



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
化石及现生真蕨类植物的根系:形态演化及古环境意义*
[刘璐1  , 刘乐
, 刘乐2 , 王德明1 , 薛进庄1 ]
, 刘乐]
|
|

第一作者简介 刘璐,女,1994年生,北京大学地球与空间科学学院古生物学与地层学专业博士生,主要研究侏罗纪古土壤与植物根系。E-mail: liulu0406@pku.edu.cn。
根是植物的重要营养器官,对维持植物生理活动及生态系统物质循环和能量流动极为重要。真蕨类植物的根系以可预测的方式起源于枝,被称为枝生根,通常也被称为不定根。相比于对种子植物根系的广泛研究,对真蕨类植物根系的认识较为薄弱。通过广泛收集各类文献中有关真蕨类根系的信息,以及观察大量化石、现生植物标本,对若干化石及现生真蕨类植物根系的形态学特征进行了综述。在繁盛于中、晚泥盆世的早期真蕨类植物中,密集的枝生根着生于茎轴、或推测的根状茎、或主干膨大的基部上;根系的分枝罕见,若存在,则仅表现为等二歧式分枝;而在真蕨纲冠群植物中,枝生根密集或稀疏,根系呈单轴式分枝,侧根发育,可达 2级甚至更多。真蕨类根系统的形态演化可采用“顶枝学说”进行解释: 真蕨类植物的根系具有一定的演化保守性,均为枝生根;早期真蕨类的原始根系可视为不分枝的或偶见二歧式分枝的顶枝束;经历顶枝束的越顶生长,侧根系统在真蕨纲冠群中演化出现。真蕨类根系的构型、形态、解剖结构、生物量分配等信息可在化石记录中得以揭示,这些信息可为探讨植物根系的演化及研究地史时期的植物—土壤系统提供重要依据。最后,简述了这方面研究的 1个案例,即北京西山侏罗纪地层中的真蕨类根系化石及根系—土壤系统。
About the first author Liu Lu,born in 1994,is a doctoral candidate at Peking University. Her current research focuses on the Jurassic paleosols and plant roots. E-mail: liulu0406@pku.edu.cn.
Roots are important vegetative organs of plants,and play essential roles in plant physiology as well as in the nutrient cycle and energy flow of an ecosystem. Fern roots are known to arise from the shoots in a predictable pattern,and thus are called shoot-borne roots,or more commenly adventitious roots. In contrast to the extensive studies of seed plant roots,relatively little information is available on fern roots. Based on a compilation of the available data and our own observations on both fossil and extant fern specimens,this study presents a review of root morphology of several fossil and extant ferns. In the early ferns that flourished during the Middle-Late Devonian,dense shoot-borne roots are attached on stems,or inferred rhizomes,or on expanded bases of trunks;such roots seldom branch,but when they do they display an isotomous branching pattern. In the crown groups of ferns,however,shoot-borne roots may be either densely or sparsely arranged,and show a monopodial branching pattern with two or more orders of lateral roots. The morphological evolution of fern roots may be interpreted by“the telome theory”: shoot-borne roots seem to be evolutionary conservative;primitive roots in early ferns can be considered as telome trusses that occasionally branch isotomously;and lateral rooting systems in crown groups of ferns evolved through overtopping of telome trusses. Information on fern root architecture,morphology,anatomical structure,and allocation ratios is documented in the fossil record,and is of significance in understanding the evolution of plant rooting systems and plant-soil interactions over geological time. As a case study,fossils of fern roots and root-soil systems from the Jurassic of Western Hills,Beijing,are briefly introduced.
根是植物地下部分(除少数气生根外)的营养器官, 对维持植物生理活动及生态系统物质循环和能量流动极为重要(Osmont et al., 2007)。除了众所周知的支撑、吸收和传输功能之外, 根还具有合成、贮藏、分泌等作用。根合成的物质包括多种氨基酸、生长激素和植物碱, 这些物质对植物地上部分的生长、发育有着较大的影响(马炜梁等, 2009)。一些植物根内的薄壁组织较发达, 能贮藏有机物质。根的分泌物如糖类、氨基酸、有机酸、维生素、核苷酸和酶等, 可以促进土壤中的微生物生长, 进而在根表面及其周围形成一个特殊的微生物区域(根围, rhizosphere), 这些微生物对植物体的代谢活动、吸收作用、抗病性等有促进作用, 并能减少根在生长过程中与土壤的摩擦力(马炜梁等, 2009; Bellini et al., 2014)。细根(通常指直径小于2 mm的根)是根系中高度动态的组成部分, 不断生长、死亡和分解, 是土壤有机质及养分的重要来源(Bellini et al., 2014)。植物根系活动直接影响外界环境, 如植物可以通过纵横交错的网状结构形成坚固的根土复合体(Xue et al., 2016)。良好的根系构型不仅能提高根系对土壤养分和水分利用的效率, 也是构建稳定生态群落的基础(毛齐正等, 2008)。目前在植物茎、叶系统的性状演化方面的研究取得了重大进展, 但对根系统以及根与其他器官之间关系方面的认识仍然有限(McCormack et al., 2017)。
近年来, 植物根系的起源、演化及对地球系统的影响, 引起了学术界的广泛关注。现有化石证据显示, 最古老的根系为早泥盆世的工蕨类植物Zosterophyllum shengfengense(胜峰工蕨)的簇生根(Hao et al., 2010), 这种纤细的根提供了植物向地性生长的最早证据。下泥盆统的莱尼燧石层中保存了早期维管植物精美的内部结构, 如Aglaophyton带有假根的根状茎、Asteroxylon的根系中已知最早的根尖分生组织等(Kenrick and Crane, 1997; Hetherington and Dolan, 2018)。研究者认为, 自泥盆纪以来, 特别是自中泥盆世高大的树木产生以来, 植物根系的生长和活动直接或间接地影响着大陆风化, 在土壤形成和地表风化中发挥着重要作用(Morris et al., 2015; Pawlik et al., 2016)。
真蕨类植物在陆地生态系统中具有特殊的生态位, 其根系结构与种子植物不同(Hou and Blancaflor, 2009)。相比较于对种子植物根系的遗传、演化及分子生物学等多个方面的广泛研究, 对真蕨类植物根系的相关研究一直较为滞后, 文献也较为分散, 因此有必要进行综合梳理与讨论。通过广泛收集各类资料, 综合分析各类文献中有关真蕨类根系的数据, 以及观察大量化石、现生植物标本, 总结了真蕨类中若干类群的根系的形态学特征, 分析、对比其异同, 并对真蕨类植物根系的形态演化提出初步解释。最后, 结合笔者等在北京西山侏罗纪地层中获取的真蕨类根系的新材料, 讨论如何在化石记录中识别真蕨类根系并分析其古环境意义。
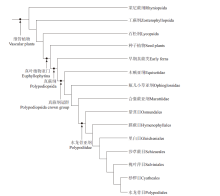
在传统的分类体系中, 蕨类植物门被分为5个亚门: 松叶蕨亚门、石松亚门、水韭亚门、楔叶蕨亚门和真蕨亚门; 通常, 前4个亚门被称为拟蕨类, 而真蕨亚门被称为真蕨类(秦仁昌, 1978a, 1978b; 陆树刚, 2007)。然而, 分子系统学的兴起对传统的蕨类分类系统提出了挑战(Pryer et al., 2001; 李春香等, 2004, 2007)。蕨类植物系统发育工作组(The Pteridophyte Phylogeny Group)整合大量研究成果, 提出了一个新的有关现生蕨类植物的分类方案(PPG I, 2016), 其影响力日益扩大, 逐渐成为蕨类植物系统学研究的重要参考。PPG I系统中, 现生蕨类植物被划分为石松纲(Lycopodiopsida/lycophytes)与真蕨纲(Polypodiopsida/ferns), 前者包括传统分类中的石松亚门和水韭亚门, 而后者包括松叶蕨亚门、楔叶蕨亚门及真蕨亚门。真蕨纲与种子植物互为姐妹群, 共同构成真叶植物支系(euphyllophytes)。真蕨纲分木贼亚纲、瓶儿小草亚纲、观音座莲亚纲及水龙骨亚纲, 其中水龙骨亚纲包括7个目。
值得注意的是, PPG I系统针对的是仅现生植物, 该系统定义的真蕨纲实际上仅为真蕨纲冠群, 即现生所有真蕨类植物的共同祖先及其所有后裔(有关冠群的释义, 请参见薛进庄和张华侨, 2010)。因此, 如何将化石类群融入到PPG I系统中, 形成更为完整的有关蕨类植物的分类体系, 是未来工作的一个重要挑战。目前, 很多化石类群的系统位置并未取得共识(薛进庄和郝守刚, 2014; 薛进庄等, 2014; Li et al., 2020)。化石记录显示, 古生代期间繁盛着一些现生真蕨类的基干类群, 它们被归入枝蕨纲(Cladoxylopsida)或早期似真蕨类植物(Early fernlike plants), 包括瘤指蕨目、伊瑞蕨目、羽裂蕨目等多个类群(Taylor et al., 2009; Xue et al., 2010; Wang et al., 2015)。这些类群之间、以及它们与真蕨纲冠群之间的系统关系仍有待确定。为了论述方便, 笔者将这些基干类群统称为早期真蕨类(early ferns), 暂时作为真蕨纲冠群的姐妹群。国内一些研究现生蕨类的学者, 将lycophytes和ferns分别翻译为石松类和蕨类(刘红梅等, 2008; 张宪春等, 2013), 但若兼顾化石类群, ferns译为真蕨类或广义真蕨类, 可能更为妥当(薛进庄等, 2014)。笔者认为, PPG I系统中的ferns实为真蕨纲冠群, 而早期真蕨类+真蕨纲冠群, 构成真正完整的真蕨纲(图 1)。文中涉及的类群包括早期真蕨类、合囊蕨目、紫萁目、膜蕨目、里白目、槐叶萍目、桫椤目、水龙骨目中的若干代表性植物。
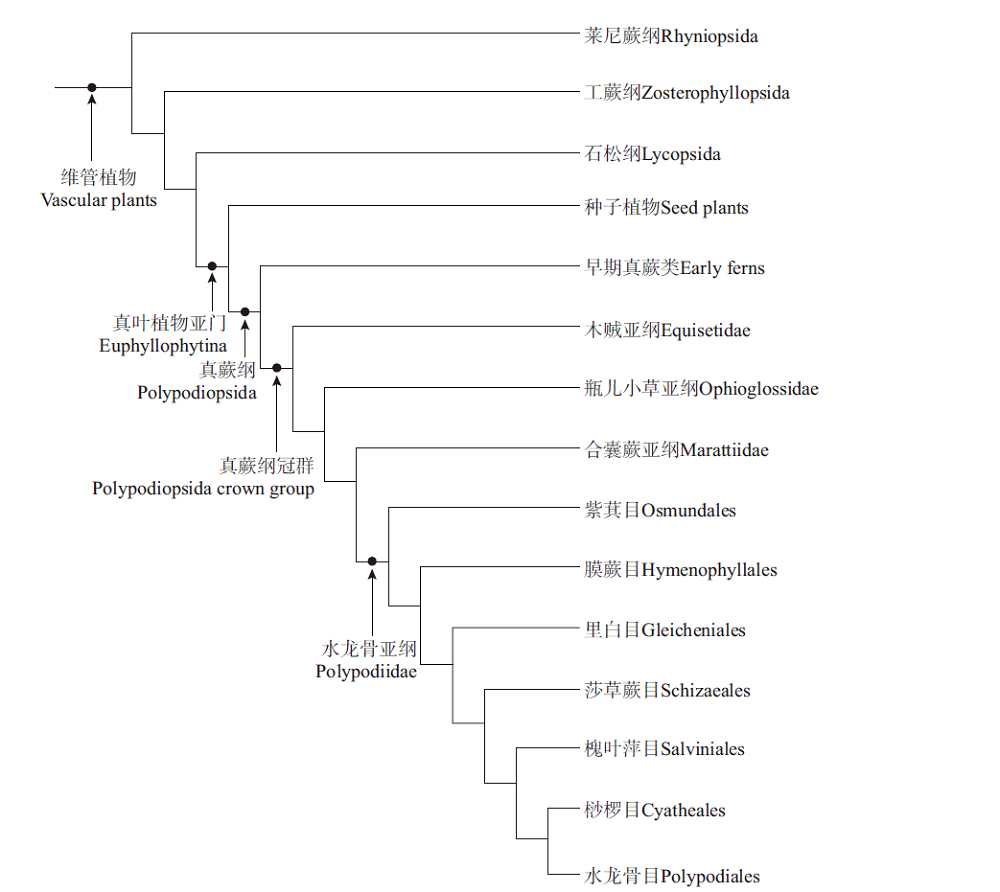 | 图 1 维管植物系统发育树, 示真蕨类及其内部类群的系统位置(依据Hao and Xue, 2013; PPG I, 2016)Fig.1 Phylogenetic tree of vascular plants, showing the phylogenetic positions of ferns and their sub-groups(based on Hao and Xue, 2013; PPG I, 2016) |
1.2.1 根、枝、根状茎
Raven和Edwards(2001)对根的定义进行了详细阐述, 文中主要依据他们的论述, 介绍其中的一些要点。根是维管植物孢子体的轴性多细胞结构, 从胚轴、其他的根或者茎上发生; 通常在地下生长, 具有严格的顶端伸长生长模式。根具有典型的向地性, 可分枝形成侧根。在发育过程中, 由1个或多个顶端分生组织细胞分裂产生根冠、中柱原始细胞、皮层和表皮。根尖顶端分生组织通常由根冠覆盖。根冠可以通过分泌黏液使成熟的根粘附到土壤中, 根冠脱离细胞(根部边缘细胞)可以作用于根围的微生物。侧根从中柱鞘中形成, 然后穿过皮层和表皮。
枝是维管植物的另一套器官系统, 通常包括茎轴、叶、孢子囊和它们的变体(如球果、花以及它们的衍生物如种子和果实)。它们在地上生长, 与根有很多不同的习性, 包括反向重力性及茎端分生组织不产生类似根冠的组织(Raven and Edwards, 2001)。枝的附属物如分枝和叶都产生自表面, 或称外起源(Groff and Kaplan, 1988)。
根状茎(rhizome)在真蕨类植物及种子植物中广泛存在, 通常为根的着生器官, 但经常被混淆为根系的一部分(Shekhar et al., 2019)。很多学者曾经尝试给根状茎做出精确的定义, 但往往不能全面地概括其丰富的变异。笔者采用Bell和Tomlinson(1980)较为宽泛的定义, 他们认为根状茎指的是植物在基质之上或基质内部由轴的延伸或分枝而产生的变态茎, 因此根状茎具有茎的属性, 可以是肉质的、匍匐的、气生的、具鳞片的或具叶的(Bell and Tomlinson, 1980)。
1.2.2 不定根、枝生根
长期以来, 有关植物根系的概念主要源自对种子植物的研究。裸子植物和双子叶植物通常为直根系, 包括主根和侧根, 主根通常较粗, 可能形成被称为侧根的次级根系(Cannon, 1949)。侧根进一步分枝, 形成更高级别的侧根。虽然不同种类的植物具有不同样式的根系统, 但是直根系植物都有一个起源于胚根的主根。单子叶植物的主根较小, 生长时间很短, 形成不久后就死亡或者停止生长, 在胚后期从枝、叶或者茎基部的节上生出许多不定根, 被称为须根系(adventitious root system)(Cannon, 1949; Osmont et al., 2007; Bellini et al., 2014)。
不定根在种子植物及真蕨类植物中均有发育, 起源于气生组织的不定根, 通常源自茎, 有时源自叶(Bellini et al., 2014)。大多数情况下, 不定根的发生是由于植物器官受伤或激素、病原微生物等外界因素的刺激, 因此表现为植物的再生反应(Bellini et al., 2014)。不定根会被一些自然现象或人为因素(如切割伤害)以及激素诱导产生。此外, 不定根是植物无性繁殖的关键组分。目前, 不定根的来源及发育机制仍不清楚, 它们可能来源于中柱鞘、韧皮部或木质部薄壁细胞等(Bellini et al., 2014)。不定根也可分枝形成侧根。
真蕨类植物的根具有特殊的发育模式。大多数真蕨类植物的根来源于一个根尖细胞(Hou and Blancaflor, 2009)。对于真蕨类植物, Groff和 Kaplan(1988)建议使用枝生根这个术语来取代不定根, 因为发育生物学的研究表明, 真蕨类植物的根是以可预测的方式起源于枝的, 包括茎生根和叶生根2类(Hou and Blancaflor, 2009)。茎生根起源于正在发育的茎, 而叶生根起源于叶的基部; 发育过程中, 2种类型的枝生根均可产生侧根。在现生真蕨纲植物中, 少数种类如Salvinia和一些Trichomanes的种不具根(Bierhorst, 1971), 其他大多均可以形成枝生根(Groff and Kaplan, 1988)。但是值得注意的是, 不定根这一术语广泛使用于以往有关真蕨类植物的绝大多数文献中。因此, 笔者对真蕨类植物的叙述中, 根系、根、不定根及枝生根(具或不具侧根)这几个术语的内涵是一致的。
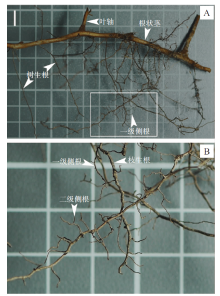
图 2以Dicranopteris dichotoma(Thunb.) Bernh.(芒萁)为例, 显示了典型的现生真蕨类植物的根状茎、叶轴、枝生根及侧根的着生关系。
 | 图 2 真蕨类植物根系的术语示意图 A— 以Dicranopteris dichotoma(Thunb.)Bernh.(芒萁)为例, 示横生的根状茎及着生于根状茎的枝生根; 比例尺: 1 cm。B— 图A的局部放大(白色方框区域), 示枝生根、一级侧根和二级侧根Fig.2 Diagrams showing the terminology for roots of ferns |
早期真蕨类始现于中泥盆世, 包括的属种众多、类群庞杂, 与现生真蕨类一些类群的起源有密切的关系(Taylor et al., 2009)。中、晚泥盆世的早期真蕨类, 对于探讨真蕨类根系的演化具有尤为重要的意义。下文介绍这一时段的6个代表属、种的根系特征。
Melvillipteris quadriseriata发现于加拿大极区Melville岛晚泥盆世地层(Xue and Basinger, 2016)。该植物的主轴直径3~5 mm。一些主轴呈Z字型, 其节间着生密集的枝生根, 超过15个根形成簇状(图 3-A), 表明这些主轴可能与潮湿的基质直接接触。根在主轴相邻的2个节间上交错生长, 在每个节间, 根只着生于主轴的一侧; 即在第1个节间, 根向一侧展开, 而在与之相邻的第2个节间, 根向另一侧展开(图 3-A)。主轴上的枝生根宽为0.5~0.9 mm, 保存的最大长度27 mm。
 | 图 3 早期真蕨类植物Melvillipteris和Denglongia的根系 A— Melvillipteris quadriseriata Xue et Basinger, Z字型主轴以及着生在节间的密集枝生根(箭头), 标本号US734-9144, 收藏于加拿大萨斯喀彻温大学地质学系(首次发表于Xue and Basinger, 2016, Fig.3h), 比例尺: 10 mm; B— Denglongia hubeiensis Xue et Hao, 主轴的节部之下着生密集的根, 未正式编号标本, 比例尺: 5 mm; C— Denglongia hubeiensis Xue et Hao, 主轴的枝生根, 具至少1次等二歧式分枝(箭头), 标本号PKU-XH119a(首次发表于Xue and Hao, 2008, Fig.2d), 比例尺: 2 mm; B、C标本收藏于北京大学地质学系Fig.3 Roots of the early ferns Melvillipteris and Denglongia |
Rhacophyton(羽裂蕨)常见于欧美地区晚泥盆世晚期的地层中(Taylor et al., 2009)。在其中的1个种Rhacophyton ceratangium中, 主轴节间的一侧具有枝生根, 且枝生根也成簇分布(Cornet et al., 1976; Scheckler, 1986), 反映了与Melvillipteris相似的生长习性。
Denglongia hubeiensis(湖北灯笼蕨)发现于湖北长阳上泥盆统黄家磴组(Xue and Hao, 2008)。该植物具主轴和至少2级侧枝。一级枝以轮生方式着生于主轴上, 主轴和一级枝均具有枝生根(图 3-B, 3-C)。灯笼蕨为一类草本植物, 着生在主轴基部的节下方的枝生根较为密集, 长可达10 cm, 宽度平均为0.8 mm, 可能生长在潮湿地面。有些枝生根具等二歧式分枝, 但仅可见分枝1次(图 3-B)。相比较而言, 着生在一级侧枝上的枝生根更短(长0.4~2 cm)且更稀疏, 这些枝生根可能类似于一种固着器官, 使得侧枝从母体植物中分离后可以找到新的支撑, 灯笼蕨可能利用这种方式去扩展更多的生态空间(Xue and Hao, 2008)。
Shougangia bella(优雅守刚蕨)是产自中国安徽、江苏等地上泥盆统五通组中的一类早期真蕨类植物(Wang et al., 2015)。该植物具有部分匍匐的茎轴和单侧着生的枝生根, 根只着生在根状茎的一侧, 形成密集的簇。每个压型保存的根中具1个维管束痕迹。根的长度可达60 mm, 宽度为0.4~1.0 mm, 根中的维管束痕宽约0.1 mm。
Matacladophyton tetrapetalum产自中国湖北中泥盆世晚期(吉维特期)地层(Wang and Geng, 1997), 其气生部分包括3级分枝。一个标本显示根状茎由水平延伸至直立向上弯曲, 直径约9 mm, 在节间底部可以看到2簇根, 根的长度达8 cm, 但未保存末端部分。每条根的直径约2.5 mm, 未观察到分枝。
在最近报道的美国纽约Cario附近的中泥盆世化石森林中, 识别出早期真蕨类Eospermatopteris的根系特征(Stein et al., 2020)。Eospermatopteris是一类树蕨状植物, 高度可达8 m, 构成地球上最早森林的一部分。Eospermatopteris的根着生在茎干基部的底面和侧面, 呈近水平状辐射, 密集排列, 延伸至(古)土壤表层以下1~2 m。根的直径为7~10 mm, 表面平滑或具纵向皱褶, 极少分枝。根向下延伸到古土壤中, 表明Eospermatopteris的植株在洪水期仍可以挺立, 而且可能在洪水过后的一段时间内仍然可以存活。这也表明, 早期的根系已经对真蕨类植物的生存和群落复苏发挥着重要的作用。
现生合囊蕨亚纲仅包括1目(合囊蕨目)、1科(合囊蕨科)、6属(PPG I, 2016)。合囊蕨目的化石记录可以追溯到石炭纪, 数量多、分布广, 其中最为常见且了解最为详细的化石代表为Psaronius(辉木)(Taylor et al., 2009)。Psaronius是一种约10 m高的树蕨, 繁盛于石炭纪和二叠纪, 化石材料以矿化的茎为主, 如Psaronius sinensis(中国辉木)等, 其解剖信息详细, 但根系的形态学信息较少(中国科学院南京地质古生物研究所和中国科学院植物研究所, 1974)。少量的化石证据以及茎轴切片的证据表明, 辉木的枝生根在茎下部的一段距离形成, 并向下延伸, 形成一个密集的根套(Taylor et al., 2009)。
现生紫萁目仅包括1科即紫萁科, 为多年生草本或小型树形蕨类植物(陆树刚, 2007)。现生紫萁科包括Osmunda、Todea和Leptopteris等6个属(PPG I, 2016)。文中依据中国科学院植物研究所植物标本馆的收藏标本, 选取了紫萁科的若干代表属种进行观察和测量(图 4)。紫萁科植物具斜生至直立的根状茎或地上直立树状茎, 叶柄长而坚实, 基部膨大。不同属种的根状茎差别较大, 有的短且粗壮, 有的则长而匍匐生长。粗壮的根状茎直径2~6 cm(图 4-A, 4-E至4-I), 其上着生枝生根。枝生根或长或短, 或密或疏(个别属种可见枝生根十分密集, 生物量较大; 图 4-B, 4-C), 至少具一级侧根, 或进一步分枝形成更高级别的侧根。匍匐生长的根状茎长度可达17 cm(图 4-D), 其上的枝生根较为稀疏, 侧根也少有发育。
 | 图 4 紫萁科代表属种的根系 A— Osmunda cinnamomea var. asiatica Fernald, 标本号PE00289592; B— Osmundastrum cinnamomeum(L.)C. Presl., 标本号PE01623267; C— Osmunda japonica Thunb.; D— 属种未定; E— Osmunda japonica Thunb., 标本号PE01396600; F— Osmunda angustifolia Ching, 标本号PE00289559; G, H, I— 属种未定。比例尺: 20 mm(A~F); 10 mm(G~I)。标本收藏于中国科学院植物研究所植物标本馆Fig.4 Roots of representative plants of the Osmundaceae |
紫萁科的化石记录较为丰富。迄今为止, 世界范围内已经发现了紫萁科根状茎化石约14属80余种(Tian et al., 2008)。Tian等(2013)、田宁等(2014)报道了产自辽西侏罗系的2种紫萁科的矿化化石, 其中Ashicaulis beipiaoensis(北票阿氏茎)的根状茎直径为4.0~4.5 cm(含叶柄基和根套), 被叶柄基部和不定根包裹; Ashicaulis wangii(王氏阿氏茎)的根状茎直径为3.0~5.0 cm(含叶柄基)。发现于瑞典侏罗纪地层的Osmunda pulchella, 根状茎直径可达4 cm, 被紧密的、螺旋状排列的叶柄基和散布的枝生根包围(Bomfleur et al., 2014, 2015)。解剖学证据显示紫萁科的根多数发育自根茎中柱的木质部圆筒, 个别可见发育自叶柄基的木质部。紫萁科矿化根状茎材料的内部结构信息丰富, 其枝生根的基部直径0.4~1.0 mm, 维管束为二极型。但外部形态学特征有限, 缺乏枝生根的度量、密度以及是否具侧根等信息。
膜蕨目包括膜蕨科1科9属, 约434种(PPG I, 2016)。膜蕨类植物非常小, 通常长度只有几个厘米。匍匐的根状茎更细长, 其上着生一些非常细的根。Hymenophyllum barbatum(华东膜蕨)可见有细长的根状茎, 枝生根着生在根状茎上, 短小且几乎没有侧根发育(陆树刚, 2017, p.44, 图 212)。Schneider(2000)描述了若干膜蕨科植物的根以及它们的特征, 这些植物的根状茎直立、半直立或匍匐, 一些属种的根状茎具直径超过0.5 mm的枝生根, 另一些属种则不具或很少具根。
邓胜徽(1995)、Deng(1997)报道了中国内蒙古早白垩世地层中的2种已灭绝的膜蕨科植物(Eogonocormus cretaceum和Eogonocormus linearifolium), 认为它们具有较小的匍匐根状茎, 但其枝生根的情况不明。
里白目包括3个科, 即里白科、双扇蕨科和马通蕨科。笔者依据中国科学院植物研究所植物标本馆的收藏标本, 观察并测量了里白科植物的若干标本(图 5)。里白科植物的根状茎较长且横向匍匐生长, 向上发育叶轴。Dicranopteris dichotoma的根状茎长度可达22 cm, 宽约2.7 mm。根状茎上着生枝生根, 长短不一, 枝生根具1至2级的侧根(图 5-B)。据测量, Dicranopteris dichotoma的枝生根长度为9.9~53 mm, 宽为0.4~1.5 mm, 分枝密度为5次/10 mm(N=20); 一级侧根长0.9~4.3 mm, 宽为0.1~0.3 mm, 分枝密度为8次/10 mm(N=12)(图 5-B)。Calymella squamosissima的根状茎长43~93 mm(但不完整), 宽为2.2~2.5 mm(N=2); 枝生根长13~50 mm, 宽0.4~0.6 mm, 分枝密度为6次/10 mm(N =12), 一级侧根长0.9~3.3 mm, 宽0.2~0.3 mm, 分枝密度为5次/10 mm(N=8)(图 5-D)。
 | 图 5 里白科代表属种的根系 A— Dicranopteris ampla Ching et Chiu, 标本号PE00233994, 比例尺: 20 mm; B— Dicranopteris dichotoma (Thunb.)Bernh., 标本号PE00234158, 比例尺: 20 mm; C— Dicranopteris linearis(Burm. f.)Underw., 标本号PE234223, 比例尺: 10 mm; D— Calymella squamosissima Nakai, 标本号PE01623521, 比例尺: 10 mm。标本收藏于中国科学院植物研究所植物标本馆Fig.5 Roots of representative plants of the Gleicheniaceae |
在化石记录中, 发现于美国新泽西州晚白垩世地层的Boodlepteris turoniana被归入里白科, 其根从根状茎的腹面发出, 叶轴从根状茎的背面发出。根状茎直径1.1 mm, 横截面呈圆形, 枝生根直径0.1~0.2 mm(Gandolfo et al., 1997)。发现自加拿大不列颠哥伦比亚地区始新世地层的Gleichenia appianensis, 也被认为属于里白科, 该植物的根状茎长7.5 mm, 直径3.0 mm, 根从根状茎的各个方向伸出(Mindell et al., 2006)。印度Rajmahal地区发现的保存解剖构造的根化石Gleichenioamyelon diarcha, 推断应归入里白科, 其根状茎圆柱形, 直径3.8~5.5 mm, 单侧分枝, 外表面光滑或具小的多细胞鳞片(Bohra and Sharma, 1979)。
槐叶萍目为浮水生, 仅包括1科即槐叶萍科。槐叶萍(Salvinia natans)植物体漂浮水面, 茎横卧, 有毛, 无根。满江红(Azolla pinnata subsp. asiatica)植物体小, 茎横卧, 羽状分枝, 根垂于水中。这2种植物在中国广泛分布(马炜梁等, 2009)。发现于日本西部晚侏罗世地层的Regnellites nagashimae被认为属于槐叶萍目, 具有长、匍匐状的根状茎, 直径2.0~3.0 mm, 发育大量的直径约0.2 mm的枝生根(Yamada and Kato, 2002)。此外, 发现于阿根廷晚白垩世地层的Azolla coloniensis, 其根直接着生在茎上, 根长可达2.2 mm, 具有维管束, 不分枝; 发现于阿根廷古新世地层的Azolla keuja具有簇生根, 每一簇有9~23个根, 根不分枝, 根簇长5.9~10.3 mm, 根宽0.2~0.3 mm(Hermsen et al., 2019)。
现生桫椤目植物包括8科13属, 其中桫椤科的属种最为丰富, 共3属约643种(PPG I, 2016), 大多数植物为树蕨, 有高达20 m的树干状茎, 但也有横走茎的小型植物。桫椤科的化石记录可追溯至侏罗纪(邓胜徽, 2007)。朱家柟(1963)在内蒙古鄂尔多斯罕台川中侏罗统报道了Cyathea(桫椤属)的1个种, 但标本仅保存有羽片及孢子囊, 无解剖和根茎结构。保存有解剖结构的树蕨类化石材料十分丰富, 如Vera(2015)报道的采自南极洲早白垩世地层的树蕨类矿化茎干(属种未定), 其周围保存有不定根套, 不定根具原生中柱。中国首次发现和报道的桫椤科茎干化石(可归入Cyathocaulis或Cibotiocaulis), 采集自松辽盆地白垩纪地层, 保存有髓和皮层, 其中分布有丰富的维管束和不定根, 不定根由中央的二极型初生木质部及周围的厚壁组织等组成(程业明和刘凤香, 2017); 同一产地, 还发现可能属于桫椤目的Tempskya zhangii, 其树干由许多纤细的茎、叶柄基和周围大量紧密排列的不定根组成, 构成一种由茎、叶柄基和根缠绕在一起的假树干。Tempskya zhangii的不定根直径0.38~1.5 mm, 不定根的木质部由内厚壁组织和外薄壁组织皮层包围; 根从茎上水平长出, 但很快就向下延伸, 与其他的根和茎相互交织(Yang et al., 2018)。
水龙骨目是薄囊蕨类的优势类型, 占现生真蕨类植物90%的种类(陆树刚, 2007)。但是, 在真蕨类植物的系统描述中, 大多仅涉及根状茎的信息, 而很少提及枝生根的特征, 因此很难进行深入的对比分析。总体上, 该目植物具不同形态的根状茎, 其上着生枝生根。文中依据陆树刚(2017)发表的90幅显示了根状茎及枝生根的图片, 对少数几个科中的一些属种的特征简述如下。Arachniodes reducta(缩羽复叶耳蕨)属鳞毛蕨科, 其根状茎直立、斜生或长而横走, 鳞片大, 披针形至卵形, 枝生根密度大, 较粗, 可明显观察到侧根。Dryopteris panda(大果鳞毛蕨)属鳞毛蕨科, 与缩羽复叶耳蕨相比, 根状茎短小、粗壮, 枝生根密度大, 偶见侧根。Lomagramma mattewii(网藤蕨)属藤蕨科, 根状茎攀附于树干上, 腹背压扁, 腹面生根, 背面生叶, 叶柄基部下延与根状茎而形成棱脊, 被黑色鳞片, 披针形, 边缘具睫毛。Nephrolepis auriculata(肾蕨)属肾蕨科, 根状茎短而直立。Oleandra wallichii(高山条蕨)属条蕨科, 根状茎长而横走或直立, 多分枝, 密被鳞片, 枝生根坚硬细长。Phymatosorus cuspidatus(光亮瘤蕨)属于水龙骨科, 是附生或土生植物, 根状茎横走至横卧, 被鳞片, 鳞片盾状着生。与耳蕨粗壮的根状茎相比, 光亮瘤蕨的枝生根较细, 可见侧根。Pseudocyclosorus repens(长根假毛蕨)属金星蕨科, 根状茎直立、斜生或长而横走, 被毛和鳞片, 通常较为粗壮, 枝生根密度大, 较粗, 可明显观察到侧根。
化石记录方面, 邓胜徽(1995)报道了中国东北地区早白垩世地层中的Coniopteris concinna(雅致锥叶蕨)的匍匐状根状茎, 其直径约1.0 cm, 但不清楚是否具枝生根。Li等(2020)的最新研究认为Coniopteris与水龙骨目的亲缘关系更近(尤其是现生碗蕨属Dennstaedtia), 因此提议将Coniopteris归于水龙骨目的基部类群。
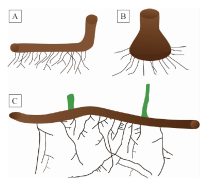
真蕨类植物的根系具有一定的演化保守性, 均为枝生根(图 6)。在早期真蕨类中, 枝生根着生于茎轴、或推测的根状茎、或主干膨大的基部上(图 6-A, 6-B)。所谓的根状茎通常在形态学上与气生茎无明显区别, 一些体现为主轴的下延部分, 一些呈Z字型弯曲(反映其可能并非直立生长), 事实上, 在化石中仅能合理地推测一些具根的茎轴是否为根状茎、是否接触基质或是否匍匐生长(Cornet et al., 1976; Scheckler, 1986; Wang and Geng, 1997; Xue and Hao, 2008; Wang et al., 2015; Xue and Basinger, 2016)。在真蕨纲冠群中, 根状茎通常在地下匍匐生长。根据一些测量数据可以看出, 枝生根的长度、宽度, 在一些早期类群如Denglongia、Melvillipteris以及现生里白科、紫萁科如Dicranopteris、Osmunda等中, 并没有显著变化(见前述)。但树蕨状植物如Eospermatopteris等的植株体较为高大, 其根系也较为粗壮。
 | 图 6 真蕨类植物的典型根系形态示意图 A— 早期真蕨类植物根系的典型形态, 示根状茎和细密的枝生根, 后者偶见二歧式分枝; B— 早期树蕨状真蕨类Eospermatopteris, 示基部膨大的主干, 以及其上辐射着生的枝生根, 与A相似, 根极少分枝; C— 现生真蕨类植物根系的典型形态, 示根状茎、枝生根以及不同级别的侧根, 向上的绿色线条指代叶轴的基部Fig.6 Diagrams showing typical root morphologies of ferns |
依据前文资料, 笔者对真蕨类植物根系的形态演化趋势进行初步总结:
在真蕨类植物演化出现之前, 原始的维管植物如莱尼蕨纲的根系并非真正的根, 而是具假根根系(rhizoid-based rooting systems), 代表植物如下泥盆统莱尼燧石层中的Rhynia。Rhynia不具真正的根状茎, 只有圆柱形的、匍匐的轴分布在基质表面或浅层, 起着根状茎的作用。在这些似根状茎的表面, 可见纤弱的、单细胞的假根(rhizoids, 图 7-A1, 7-A2)(Taylor et al., 2009; Kenrick and Stullu-Derrien, 2014)。具假根根系演化出现于真正的根之前, 可能来源于绿藻类祖先(Kenrick and Stullu-Derrien, 2014)。
 | 图 7 真蕨类植物根与茎— 叶系统的形态演化示意图 A— 真蕨类出现之前的原始维管植物如Rhynia、Aglaophyton等(祖先类型):A1— 三维的、等二歧式分枝构成的顶枝束及匍匐轴; A2— 具假根的匍匐轴, 方框中为放大的假根示意图(比例尺200 μ m); A3— 下泥盆统莱尼燧石层中的代表植物Rhynia gwynne-vanghanii的复原图(引自Edwards, 2004)。B— 以灯笼蕨Denglongia为例, 示早期真蕨类的形态特征:B1— 主轴, 侧枝(由顶枝束的越顶生长形成)及侧枝上的“ 小羽片” (平面化的顶枝束); B2— 根状茎及枝生根(另见图 6-A), 与茎— 叶系统相对应, 这种原始的根可视为不分枝的或偶见二歧式分枝的顶枝束; B3— 灯笼蕨的整体形态复原图(引自Xue and Hao, 2008)。C— 真蕨纲冠群代表类群的形态特征:C1— 羽状复叶(以化石类群Cladophlebis为例); C2— 真蕨类冠群植物的根状茎、枝生根及侧根(另见图 6-C); C3— 典型真蕨类冠群植物的整体复原 (以Cladophlebis为例)Fig.7 Diagrams showing morphological evolution of roots and stem-leaf systems of ferns |
中、晚泥盆世的早期真蕨类植物(如Denglongia、Melvillipteris、Shougangia等)具有直径约1.0 mm的密集枝生根, 且多见于茎轴或根状茎的一侧。在迄今报道的早期真蕨类中, 仅Denglongia的枝生根可见等二歧式分枝, 且仅可见分叉1次, 在其他属种的大量已描述的标本中, 枝生根均未见到分枝, 可能说明早期真蕨类的根系分枝罕见, 若有的话, 也仅为等二歧式分枝(图 6-A)。演化至真蕨纲冠群植物, 其根状茎呈现多种形态, 如直立、斜生和横卧等。枝生根密集或稀疏, 侧根发育, 即根系呈单轴式分枝, 侧根可达2级甚至更多(图 6-C)。
植物组织结构的进化及功能体现出模块化的模式, 即茎— 叶系统和根系统是相对独立的模块(Xia et al., 2010; Ma et al., 2018)。对于大型叶植物如真蕨类茎— 叶系统的演化, 前人多以德国学者Walter Zimmermann于1930年提出的顶枝学说进行解释(Beerling and Fleming, 2007), 体现出3个阶段: (1)早期维管植物如莱尼蕨纲中的Rhynia、Aglaophyton具原始的三维顶枝束(图 7-A)(Taylor et al., 2009), 在真叶植物基部类群中, 开始具多样化的枝— 叶复合体(branch-leaf complexes)(Hao and Xue, 2013); (2)通过顶枝束的越顶(overtopping)、平面化(planation)及部分的蹼化(webbing), 在早期真蕨类植物中, 已具有固定的分枝样式, 侧枝束或枝— 叶复合体逐渐平面化(图 7-B), 甚至在一些类群如Shougangia、Rhacophyton中已发生蹼化, 形成最早的复叶(Wang et al., 2015); (3)真蕨纲冠群中, 顶枝束完全平面化及蹼化, 真正的羽轴及叶片形成(图 7-C)(Taylor et al., 2009)。
与上述茎— 叶系统的演化相对应, 真蕨类根系统的形态演化也可采用顶枝学说进行解释。早期真蕨类的根可视为不分枝的或偶见二歧式分枝的顶枝束(图 7-B2), 是真蕨类根系的(最)原始状态; 经历分枝复杂化、顶枝束的越顶生长, 侧根系统发达的根系在真蕨纲冠群中演化出现(图 7-C2)。但是, 从形态学角度可以看出, 根并未进一步发展至平面化、蹼化阶段。
真蕨类植物的根和叶子的不同演化趋势, 与它们执行的不同功能及面临的不同选择压力有关。植物具有光合作用的能力, 影响光合作用的主要因素是光照强度, 薄而扁平的叶片具有较大的叶表面积, 且叶表面积越大, 则受光面积越大。当顶枝束经历越顶、平面化及蹼化过程演化成真正的叶子之后, 可以将光截获强度最大化, 也可能提供更好的机械稳定性(Beerling and Fleming, 2007)。另外, 由于真蕨类的孢子囊群多与叶子相关, 因此平面化、蹼化的叶性器官, 也可能对促进孢子散布有益。
根的主要功能为吸收水分和营养物质。维持植物生存的另一个不可或缺的生理过程是蒸腾作用, 主要依靠植物根系从土体中吸收水分后, 由叶片气孔和皮孔散发到大气中。因此, 植物根系在演化过程中的选择压力在于如何扩大其表面积使尽可能接触到更多的土壤基质, 以及如何通过分枝、扩展至更多的生境中去。在宏观尺度上, 大多真蕨类具密集的、纤细的枝生根, 具有很高的更新速率(与粗大的植物根系相比), 可以更有效地利用光合作用产物在土壤中扩展(Ma et al., 2018)。对于真蕨纲冠群, 侧根系统的发育使得植物体可以在较浅的基质中实现其根系密度的最大化; 而早期真蕨类不分枝或偶见二歧式分枝的根, 在土壤空间的利用效率上较为有限。在微观尺度上, 原始的维管植物, 如Rhynia等的似根状茎上着生的假根以及在后期的维管植物根系上的表皮细胞突起(即根毛, root hairs), 均可提高根系的表面积, 以利于水分和营养物质的吸收(Jones and Dolan, 2012; Shekhar et al., 2019)。根毛在化石记录中非常罕见, 因此未列入本文的讨论范围。
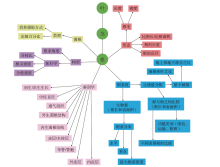
长期以来, 人们对植物化石的研究多侧重于茎— 叶系统, 而对根系统的研究相对有限, 这一方面是由于植物的根系统很难完整地保存, 另一方面是因为根系统的特征对分类学、系统学解释的贡献似乎不大(但事实上与研究程度不够有关)。迄今为止, 国内外对真蕨类根系统的研究集中在中生代, 尤其是紫萁科的矿化根状茎。McCormack等(2017)总结了现生植物根系统的大量性状(图 8), 其中的一部分性状可以在植物化石中得以识别。对于压型保存的根, 其构型、形态、根系统生物量等几个方面的信息, 如根的长度、直径、分枝密度和分枝角度等性状都可以在化石中得以测量, 可为不同类群之间的对比提供定量数据。根的构型指的是同一根系中不同类型的根或不定根在生长介质中的空间造型和分布(Lynch, 1995)。
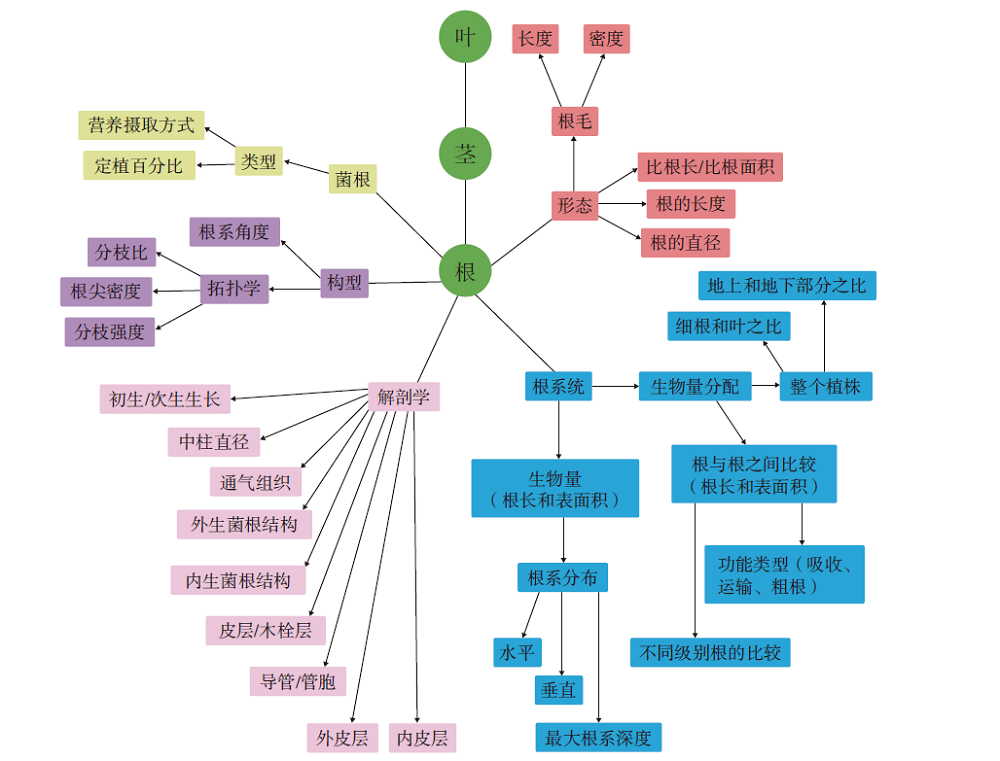 | 图 8 化石记录中可识别的根系性状示意图(修改自McCormack et al., 2017)Fig.8 Diagram showing root features that are recognizable in fossil record(modified from McCormack et al., 2017) |
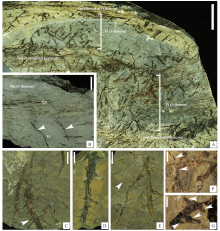
笔者对北京西山中侏罗统上窑坡组的植物根化石进行了研究, 识别出2个类型的真蕨类根系, 均由大量密集的枝生根和细侧根组成(Liu et al., 2018)。第1种根系中, 密集且纤细的枝生根着生在根状茎上, 平均长度约9.9 mm, 宽度约1.1 mm(N=84), 在枝生根上可见直径小于0.2 mm的侧根(图 9-A)。这种根系统与以Cladophlebis(枝脉蕨)为主导的植物群落相关, 其中最为丰富的是Cladophlebis cf. scariosa(膜质枝脉蕨相似种)和Cladophlebis delicatula(纤柔枝脉蕨)。在第2种根系中, 枝生根长可达22 mm, 直径为0.2~1.7 mm, 侧根平均直径约0.2 mm(图 9-C至9-G)。在度量上, 这2种根系与一些现生真蕨类植物如Dicranopteris(芒萁属)、Gleichenia(里白属)的根系较为一致。
 | 图 9 北京西山中侏罗世地层中的真蕨类植物根系及古土壤 A— 垂向剖面显示: 具丰富植物碎屑的细粒岩相(Fpd), 具原位根系的细粒岩相(Fr), Fpd可解释为改造过的土壤表层(O层), Fr可解释为土壤A层, 标本号: PKUB16258, 比例尺: 50 mm; B— 示1个自下而上的Fr-Fpd-Fnp序列, 箭头指示垂直生长的根状茎, 标本号: PKUB16282, 比例尺: 10 mm; C至E— 示另一种根系, 垂直生长的根状茎上着生细的枝生根, 枝生根上发育侧根。C标本号: PKUB16292, 比例尺: 10 mm; D标本号: PKUB16293, 比例尺: 10 mm; E标本号: PKUB16295-1, 比例尺: 5 mm; F— 放大的枝生根(E图箭头所指处), 箭头指示侧根, 比例尺: 1 mm; G— 枝生根和侧根的放大(C图箭头所指处), 箭头指示侧根, 比例尺: 1 mm。标本首次发表于Liu等(2018), 收藏于北京大学地质学系Fig.9 Fern roots and paleosols from the Middle Jurassic of Western Hills, Beijing |
化石根系统的识别可为探讨植物根系的演化提供依据。同时, 由于根大多为原位保存, 因此也可为地史时期的植物— 土壤系统研究提供依据。笔者依据上窑坡组植物根化石的研究以及其赋存层位的沉积学分析, 识别出3种不同的岩相(图 9-A, 9-B): (1)具有广泛的垂直根状茎和枝生根的细粒岩相(Fr); (2)含丰富植物叶片化石的细粒岩相(Fpd), 通常与Fr相形成层耦, Fpd岩相中积累的植物碎屑被浅水水流所改造; (3)主要由碎屑沉积物组成、没有或少见植物碎屑的细粒岩相(Fnp)。
根据岩相特征及其中的植物根系、叶片化石的保存状态, 笔者认为上窑坡组中的Fpd-Fr层耦代表了植物影响下的土壤发育过程:可以将Fpd解释为改造过的土壤O层, 即以有机质堆积为主的层位; 将Fr解释为 A层, 即有机质与矿物成分混合的层位, 位于O层之下; Fnp相中没有或很少有植物碎屑, 相当于C层, 即代表几乎没有改变和风化的母岩, 表明植被发育的终止。根系统的母体植物极可能属于Cladophlebis, 它们为半水生或湿地殖民者, 其根和根状茎由于浸水、埋藏使得腐蚀作用受到抑制, 从而留下大量原位保存的化石(Fr); 相比之下, 植物的茎叶组织在水流的侵蚀和改造下, 虽然仍在其生境埋藏, 但保存较为破碎(Fpd)。多个土壤的加积过程, 形成更厚的序列如Fr-Fpd-Fnp-Fr-Fpd累积序列, 这个序列反映了植被发育(Fr-Fpd)、碎屑物堆积(Fnp)、植被再次发育(Fr-Fpd)的动态过程。由于这些序列的成土改造作用微弱, 因此可归入新成土纲。原位保存的植物根系是鉴定埋藏古土壤的重要依据, 因为这些根系无疑代表了当时陆地表面的植被发育。
1)真蕨类植物的根系具有一定的演化保守性, 均为枝生根。在早期真蕨类如Denglongia、Shougangia、Eospermatopteris等植物中, 密集的枝生根着生于茎轴、或推测的根状茎、或主干膨大的基部上。根系的分枝罕见, 仅在少数标本中表现为等二歧式分枝。
2)在真蕨纲冠群植物中, 根状茎呈现直立、斜生和横卧等多种形态, 枝生根密集或稀疏, 侧根发育, 即根系呈单轴式分枝, 侧根可达2级甚至更多。
3)真蕨类根系统的形态演化可采用“ 顶枝学说” 进行解释: 与植物的茎— 叶系统相对应, 早期真蕨类的根可视为不分枝的或偶见二歧式分枝的顶枝束, 是真蕨类根系的(最)原始状态; 经历分枝复杂化、顶枝束的越顶生长, 侧根系统在真蕨纲冠群中演化出现。但是, 与大型叶的演化不同, 根的进化并未进一步发展至平面化、蹼化阶段。
4)真蕨类以及其他植物类群的根系的构型、形态、结构、生物量分配等信息可在化石记录中得以不同程度的识别和揭示, 这些信息可为探讨植物根系的演化及研究地史时期的植物— 土壤系统提供重要依据。
真蕨类植物具有非常丰富的多样性, 上述认识是笔者基于少数代表性类群、属种所得出的, 因此难免有管中窥豹的局限性。期待未来的深入研究、特别是解剖学等方面资料的总结分析, 能够为进一步理解真蕨类植物根系进化提供新的启示。
致谢 感谢两位审稿专家提出的建设性意见和建议, 感谢云南大学陆树刚教授的有益讨论。北京大学造山带与地壳演化教育部重点实验室倪德宝老师和贾秋月老师制作岩石薄片, 常洁琼博士绘制了图7-C3复原图, 在此一并感谢!
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|