作用、重结晶作用是改变肖尔布拉克组微生物岩结构最重要的成岩作用,且该组下部比上部遭受了更强的成岩作用改造;微生物岩结构对成岩作用改造的抵抗能力为: 砂屑结构>纹层结构>凝块结构。根据微生物岩结构,推测肖 Ⅰ段—肖 Ⅲ段形成于潮坪环境,肖 Ⅳ段和肖 V段下部形成于深水潮下环境,肖 V段上部形成于浅水潮下环境。以上成果为认识塔里木盆地肖尔布拉克组微生物岩的平面分布规律和今后开展被成岩作用强烈改造的白云岩型微生物岩的研究提供了一个重要参考。
About the first author Li Ying,born in 1997,a master degree candidate,is engaged in research of biosedimentology. E-mail: lilynut@163.com.
The Xiaoerbulake Formation(Stage 3, Cambrian Series 2)in the Aksu area of the Tarim Basin is mainly composed of microbial dolostone with its reprehensive section at Sugaitebulake,90 km southwest of Aksu city. This formation can be ideal petroleum reservoirs for its good pores which has been extensively studied. Due to serious diagenesis alteration,however, their features have not been fully identified,which causes difficulty in understanding their formative processes and hampers the petroleum exploration. Here,based on detailed observation on outcrops and thin sections,we identified four types of microbialite fabrics,including thrombolitic fabric,laminated fabric,intraclastic fabric and framestone fabric,and established high-precision vertical evolutionary sequence of microbialites. The thrombolitic and the laminated fabrics can be further subdivided. The Members Ⅰ to Ⅲ are mainly composed of thrombolitic dolostone,thrombolitic-laminated dolostone and laminated dolostone. The Member IV consists of microbial thrombolite mounds,whereas the lower Member V is net-thrombolitic dolostone and the upper Member V intraclastic dolostone and Renalcis framestone. This study reveals that the original fabrics have been affected by dolomitization,recrystallization and dissolution,and the alteration degree of the microbialite fabric is more severe in the lower parts of this formation. The intraclastic fabric has the strongest resistance to diagenesis,followed by the laminated fabric,and then the thrombolitic fabric. The Members Ⅰ to Ⅲ represent tidal flat environments;the Member Ⅳ and the lower Member Ⅴ represent deep subtidal environments;the upper Member Ⅴ represents shallower subtidal environments.
微生物岩(microbiolite)一词最早由Burne和Moore(1987)提出, 后由Riding(1991)改为microbialite, 指底栖微生物群落生长发育过程中通过微生物捕获、粘结碎屑颗粒和(或)有机生物矿化作用及无机矿化作用形成的生物沉积物, 其中参与造岩的微生物包括细菌(主要是蓝细菌)、古菌、微体藻类及原生动物等。微生物岩以微生物碳酸盐岩为主要岩石类型, 在地质历史中分布广泛。按照中观结构, 微生物碳酸盐岩可分为叠层石、凝块石、树形石、均一石、核形石、层纹石等类型(Aitken, 1967; Riding, 1991; Braga et al., 1995; 梅冥相, 2007; 吴亚生等, 2018)。结构是微生物岩研究的基础内容, 不同的结构类型反映不同的形成环境, 因此微生物碳酸盐岩结构的研究对于揭示其形成机制和形成环境、预测其时空分布有重要意义。
寒武纪是微生物碳酸盐岩发育的一个重要时期。主流观点认为寒武纪初期大气CO2浓度的降低促进了蓝细菌钙化机制(CCMs)的发生, 钙化微生物化石开始广泛出现在纹层状的叠层石及非纹层状的凝块石和树形石中(Riding, 2012)。随着寒武纪生命大爆发及后生动物的辐射, 古杯类动物— 钙化微生物礁开始大量出现, 单一微生物岩数量减少。寒武纪第二世末, 后生动物开始灭绝, 微生物岩开始恢复繁盛, 并持续至早奥陶世(Wood, 1998; Kiessling, 2009)。华北地台在寒武纪第二世末开始接受沉积, 微生物岩主要发育于苗岭统和芙蓉统, 其中苗岭统微生物岩主要以凝块石和树形石为主, 附枝菌(Epiphyton)是主要造岩微生物, 而芙蓉统微生物岩主要为柱状叠层石, 几乎不含附枝菌(Lee et al., 2010, 2014)。华南地区寒武纪微生物岩、古杯— 微生物礁也在第二世开始出现(王剑等, 1990; 王建波等, 2014), 但目前关于华南寒武纪微生物岩的研究开展较少, 尚无芙蓉统微生物礁(岩)的报道。新疆塔里木盆地寒武纪微生物碳酸盐岩发育广泛, 其与华北和华南地区的微生物岩有明显不同, 即一般都为白云岩, 罕见钙质微生物化石, 主要见于寒武系第二统第三阶肖尔布拉克组、第三统第五阶沙依里克组和第三统古丈阶下丘里塔格组(黄智斌等, 2017)。柯坪地区的肖尔布拉克组露头良好, 前人已对该组微生物碳酸盐岩进行了大量的研究(宋金民等, 2012, 2014; 胡文瑄等, 2014; 李保华等, 2015; 熊益学等, 2015; 王凯等, 2016; 白莹等, 2018; 邓世彪等, 2018; 宋亚芳等, 2019; 郑剑锋等, 2019), 但由于成岩改造强烈、微生物岩原始特征破坏严重, 导致研究难度大、认识分歧多、沉积相模式准确性不高, 严重制约了该地区微生物碳酸盐岩储集层内的油气勘探(郑剑锋等, 2019)。
越来越多的研究表明, 微生物碳酸盐岩是一类良好的油气储集层, 具有广阔的勘探前景, 目前已知的如美国的阿拉巴马州阿普尔顿油田、巴西桑托斯盆地和中国渤海湾盆地任丘油田都是储量很大的微生物碳酸盐岩型油田(戴永定和陈孟莪, 1996; 罗平等, 2013)。随着中深1井肖尔布拉克组试油的重大突破(折合日产天然气3× 104 m3、水34 m3)(王招明等, 2014), 塔里木盆地下寒武统肖尔布拉克组逐渐成为油气勘探的重要目标层段。因此, 对塔里木盆地微生物岩的研究不仅具有科学意义, 还具有十分重要的经济价值。文中通过野外剖面勘察和室内薄片观察, 对阿克苏地区苏盖特布拉克剖面肖尔布拉克组微生物碳酸盐岩开展了详细研究, 着重划分和描述微生物碳酸盐岩的结构类型, 探讨其形成环境, 以期为认识该地区沉积相平面分布规律、指导油气勘探开发提供基础依据。
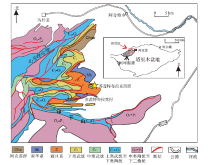
研究区位于塔里木盆地西北缘, 构造上属于塔北隆起柯坪断隆的东段, 距阿克苏市区约90 km(图 1)。新元古代末期, 古塔里木板块受一系列拉张运动的影响开始发生分裂, 依次经历裂陷、拗陷、滨浅海碳酸盐岩沉积阶段, 于早寒武世形成3个孤立碳酸盐岩台地, 分别为塔西、罗西和库鲁克塔格台地, 研究区位于面积最大的塔西台地的西北缘(赵宗举等, 2011; 严威等, 2018; 郑剑锋等, 2019)。塔西台地的寒武系为厚达1000 m的碳酸盐沉积, 现几乎全部由白云岩组成。按照最新的地层划分方案(黄智斌等, 2017), 研究区的寒武系自下而上划分为纽芬兰统玉尔吐斯组、第二统肖尔布拉克组和吾松格尔组、第三统下部的沙依里克组和阿瓦塔格组、第三统上部和芙蓉统下部的下丘里塔格组、芙蓉统上部的蓬莱坝组。
 | 图 1 塔里木盆地西北缘地质简图及苏盖特布拉克剖面位置(据李朋威等, 2015, 修改)Fig.1 Geological map of the northwestern margin of Tarim Basin and location of Sugaitebulake section (modified from Li et al., 2015) |
玉尔吐斯组为一套厚1~20 m的硅质岩— 黑色页岩— 泥质碳酸盐岩, 是一套良好的烃源岩, 整体为台地间的盆地沉积。后期碳酸盐岩台地不断向西北方向发生进积, 至肖尔布拉克组沉积时期, 逐渐演变为局限台地— 开阔台地。肖尔布拉克组整体为一套100多米厚的白云岩地层, 可分为2段(赵宗举等, 2010):下段主要为深灰色薄层泥粉晶白云岩, 称“ 肖下段” , 岩石整体较为致密, 厚约93 m; 上段颜色浅, 为灰白色薄— 中层状或丘状白云岩, 孔隙更为发育, 称“ 肖上段” , 厚约94 m, 是储集层主要发育段。
野外观察和室内薄片研究表明, 肖尔布拉克组几乎全部由白云岩组成, 只在“ 肖下段” 的顶部含有少量灰质白云岩。宏观上, 肖下段主要是薄层状的微生物岩, 肖上段下部是中层状微生物岩夹一些微生物丘, 肖上段上部是中层状微生物岩。微生物岩类型主要为凝块石和层纹石, 有少量微生物骨架岩和砂屑白云岩。按岩性将肖下段进一步分为肖Ⅰ 段、肖Ⅱ 段、肖Ⅲ 段, 将肖上段进一步分为肖Ⅳ 段和肖V段(图 2)。
 | 图 2 塔里木盆地苏盖特布拉克剖面肖尔布拉克组岩性柱状图Fig.2 Stratigraphic histogram of the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin, China |
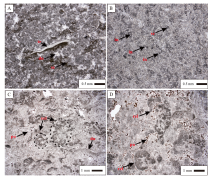
肖 Ⅰ 段: 为黑色薄板状粉细晶— 粗晶白云岩, 厚35 m, 单层厚2~10 cm。底部发育鸟眼构造和窗格构造(图 3-A, 3-B), 前者呈扁豆形, 大小为(0.2~1.5)mm× (0.2~0.5) mm, 呈孤立分散状, 分布不均匀, 一般顺层分布; 后者呈拉长的不规则状, 顺层延伸, 断续状, 宽度一般小于0.5 mm, 长度变化较大。窗格孔内部为细晶、中晶白云石, 粗亮晶孔洞内可见漂浮状细中晶白云岩晶体。
 | 图 3 塔里木盆地苏盖特布拉克剖面寒武系第二统肖尔布拉克组肖下段岩石露头及薄片特征 A— 肖Ⅰ 段下部, 鸟眼构造, 单偏光; B— 肖Ⅰ 段下部, 窗格构造, 单偏光; C— 肖Ⅱ 段, 薄板状层纹石白云岩; D— 肖Ⅱ 段, 黑色蜂窝状白云岩; E— 肖Ⅱ 段顶部, 凝块石白云岩; F— 肖Ⅲ 段, 饼干状极薄层泥质白云岩Fig.3 Outcrop and thin section features of the Lower Member of Xiaoerbulake Formation (Cambrian Series 2) of Sugaitebulake section, Tarim Basin |
肖Ⅱ 段: 厚约38 m, 呈似薄层— 块状, 层理不明显, 风化面呈褐色, 新鲜面黑色, 为黑色蜂窝状细粉晶— 中晶白云岩(图 3-D)与无溶孔细晶白云岩的互层。蜂窝状白云岩中的溶孔面积含量为30%~50%, 呈压扁的近球形、姜状, 既有全部被方解石充填的, 也有部分被充填的, 直径介于0.5~2 cm之间, 平均为0.8 cm, 间距2~10 cm, 顺层分布。该段底部局部发育少量层纹石, 向上层纹石增多(图 3-C), 亦有凝块石(图 3-E)。
肖Ⅲ 段: 为灰色极薄层泥质粉晶白云岩(图 3-F), 厚约20 m, 发育平直层理。单层厚度平均约0.2 cm, 溶孔不发育。镜下见层纹石结构十分发育。
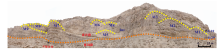
肖Ⅳ 段: 厚约25 m, 在露头上呈灰黄色, 由微生物丘和丘间的层状微生物岩组成。在500 m宽的范围内共识别出9个微生物丘体(图 4), 规模大小不一。以编号为M5的微生物丘为例, 其呈典型的丘状, 高6 m、宽10 m, 内部无层理, 主要由泥粉晶和细晶白云石组成, 具有凝块石的结构特征。微生物丘之间为黑色薄层状凝块石泥晶白云岩, 显微结构与丘体相同。
 | 图 4 塔里木盆地苏盖特布拉克剖面肖尔布拉克组肖Ⅳ 段微生物丘与丘间的层状微生物白云岩 橙色虚线之上为肖Ⅳ 段, 对应薄片S-29至S-39, 之下为肖Ⅲ 段; 微生物丘(M)呈灰白色, 具凝块石的结构, 丘体大小不一, 宽度大于10 m, 高度大于5 m, 丘间(Int)为黑色薄板状凝块石白云岩Fig.4 Outcrops of microbialite dolostone of the Member Ⅳ of Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
肖V段: 厚约69 m, 露头上呈灰黄色, 主要为中— 厚层状白云岩。可分为3段:下段为泥粉晶凝块石白云岩; 中段为肾形藻骨架岩白云岩和团块状凝块石白云岩; 上段主要为粒泥状白云岩、泥粒状白云岩及颗粒状白云岩, 有少数三叶虫化石。
微生物岩的结构是指其矿物的大小、形状及排列方式, 该结构特征可能与其形成环境的水动力条件、沉积速率、微生物群落组成、海水温度和饱和度等因素有关(Turner et al., 1997)。微生物岩的结构可以在不同尺度上进行观察、描述和分类, 目前使用较广的是Shapiro和Awramik(2000)中观结构(凝块状、纹层状等)与微观结构(钙化微生物、胶结物、球粒等)的二级划分, 前者是肉眼可见的厘米级特征, 后者是薄片上才能观察到的毫米级特征。Riding(2011)把微生物岩分为树枝石、叠层石、凝块石和均一石, 这也属于中观结构的划分。
微观结构的研究, 对探究微生物岩形成机制和形成环境具有重要意义。纹层结构和凝块结构是最常见的2种微观结构类型, 此外还有骨架结构、粘结结构等(吴亚生等, 2018)。根据薄片观察, 研究区肖尔布拉克组主要发育凝块和纹层结构, 其次发育骨架和砂屑结构, 其中凝块结构和纹层结构是最常见和最富于变化的结构。基于前人对阿克苏地区寒武系第二统微生物岩的研究(宋金民等, 2012; 黄擎宇等, 2016; 余浩元等, 2018)主要是中观尺度的, 此次研究着重在微观尺度上对微生物岩的结构进行描述和分类, 并将凝块结构进行进一步划分为蠕虫状、网状、斑点状和团块状4种亚类型, 将纹层状结构进一步划分为单纹层、纹层组和复合型3种亚类型。
凝块(clot)是凝块石的基本组成单元, 呈暗色, 被认为是微生物钙化或者捕集泥晶而形成的, 其由泥晶组成, 构成岩石的骨架。凝块的边缘明确, 形状不规则, 大小不一, 从几个毫米到小于1 mm。凝块结构是本组微生物岩的主要结构类型之一。此次统计表明, 凝块结构在每段皆有分布, 面积含量大于60%。
关于凝块石的结构分类, 前人已做过一定的研究, 但大部分是对中观尺度的分类。例如, 杜汝霖(1992)依据凝块石外部形态将其划分为多孔状凝块石、凝块状凝块石和网状凝块石; Riding(2000)从成因角度将凝块石分为钙化微生物凝块石和粗粒的粘合状凝块石等7大类; Harwood和Sumner(2016)将美国大盆地的寒武纪凝块石分为灌木状、半球微指状、合并柱状、窗孔块状、虫孔灌木状、斑点状等8种类型。
镜下研究发现, 肖尔布拉克组凝块石中的凝块主要由暗色泥晶白云石或泥粉晶白云石组成, 形状变化较大, 呈蠕虫状、椭圆状和不规则状孤立分布, 亦可相互连通呈网状。凝块之间是细晶、中晶或粗晶白云石组成的浅色区域, 称为亮斑, 由于边缘较为圆滑, 不似溶蚀作用形成的切割港湾状, 因此推测为原始凝块之间的原生孔。这种原生孔一般有2种来源:一种为菌藻生物体死亡腐烂分解后留下的空间, 一般体积很小, 需要在镜下才能识别; 另一种为凝块之间的剩余空隙。根据镜下凝块的形态特征, 笔者对凝块结构做了进一步划分, 将其分为蠕虫状、斑点状、网状和团块状4种亚类型。
4.1.1 蠕虫状结构
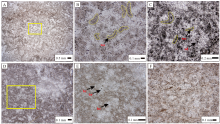
凝块呈蠕虫状, 由黑色泥晶或粉晶白云石组成, 形态呈不规则的窄条状, 构成封闭的网状, 一般或多或少弯曲(图 5-A, 5-B, 5-C)。文中对凝块的形状特征用长度、宽度、弯曲夹角3个参数进行约束。长度是把曲线划分为直线段, 测量直线段的长度, 然后相加获得近似的总长度。苏盖特布拉克剖面肖尔布拉克组凝块石中蠕虫状凝块的长度一般在0.1~0.8 mm之间, 宽度一般在0.1~0.15 mm之间, 弯曲夹角一般在120° ~150° 之间, 弯曲方向不固定。蠕虫状结构在凝块石中分布较均匀, 呈孤立状, 或点接触, 平均间距在0.5 mm以下。蠕虫状凝块之间的浅色亮斑由中晶或粗晶白云石组成。泥晶与中粗晶白云石比例约为1︰1。受重结晶作用影响, 部分黑色泥晶变为黄褐色粉晶, 从而使蠕虫结构边缘模糊并与周围亮斑难以区分。此次研究统计了各段不同结构的薄片数占该段总薄片数的百分含量(图 6), 结果反映蠕虫状结构在全组发育较为广泛, 除肖Ⅱ 段外, 其余段内皆有发育, 以肖Ⅳ 段生物丘内最为丰富, 含量可达70%, 其次为肖Ⅰ 段。该结构常与斑点状结构、网状结构和纹层结构组合出现。
 | 图 5 塔里木盆地苏盖特布拉克剖面肖尔布拉克组蠕虫状结构和网状结构显微特征 A— 肖Ⅲ 段顶部, 蠕虫状结构; B— A中黄框放大5倍, 肖Ⅲ 段顶部, 粉晶白云石组成的蠕虫状结构(wo), 相互不接触, 细晶白云石胶结; C— 肖Ⅳ 段凝块石丘内, 斑点状结构与蠕虫状结构, 凝块之间的亮斑由细晶白云石组成; D— 肖Ⅴ 段顶部, 网状结构; E— 肖Ⅴ 段顶部, D中黄框放大5倍, 网眼(br)为椭圆形— 近圆形或不规则形, 大小不一, 被细晶白云石充填; F— 肖Ⅰ 段中部, 细晶白云石组成的网状结构。所有照片均为单偏光Fig.5 Microscopic features of vermicular and reticular fabrics of the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
 | 图 6 塔里木盆地苏盖特布拉克剖面肖尔布拉克组微生物岩不同结构类型的地层分布和相对含量Fig.6 Distribution and relative content diagram of microbialite fabrics of the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
4.1.2 网状结构
指由暗色泥晶和粉晶白云石构成的不规则网形的结构(图 5-D, 5-E, 5-F)。网状结构是肖尔布拉克组凝块石的主要结构类型, 由网线和网眼两部分构成。网线由泥粉晶白云石组成, 宽度约0.01 mm。网眼则由细晶、中晶、粗晶或亮晶白云石组成, 形状不规则, 直径在0.01~0.5 mm之间。该结构主要见于肖Ⅱ 段、肖Ⅳ 段及肖Ⅴ 段下部。肖Ⅱ 段主要为细晶白云石构成的网状结构, 网眼由中晶白云石组成, 直径在0.05~0.13 mm之间, 平均直径为0.1 mm, 构成岩石骨架; 肖Ⅳ 段及肖V段底部的网状结构网线主要由泥粉晶白云石组成, 网眼由细晶白云石组成, 平均直径为0.05 mm, 并常与斑点状结构和蠕虫状结构组合出现; 肖V段中部的网状结构主要存在于纹层结构的亮层中, 直径介于0.1~0.5 mm之间, 平均0.25 mm, 形态多样, 但含量有限, 统计得出的面积含量约为20%~30%, 且向上含量逐渐降低。
4.1.3 斑点状结构
在镜下表现为由黑色泥晶或粉晶白云石组成的近圆形斑点(图 7-A, 7-B), 直径约0.01 mm, 分布不均匀, 呈分散孤立状分布, 平均间距大于0.05 mm; 或聚集在一起呈点簇状, 直径约0.05 mm; 或大面积弥散为不规则斑状凝块, 边缘不明显, 其间一般是重结晶形成的粉细晶— 中晶白云石。研究区肖尔布拉克组的斑点状结构可能有2种成因, 一是微生物体新陈代谢活动所引起的碳酸盐沉淀(Riding, 2000), 二是蠕虫状结构、网状结构等被重结晶作用改造破坏后的残余。在后一种情况下, 点状结构周围一般为细晶— 中晶白云石, 晶体上可见原结构留下的微小包裹体。斑点状结构主要分布于肖Ⅰ 段— 肖Ⅳ 段, 肖Ⅲ 段和肖Ⅳ 段微生物丘是斑点状结构的主要发育层段, 但因体积小且成岩作用破坏强烈, 目前所见仅约10%。就肖尔布拉克组而言, 该结构不是微生物岩的主要结构类型, 常与蠕虫状结构、网状结构及纹层结构组合出现。
 | 图 7 塔里木盆地苏盖特布拉克剖面肖尔布拉克组斑点状结构和团块状结构显微特征 A— 肖Ⅰ 段, 斑点状结构(sd)及其构成的点簇状结构(clu), 簇状体直径0.1~0.25 mm; B— 肖Ⅲ 段, 纹层结构内的粉晶白云石组成的点簇状结构(clu); C— 肖V段中部, 不规则团块状骨架, 发生重结晶变成粉晶(th), 后被打碎成砾屑, 砾屑之间为泥晶白云石(mi), 后泥晶白云石被溶蚀, 溶蚀孔洞被巨晶白云石(do)充填; D— 肖Ⅲ 段顶部, 团块状结构。所有照片均为单偏光Fig.7 Microscopic features of spotted and lumpy fabrics of the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
4.1.4 团块状结构
由暗色泥粉晶白云石构成, 呈不规则斑块状, 直径介于0.5~1.5 mm之间; 大部分情况下, 组成团块状结构的泥晶重结晶为粉晶, 仅保留黑色泥晶边。团块之间的亮斑由粗晶白云石组成, 面积含量在20%~25%之间(图 7-C, 7-D)。团块在岩石中的面积含量可达70%, 原来可能构成岩石的骨架, 但现今一般呈孤立分散状。见到具棱角状的不规则团块向圆形或椭圆形砂屑颗粒过渡的现象, 推测它们为团块状凝块被水流打碎和磨蚀形成的。该结构在肖尔布拉克组分布局限, 且纵向上不具有连续性, 大部分已完全碎屑化, 主要发现于肖V段中下部, 与砂屑白云岩组合产出, 愈往上团块含量越低, 砂屑含量越高。
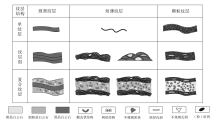
由黑色泥粉晶白云石构成的暗层与中粗晶白云石构成的亮层纵向上交替出现组成。暗色纹层分平直纹层和波状弯曲纹层2大类, 前者平直, 一般是沉积作用造成的, 后者呈不规则弯曲状, 一般是微生物席晶出或捕集碳酸盐泥晶形成。波状纹层是肖尔布拉克组纹层结构的主要类型, 面积含量可达70%, 下文所述纹层皆为微生物成因的纹层结构。
前人曾按成分将微生物成因的纹层进一步划分为致密微晶型、微球粒型和凝聚型(Kennard and James, 1986; Riding, 1991; Dupraz and Strasser, 1999), 其中前两者由生物矿化作用形成, 后者由微生物席捕集碳酸盐颗粒形成。肖尔布拉克组发育多种不同类型的纹层结构(图 8)。在前人分类的基础上, 笔者按成分将纹层分为致密纹层、短薄纹层和颗粒纹层, 同时按纵向组合形式进一步划分为单纹层、纹层组和复合纹层3大类。其中, 复合纹层是该组纹层结构的主要类型, 面积含量大于70%。
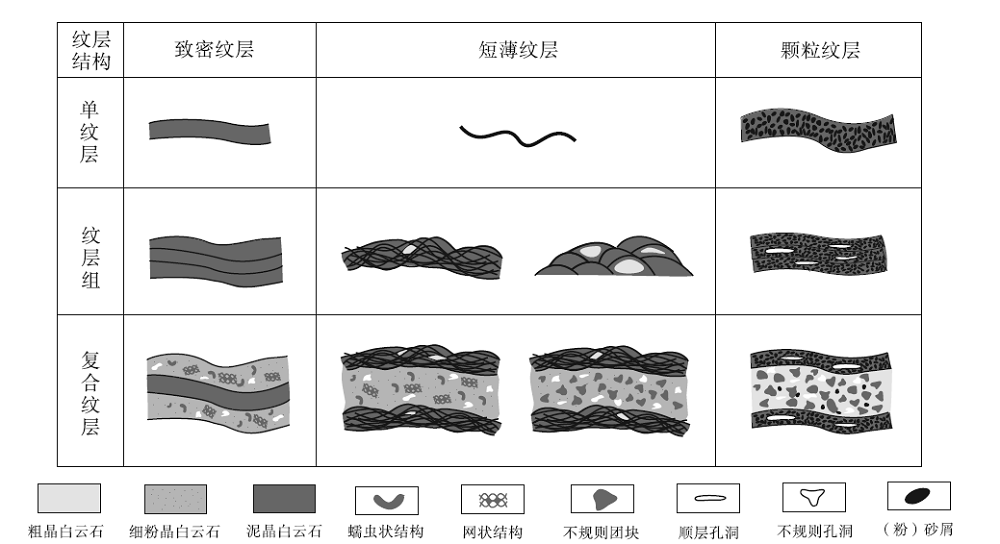 | 图 8 塔里木盆地苏盖特布拉克剖面肖尔布拉克组的纹层结构类型Fig.8 Types of laminated fabrics in the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
致密纹层是由单一泥晶构成深灰色暗层, 上、下为粉晶、细晶或中晶组成的浅灰色亮层(图 9-A, 9-B, 9-C), 横向连续性好, 整体起伏不大, 亮层下表面通常为波状, 一定程度上指示了对微生物席微地形的继承。致密纹层纵向上以单层形式或多层叠置状与亮晶层组成互层, 单层暗层厚度不超过0.5 mm, 亮层厚度变化较大, 介于0.5~2 mm之间。复合纹层的亮层内常有蠕虫状结构、网状结构和点簇状结构伴生, 部分发育由亮晶白云石充填的不规则孔洞。致密泥晶型纹层在该组发育较局限, 主要出现于肖Ⅲ 段, 含量丰富。
 | 图 9 塔里木盆地苏盖特布拉克剖面寒武系第二统第三阶肖尔布拉克组纹层结构显微特征 A— 肖Ⅲ 段底部, 泥粉晶致密纹层(la)与具点簇状结构(clu)的细晶— 中晶亮层组成的复合纹层, 见点簇状结构(clu); B— 肖Ⅲ 段底部, 致密复合型纹层, 细晶亮层内有粉晶白云石组成的网状结构(net), 亮层顶部见压溶缝合线(st); C— 肖Ⅲ 段底部, B图网状结构放大5倍, 不规则网眼(br)被细晶白云石充填, 大小0.05~0.1 mm; D— 肖Ⅴ 段下部, 丝状单纹层(la), 暗色泥晶粉晶白云石组成近同心状纹层, 不连续, 纹层间距约1.5 mm, 粉晶亮层内发育粗晶充填的不规则形状的溶孔; E— 肖V段下部, 叠瓦状纹层组, 泥晶粉晶白云石组成的暗色纹层(la)呈叠瓦状排列, 其间被粗晶白云石充填, 晶间孔(po)发育; F— 肖V段中部, 丝状体(la)相互缠绕构成纹层组(mla), 纹层内存在孔洞(pf), 被粉晶— 细晶白云石充填; G— 肖V段中部, 纹层状细晶— 中晶白云岩, 暗色泥晶白云石组成丝状纹层, 纹层单个(la)或成组(4~5个为1组)(mla), 纹层之间的亮层由细晶中晶白云石组成, 不均匀, 见蠕虫状结构(wo)和粗晶白云石组成的亮斑(br); H— 肖V段中部, 纹层状砂屑粉砂屑粒泥状白云岩, 砂屑(sc)顺层状分布在泥粉晶暗层和亮层内, 顺层的溶洞(sp)被粗晶白云石充填; I— 肖Ⅰ 段上部, 重结晶作用强烈, 泥晶纹层的残余呈短线状, 纹层类型不可辨。所有照片均为单偏光Fig.9 Microscopic features of laminated fabrics in the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
短薄纹层(图 9-D至9-G)不同于致密纹层, 其弯曲程度更高, 单纹层厚度不超过0.1 mm, 起伏幅度为0.1~1 mm, 总长1~2 mm, 呈断续状, 稀疏均匀, 垂向上亮暗叠置韵律较少, 长度变化较大。纹层组有缠绕辫状和叠瓦状2种类型: 前者为由一系列丝状体不规则缠绕而成, 连续性较好, 厚度介于0.8~1.2 mm之间, 亮层通常由中粗晶组成, 厚度一般大于2 mm, 纹层间为泥粉晶白云石, 部分发生溶蚀, 形成粗晶白云石充填的亮斑; 后者由一系列上凸的短纹层垂向上相互叠置而成, 其间常发育晶间孔, 单个短纹层长为2~4 mm。叠瓦状纹层组含量远远少于缠绕辫状纹层组, 因此复合类型纹层主要由缠绕辫状纹层组组成, 亮层与致密纹层类似, 或含不规则团块状凝块。短薄纹层主要见于肖V段中上部, 分布普遍, 纵向上可观察到自下而上由网状结构为主的亮晶层向不规则团块状亮晶层转变。肖Ⅰ 段上部受重结晶作用影响强烈, 仅有断续的、黑色泥晶组成的短线状残余(图9-I)。
由暗色泥粉晶白云石构成的椭圆形、近圆形砂级颗粒组成(图 9-H; 图 10-A, 10-B), 砂屑直径0.1~0.3 mm, 具有明显光滑的颗粒边缘, 呈孤立状或线接触, 发育粒内孔(直径0.1~0.2 mm)和粒间孔(直径0.2~0.3 mm), 颗粒间为中晶白云石。少数砂屑中含有鲕粒, 部分薄片可见砂屑内部经过强烈重结晶作用后已经变成黄褐色细— 中晶白云石, 仅剩黑色泥晶轮廓的残余。镜下可观察到砂屑具有随机散乱分布和顺层排列成颗粒纹层状2种分布方式。砂屑结构主要发育于肖V段上部, 其前身应该是凝块结构, 所以砂屑结构是一种次生结构, 代表了微生物岩遭到非生物作用的改造, 是否属于微生物岩的范畴是一个需要探讨的问题。
 | 图 10 塔里木盆地苏盖特布拉克剖面肖尔布拉克组砂屑结构和骨架结构显微特征 A— 肖Ⅴ 段中部, 白云石化的砂屑粒泥状白云岩— 颗粒状白云岩, 砂屑(sc)近圆形, 直径0.1~0.2 mm, 大多数地方的砂屑之间发生白云石重结晶变成中粗晶(do), 有少数三叶虫碎屑(tr), 少数砂屑中含有鲕粒; B— 肖Ⅴ 段顶部, 中晶白云岩有大量砂屑残余(sc), 推测原岩为砂屑粒泥岩, 砂屑由泥晶白云石组成的边和中晶白云石组成的内核(in)组成; C, D— 肖Ⅴ 段中部, 肾形藻骨架结构(rel), 含盾形生物(du), 发育晶间孔(po)。所有照片均为单偏光Fig.10 Microscopic features of intraclastic fabric and framestone fabric in the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
由相互连接或分离的钙化肾形菌化石形成岩石骨架(图 10-C, 10-D)。肾形菌化石为由一系列泥晶白云石构成的中空球状— 肾状体紧密排列组成的分枝状结构, 大小为(0.5~1) mm× (0.5~2) mm, 内部一般有被粗晶白云石充填的花朵状空腔。关于该结构成因, 目前还没有统一的认识, 部分学者认为肾形菌化石属于蓝细菌门色球菌纲的钙化类型之一(Hofman, 1975; Pratt, 1984), 另有一些学者认为是细菌诱导发生沉淀形成的(Stephens and Sumner, 2002)。肖尔布拉克组肾形菌化石主要分布于肖Ⅴ 段上部, 镜下可见受水流冲刷形成的碎片, 且其上下层位发育有砂屑结构, 故认为它们形成于较高能的浅水潮下带环境。
微生物岩结构受微生物作用和成岩作用双重制约。微生物的类型决定微生物岩的原始结构和原始孔隙特征, 而不同类型的成岩作用对微生物岩的原始结构有不同的改造。根据薄片观察, 肖尔布拉克组微生物岩至少经历了3种主要成岩作用: 准同生白云石化作用、重结晶作用及溶蚀作用。准同生白云石化作用发生的时间最早, 为原始灰质晶体的拟态白云石化, 不破坏微生物岩的原始结构。重结晶作用可以发生在整个成岩期, 使白云石晶体逐步增大, 由泥晶变为粉晶甚至中晶— 粗晶, 凝块的重结晶最终使得其与周围胶结物晶体的大小与颜色趋于一致。薄片观察表明, 肖下段整体重结晶程度高于肖上段, 并且有很强的非均质性, 甚至在同一薄片上可见到重结晶作用程度不同的区域。例如, 肖Ⅳ 段微生物丘的一张薄片上, 可见由重结晶较强的区域向较弱区域过渡的现象(图 11-A)。
 | 图 11 塔里木盆地苏盖特布拉克剖面肖尔布拉克组微生物岩的成岩特征 A— 肖Ⅳ 段, 微生物丘, 重结晶作用使左边的白云石晶体比右边的粗, 颜色较浅, 隐约可见网状结构残余(se); B— 遭受了比照片A更强烈的重结晶作用后的网状结构, 由残余的黑色泥晶白云石(ret)组成; C— 肖Ⅳ 段, 微生物丘, 重结晶作用使蠕虫状结构被破坏而残留斑点状结构(sd); D— 肖Ⅰ 段上部, 重结晶作用使纹层结构仅剩断续状的残余(la); E— 肖Ⅴ 段顶部, 砂屑内部(in)重结晶为细晶白云石, 砂屑的边缘仍然为黑色泥晶白云石; F— 肖Ⅱ 段底部, 溶蚀作用形成的孔洞中充填栉状和半自形方解石。所有照片均为单偏光Fig.11 Diagenetic characteristics of microbialites of the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
蠕虫状结构的重结晶由边缘向中心逐渐进行, 所以凝块中心晶体小, 颜色深, 最终仅残留泥晶白云石组成的斑点(图 11-C)。对网状结构来说, 随着重结晶作用的不断进行, 网线与网眼的白云石晶体大小和颜色趋于一致(图 11-B), 最后只残留交叉点处的斑点状泥晶残余。肖Ⅰ 段的纹层结构重结晶作用较强, 由粉晶构成, 表现为断续、平直— 微波状, 纹层较细且稀疏, 纹层间为细晶白云石, 部分细晶白云石晶间可见黑色泥晶白云石, 原始亚结构难以分辨(图 11-D)。砂屑和团块状结构的重结晶作用一般是从砂屑或团块内部向边缘进行, 重结晶的砂屑和团块内部呈灰白色— 灰黄色粉细晶白云石、边缘由黑色泥晶白云石构成(图 11-E)。砂屑边缘为何会有更强的抵抗重结晶能力, 作者尚无认识。
肖尔布拉克组白云岩主要经历了3种溶蚀作用: 表生期大气水溶蚀作用、埋藏期热液溶蚀作用和有机酸溶蚀作用(沈安江等, 2016; 白莹等, 2018), 可通过准同生期栉状胶结物及埋藏期鞍形晶体进行识别。前人对于以上3种作用对储集层的影响已做过十分详细的研究(沈安江等, 2016), 在此不再细致探讨。总的来说, 溶蚀作用一方面对于微生物岩结构具有破坏作用(图 11-F), 另一方面具有改善储集层的作用。图 12粗略概括了不同微生物岩结构类型受成岩改造的演化特征。
 | 图 12 塔里木盆地苏盖特布拉克剖面不同微生物岩结构演化模式简图Fig.12 Evolution models of microbialites fabrics in the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
总体而言, 肖下段微生物岩结构受成岩作用破坏程度更高的特征。肖Ⅰ — Ⅱ 段为黑色致密白云岩, 主要见断续状稀疏纹层结构及粉晶或细晶化的凝块结构, 证明重结晶作用强烈, 而窗格构造的发育说明表生期经历了较强的溶蚀作用。肖Ⅲ 段含有灰质, 推测经历了与肖Ⅰ 段、肖Ⅱ 段不一样的成岩历史。该段凝块结构含量减少, 纹层结构较为发育, 且主要为致密纹层, 连续性较好, 保存完整, 但纹层间的网状结构和蠕虫状结构破坏较严重, 多残留点簇状结构(图 7-B; 图 9-A)。肖Ⅳ 段及肖V段下部主要为蠕虫状结构和网状结构, 仅部分发生不均匀重结晶化, 总体结构轮廓易于辨别。肖V段上部开始出现砂屑结构及纹层结构, 纹层间虽可见部分溶蚀孔, 但纹层重结晶作用弱, 亮暗层交替的特征保存较好, 纹层种类最为丰富, 部分纹层间夹杂的网状结构或蠕虫状结构保存较好(图 9), 而砂屑颗粒除部分内部重结晶为细晶外, 亦保存较完整。
微生物岩原始结构可一定程度反映其形成环境的特征, 如水体深度、水动力强度等。一般来说, 叠层石形成环境的水深要浅于凝块石, 主要形成于潮间及潮下带; 凝块石则主要形成于低能、低沉积速率的浅潮下带(Aitken, 1967; Kennard and James, 1986; Ezaki et al., 2003); 层状叠层石一般形成于水动力能量较弱的平均高潮面附近或潮下带, 丘状、柱状叠层石形成于较高能的潮间带; 树枝状凝块石可形成于蒸发台地潟湖相中(钱迈平, 1991; Jahnert and Collins, 2012; Mettraux et al., 2015)。目前, 大部分学者的研究是通过微生物岩的中型结构判断形成环境, 对显微结构与环境的关系尚缺乏研究。
前人研究认为, 肖尔布拉克组整体为台地沉积(冯增昭等, 2002, 2006; 胡明毅等, 2019)。对于沉积相的解释不同学者持不同观点: 白莹等(2018)认为, 阿克苏地区肖尔布拉克组沉积时期整体处于碳酸盐缓坡向镶边碳酸盐岩台地过渡地区, 肖下段以微生物层状建造为主, 位于台缘带和台内带, 肖上段发育凝块石礁和表附菌礁2种微生物礁建造, 主要位于台缘礁带。郑剑锋等(2019)、乔占峰等(2019)则认为阿克苏地区肖尔布拉克组整体为缓坡背景下的丘滩— 滩坪沉积。笔者研究发现, 肖下段岩石主要由泥粉晶白云石组成, 几乎不含碎屑和颗粒, 代表了一种水动力能量较弱的局限水体环境; 从肖V段中部开始, 出现砂屑结构和肾形菌骨架结构, 两者皆是水动力变强的标志。因此, 苏盖特布拉克一带肖尔布拉克组沉积期间为局限台地向开阔台地的过渡环境。
肖Ⅰ 段主要发育泥粉晶薄层状层纹石, 下部常见鸟眼构造, 指示潮间— 潮上带环境; 向上鸟眼构造逐渐减少, 且缺乏潮上带特征, 指示水体逐渐加深, 变为潮下带。肖Ⅱ 段主要发育似薄层状凝块石白云岩, 可能是薄层构造被成岩作用改造所形成的。肖Ⅱ 段中下段普遍有扁平的、顺层分布的厘米级的溶孔, 因为溶孔顺层分布, 可能是由原来的鸟眼孔溶蚀扩大形成的, 所以此段可能原来也普遍具有鸟眼构造。因此, 推测其形成环境可能也是潮间带— 潮上带。肖Ⅱ 段上部发育泥粉晶网状结构凝块石白云岩及少量层纹石白云岩, 不发育鸟眼构造, 可能为潮下带环境。肖Ⅲ 段为泥质含量高、层理极其平直、发育毫米级微生物纹层的极薄层白云岩, 推测为水动力能量较弱的潮间— 潮上带环境。
肖Ⅳ 段发育散布的凝块石微生物丘, 单个丘体的高度至少6 m, 且这段地层的厚度在20 m以上, 代表了较深水的局限潮下带环境。 肖Ⅴ 段下部以致密的网状凝块岩为主, 代表潮下带环境。 肖Ⅴ 段上部以纹层结构的微生物岩为主, 再向上出现砂屑结构、 砂屑— 纹层复合结构及骨架结构, 部分岩石内发现三叶虫碎片, 指示了一种开阔的浅水较高能环境。
因此, 肖尔布拉克组的沉积环境可以分为明显不同的3个阶段: 肖Ⅰ ~Ⅲ 段以潮坪环境为主; 肖Ⅳ 段及肖V段下部代表较深水的环境; 肖V段上部代表浅水的潮下环境(图 13)。
 | 图 13 塔里木盆地苏盖特布拉克剖面肖尔布拉克组微生物岩形成环境及其演化模式Fig.13 Formation environmental model of microbialites of the Xiaoerbulake Formation of Sugaitebulake section, Tarim Basin |
1)塔里木盆地苏盖特布拉克剖面寒武系第二统第三阶肖尔布拉克组白云岩全部为微生物成因的碳酸盐岩, 具有层状和丘状2种宏观产状。微生物岩结构类型主要包括凝块结构和纹层结构, 其次还有骨架结构和砂屑结构。
2)根据显微特征, 将凝块结构划分为4种亚类: 蠕虫状结构、网状结构、斑点状结构和团块状结构。网状结构是主要类型, 主要见于肖Ⅱ 段、肖Ⅳ 段及肖V段下部。按成分将纹层结构划分为致密纹层、短薄纹层、颗粒纹层, 按组合形式将纹层结构划分为单纹层、纹层组、复合纹层。
3)该组微生物岩原始结构主要受白云石化作用、重结晶作用及溶蚀作用影响。白云石化作用主要为准同生期的拟态白云石化, 对原始结构破坏较小; 而重结晶作用和溶蚀作用几乎贯穿整个成岩期, 对微生物岩结构破坏较大。微生物岩结构对成岩作用改造的抵抗力为: 砂屑结构> 纹层结构> 凝块结构。
4)肖Ⅰ 段至肖Ⅲ 段沉积环境主要为潮坪, 肖Ⅳ 段和肖V段下部形成于较深水环境, 肖V段上部形成于浅水潮下环境。纵向上, 肖尔布拉克组沉积环境依次经历了由局限浅水潮坪— 局限深水潮下带— 开阔浅水潮下带的演变。
文章贡献说明 吴亚生和潘文庆负责项目设计和研究指导; 李莹、吴亚生负责显微结构研究; 杨果、孙崇浩、姜红霞负责中观和宏观结构研究; 吴亚生、郑丽婧、张月阳、蒋子文负责野外剖面测量和采样。文章撰写由李莹完成。吴亚生和姜红霞负责文章审核、修改、定稿。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|