



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
贵州震旦系陡山沱组磷块岩成磷作用及与新元古代末期氧化事件(NOE)的耦合*
[张亚冠1, 2  , 杜远生
, 杜远生2, 3 , 刘建中2, 4 , 王泽鹏2, 5 , 邓超6 ]
, 杜远生, 刘建中|
|

通讯作者简介 杜远生,男, 1958年生,中国地质大学(武汉)教授、博士生导师,主要从事沉积地质与沉积矿产研究。 E-mail: duyuansheng126@126.com。
第一作者简介 张亚冠,男, 1990年生,中国地质大学(武汉)博士后,主要从事沉积地质与沉积矿产研究。 E-mail: zyg1000800@sina.cn。
新元古代末期历经“雪球地球”和大气、海洋增氧事件后,发生了全球性成磷事件,贵州省震旦系陡山沱组大规模磷矿床沉积是本次成磷事件的典型代表。然而现阶段对贵州陡山沱组磷块岩成磷作用机制研究存在诸多争议,成磷事件与同期古海洋环境转变之间的关联研究也较为薄弱。作者以贵州省瓮安、遵义和丹寨地区陡山沱组原生磷块岩为研究对象开展的沉积学、岩石学、矿物学和地球化学特征研究表明: 陡山沱早期成磷作用被限制在浅水海岸,瓮安地区 A矿层球粒磷块岩中大量的含铁自生矿物黄铁矿、海绿石以及无 Ce负异常指示的贫氧沉积水体环境均表明, Fe-氧化还原泵成磷模式在富磷过程中发挥了重要作用;陡山沱晚期磷块岩分布扩散至较深水陆棚—斜坡沉积相区,磷块岩与富有机质岩层共生,矿物晶体形态特征与矿石内富含的大量生物化石均表明,有机质沉降聚磷作用和生物成磷作用促使磷块岩大量沉积,较明显的 Ce负异常值也指示了海水氧气含量的提升。成磷作用模式的转变和磷块岩分布的扩展是对海洋增氧事件的沉积响应,同时造成的多细胞动物演化也影响了深部水体的氧化还原状态,进而反映了贵州省陡山沱组磷块岩大规模沉积与新元古代末期氧化事件( NOE)密切的耦合关系。
About the corresponding author Du Yuan-Sheng,born in 1958,professor and Ph.D. supervisor of China University of Geosciences(Wuhan),is mainly engaged in sedimentary geology and sedimentary minerals. E-mail: duyuansheng126@126.com.
About the first author Zhang Ya-Guan,born in 1990,postdoctoral fellow at China University of Geosciences(Wuhan),is mainly engaged in sedimentary geology and sedimentary minerals. E-mail: zyg1000800@sina.cn.
Phosphate deposits of the Ediacaran Doushantuo Formation accumulated in the central Guizhou Province are the typical phosphate-rich sediments during the Neoproterozoic Phosphogenic Episodes,which occurred after the “Snowball Earth”period and Neoproterozoic Oxygenation Event. However,the mechanisms of phosphate enrichment in seawater have always been hotly debated,and the research for correlations between phosphogenesis and transition of Ediacaran palaeo-ocean environments is still unsubstantial. This study focused on the sedimentological,petrological,mineralogical and geochemical analyses on the Doushantuo pristine phosphorite in Weng'an,Zunyi and Danzhai area. Documented by spherulitic phosphorites in the Lower Phosphorite beds from Weng'an area that contain abundant autogenetic Fe-bearing minerals such as pyrite and glauconite and show weak Ce negative anomaly,Fe-redox pumping in low-oxygen environments are the important phosphogenesis mechanism but only limited in coastal waters in the Early Doushantuo Period. Phosphorite in the Upper Phosphorite beds deposited within organic-rich beds and contain massive biological fossils suggests that phosphogenesis might have been triggered by degradation of organic matter and biological action in the Late Doushantuo Period,and distribution of phosphatic sediments extended to the deeper shelf to slope setting. Obvious Ce negative anomaly implies the increase of oxygen content in seawater. The transition of phosphogenesis mechanisms and the expansion of phosphorite deposits are the sedimentary response of ocean oxygenation,and the associated evolution of metazoans also changed the redox conditions of the deep seawater. These sedimentary and geochemistry data reflect the closed coupling relation between Doushantuo phosphorite in Guizhou Province and the Neoproterozoic Oxygenation Event.
在新元古代末期全球性成磷事件背景下, 扬子地块震旦系(埃迪卡拉系)陡山沱组广泛发育了富磷沉积物, 这一时期所形成的磷矿床不仅资源储量丰富, 而且保存了丰富的古生物化石, 具有极其重要的科学研究和经济利用价值(叶连俊等, 1989; 吴祥和等, 2000)。贵州省陡山沱组大规模磷矿床沉积是新元古代末期成磷事件的典型代表, 然而由于复杂沉积环境下不同类型磷块岩矿石成因存在多样性, 关于成磷作用机制的研究仍存在不同的观点。最早提出的“ 生物成磷说” 认为磷块岩是生物在生长过程中通过不断的捕获、粘结海洋中分散的磷酸盐并最终形成生物遗体堆积而成(王砚耕和朱士兴, 1984; 叶连俊等, 1989)。黔中地区广泛发育的叠层石磷块岩和瓮福磷矿B矿层中的含“ 瓮安生物群” 化石磷块岩均对生物成磷作用提供了很好的证据支撑(Chen et al., 2014; 张伟等, 2015)。虽然生物成磷作用可能是磷矿层沉积的重要途径, 但是“ 生物成磷说” 难以解释无生物结构的原生磷块岩沉积模式, 生物作用规模也不足以形成超大型磷矿床沉积(密文天等, 2010; 张亚冠等, 2016)。伴随对大洋磷循环系统的深入研究, “ 上升洋流成磷说” 得到了越来越多的学者认可, 即上升洋流携带深部富磷海水进入浅水海岸, 刺激表层海水大洋生产力, 促使藻类生物的生长繁盛, 藻类生物死亡后以有机质形式携带大量磷质沉降, 在硫酸盐还原细菌等微生物作用下使有机质迅速释放磷酸盐并形成磷块岩的大规模沉积(郭庆军等, 2003; 牟南和吴朝东, 2005; 聂文明等, 2006; 林丽等, 2010; 密文天等, 2010, 2013; 陈国勇等, 2015; 吴文明等, 2017; 张亚冠等, 2019; Zhang et al., 2019)。然而在震旦纪初期古海洋中极低含量的硫酸盐浓度能否促使有机质有效降解释放磷酸盐仍存在很大的争议(Poulton and Canfield, 2006; 肖朝益等, 2018)。考虑到震旦纪之前整体海洋为铁化缺氧状态, Fe-氢氧化物在氧化还原界面附近不断释放出吸附的磷酸盐, 也可以使孔隙水中的磷不断富集形成磷块岩沉积, Fe-氧化还原泵成磷模式可能在富磷过程中发挥了重要作用(She et al., 2014; Cui et al., 2016; 肖朝益等, 2018; Algabri et al., 2020)。
此外, 磷作为控制大洋生产力的限制性营养元素, 其沉积成矿作用是地球表层系统大气圈、水圈、生物圈和沉积圈相互作用的产物, 海洋的磷块岩大规模沉积往往蕴含着全球系统突变的关键信息(Fö llmi, 1996; Filippelli, 2008, 2011)。新元古代末期是地质历史时期的重要转折, 近年来越来越多的生物地球化学研究表明, 新元古代末期全球性成磷事件, 与大陆构造演化、气候变化、大气与海洋增氧和海洋生态系统演化均存在密切的相互制约和耦合关系(Nelson et al., 2010; Pufahl and Hiatt, 2012; Hiatt et al., 2015; Cui et al., 2016; Caird et al., 2017; 肖朝益等, 2018; 纪秋梅等, 2019; Zhang et al., 2019), 然而现阶段对华南震旦系陡山沱组磷矿成磷作用与深时全球变化的响应机制研究刚刚起步, 特别是初始成磷作用与同期古海洋地球化学环境转变之间的关联研究较为薄弱。贵州震旦系陡山沱组磷矿层时空展布完整连续, 且磷矿层内蕴含了以“ 瓮安生物群” 为主的丰富化石保存, 不仅记录了成磷事件进程与生命演化发展, 而且反映了古海洋环境的转变, 是研究新元古代末期成磷事件及其古海洋地球化学状态的理想材料。针对瓮安、遵义和丹寨地区陡山沱组磷块岩开展岩石学、矿物学、沉积学和沉积地球化学研究, 解析复杂沉积环境下不同类型原生磷块岩成磷作用过程, 并利用磷在氧化还原界面附近富集成矿的沉积属性, 探讨成磷作用与海洋增氧、生命演化之间的耦合机制, 为探索新元古代末期古海洋环境提供新窗口。
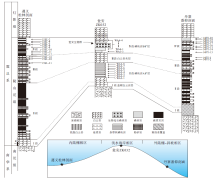
震旦系陡山沱组是新元古代末期雪球事件后, 广泛发育在扬子地块的一套碎屑岩、碳酸盐岩和磷块岩混合沉积序列, 底部盖帽白云岩直接覆盖于南华纪南沱组冰碛砾岩之上, 并与上覆灯影组白云岩沉积地层呈整合接触(Zhu et al., 2007; Jiang et al., 2011; 杨爱华等, 2015), 其沉积时限为635— 551 Ma(Condon et al., 2005)。陡山沱组沉积期扬子地块保持古陆或隆起环绕的浅水海岸、内陆棚和外陆棚相沉积古地理格局, 并向南东方向延伸相变为斜坡— 深水盆地沉积相区(图 1)(Vernhet, 2007; Zhu et al., 2007; Vernhet and Reijmer, 2010; Jiang et al., 2011; 刘静江等, 2015)。陡山沱组在贵州省内除黔西南外的大部分地区均有出露, 其中在黔中古陆北缘和东缘浅水海岸沉积相区的开阳、瓮福(瓮安和福泉)地区形成了大规模磷矿床沉积, 矿石类型以碎屑状、球粒状和含生物化石磷块岩为主; 位于黔北内陆棚相区的遵义地区和黔东南外陆棚— 斜坡相区的丹寨地区陡山沱组地层岩相组合以泥页岩、粉砂岩和泥晶白云岩为主, 磷块岩通常以泥晶磷块岩夹层或磷质结核、透镜体形式散布于地层中(叶连俊等, 1989; 吴祥和等, 2000; 张亚冠等, 2019)。
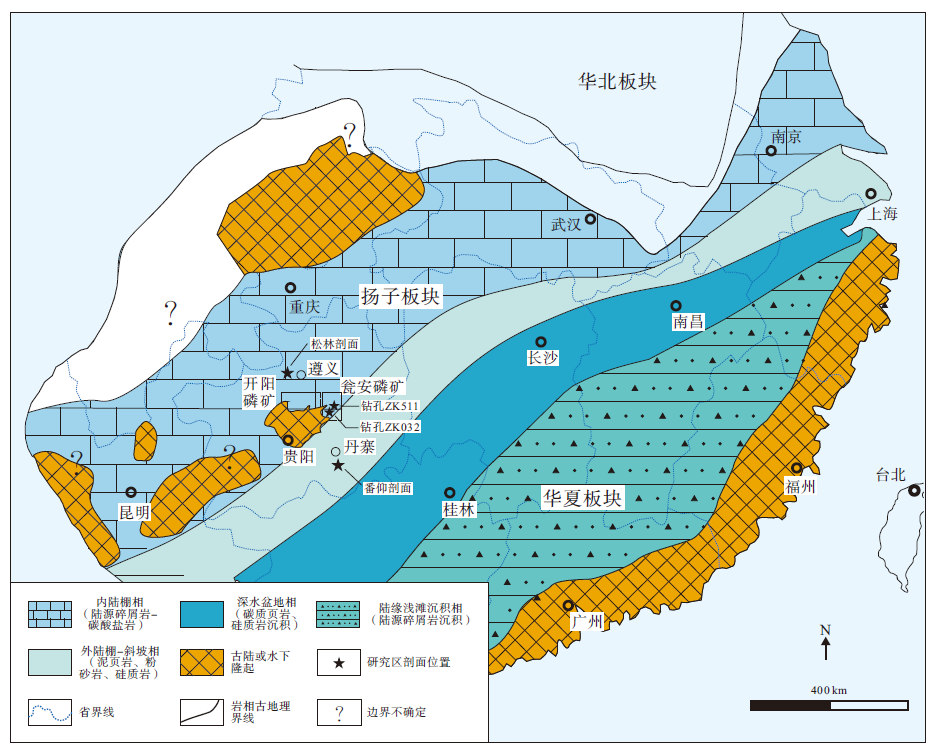 | 图 1 扬子地块震旦系陡山沱组沉积期岩相古地理图与研究区剖面位置(据Zhang et al., 2019; 有修改)Fig.1 Palaeogeographic map of Yangtze Block during the Sinian Doushantuo Period and profile locations of study area (modified from Zhang et al., 2019) |
瓮福地区陡山沱组假整合于南沱组冰碛砾岩之上或超覆于青白口系之上, 层内发育A、B共2个磷矿层, 中间由1层白云岩夹层相隔(图 2)。瓮安ZK032钻孔剖面陡山沱组地层底部发育厚约3.5 m的盖帽白云岩层, 层内可见帐篷构造、重晶石扇等; A矿层底段为含磷白云岩, 中上段发育含白云质条带、纹层状砂屑或球粒磷块岩, 条带由砂屑、球粒磷块岩条带和细晶白云石条带相间组成, 总厚度约15 m; A矿层之上夹层为灰色、深灰色厚层细晶白云岩, 顶部可见溶蚀孔洞等暴露构造, 厚约6 m; B矿层下段为黑色碳质泥晶磷块岩层, 中上段发育白云质磷块岩、碎屑状磷块岩和含生物化石磷块岩, 顶段发育含磷白云岩, 并再次出现暴露不整合面, 总厚度约11 m。
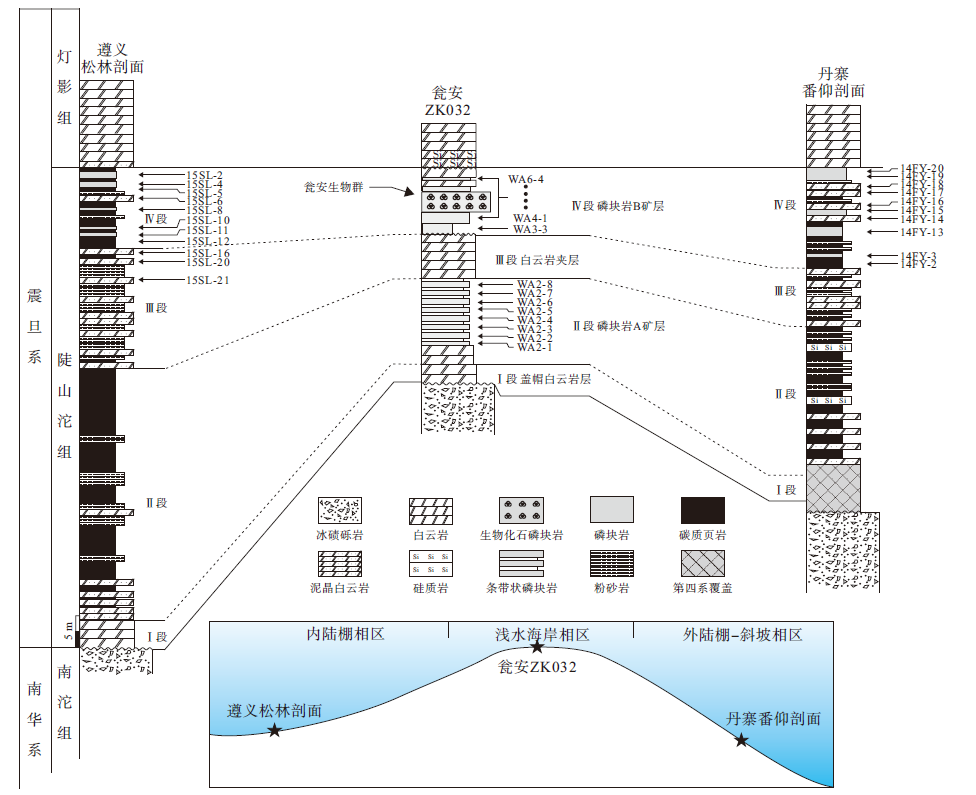 | 图 2 贵州省遵义、瓮安和丹寨地区震旦系陡山沱组地层剖面柱状对比图及采样位置Fig.2 Stratigraphic correlation of the Sinian Doushantuo Formation and sampling locations in Zunyi, Weng'an and Danzhai areas, Guizhou Province |
遵义松林陡山沱组剖面和丹寨番仰剖面陡山沱组地层自下而上均发育可与瓮安地区对比的四段沉积层序(图 2)。遵义地区第Ⅰ 段为盖帽白云岩沉积层(厚4 m); 第Ⅱ 段底部发育中薄层泥晶白云岩层, 并向上相变为灰黑色薄层粉砂质页岩、碳质页岩及少量泥晶白云岩(厚约47 m); 第Ⅲ 段为灰绿色薄层粉砂质泥岩与中薄层泥晶白云岩互层, 顶部出现磷质结核(厚23 m); 第Ⅳ 段为灰黑色薄层碳质页岩与泥晶白云岩夹中薄层泥晶磷块岩、磷质页岩(厚16 m)。丹寨地区底部第Ⅰ 段地层缺失, 第Ⅱ 段中下段为黑色薄层页岩与泥晶白云岩互层或夹层, 上段为中薄层硅质页岩、黑色页岩、粉砂岩夹硅质岩(厚度大于19 m); 第Ⅲ 段为灰色泥晶白云岩夹薄层黑色碳质页岩、粉砂岩(厚17 m); 第Ⅳ 段发育灰黑色中薄层泥晶磷块岩矿层, 夹薄层碳质页岩、磷质粉砂岩和泥晶白云岩, 可见滑塌构造和包卷层理(厚约17 m)。
本研究所采集岩相学和地球化学分析样品为瓮安ZK032钻孔剖面、遵义松林剖面和丹寨番仰剖面陡山沱组地层按一定的顺序取样所得, 具体采样层位与样品岩性见图 2和表 1、表 2, 另有部分岩石薄片样品采自瓮安ZK511钻孔。岩相学分析在中国地质大学(武汉)生物地质与环境地质国家重点实验室完成, 首先选用尼康LV100POL型号光学显微镜对样品岩石薄片进行观察, 并进一步采用SU8010型号高分辨率场发射扫描电子显微镜(SEM)观测分析磷块岩样品微观形貌特征和矿物组成, 测试前将样品切割为长3~5 mm、厚1~3 mm的岩块, 并保留一面为自然破裂断口面, 岩矿表面镀金膜后放置样品台, 分别在7.5 kV和15 kV加速电压下进行二次电子成像(SE)和背散射成像(BSE), 并利用能谱分析(EDS)对矿物成分进行半定量测试分析。稀土元素测试在澳实分析检测(广州)有限公司实验室采用电感耦合等离子体质谱仪(ICP-MS, ME-MS82)完成, 首先通过镜下观测挑选原生沉积磷块岩样品和白云岩样品并进行破碎处理, 选取新鲜无次生脉体岩块并粉碎至小于200目的粉末, 之后将粉末样品加入到偏硼酸锂/四硼酸锂溶剂中混合均匀, 在1000 ℃以上的熔炉中融化, 待冷却后用硝酸、盐酸和氢氟酸定容, 再用等离子体质谱仪分析, 测试标准偏差低于5%。为避免La富集和Gd富集造成的对Ce异常和Eu异常值计算失真, 使用了Lawrence 等(2006)提出的Ce异常和Eu异常计算方法:
Ce/Ce* =CeN/(P
Eu/Eu* =EuN/( ⅔SmN+1/3TbN)
| 表 1 瓮安ZK032钻孔陡山沱组磷块岩与磷质白云岩样品部分主量元素、稀土元素及Ce* 、Eu* 、DyN/SmN数据 Table1 Data of partial major elements, REEs, Ce* , Eu* and DyN/SmN of Doushantuo phosphorites and phosphatic dolostones in Weng'an core ZK032 |
| 表 2 遵义松林、丹寨番仰陡山沱组剖面磷块岩、磷质页岩与磷质白云岩样品部分主量元素、稀土元素及Ce* 、Eu* 、DyN/SmN数据 Table2 Data of partial major elements, REEs, Ce* , Eu* and DyN/SmN of Doushantuo phosphorites, phosphatic shales and phosphatic dolostones in Zunyi and Danzhai areas |
其中, N为澳大利亚后太古代页岩标准化数值。
陡山沱组磷块岩成因类型复杂, 在不同沉积环境下磷块岩形态特征各不相同。瓮安地区A矿层主要由含白云质条带的球粒状磷块岩和碎屑状磷块岩组成, B矿层则存在泥晶磷块岩、碎屑状磷块岩、含生物化石磷块岩等多种矿石类型。遵义松林和丹寨番仰地区磷块岩类型则相对单一, 主要以泥晶磷块岩为主。为排除后期改造作用对磷块岩原生沉积环境造成的影响, 本次研究主要选取原生成因类型磷块岩进行重点分析, 包括球粒状磷块岩、泥晶磷块岩和含生物化石磷块岩。
3.1.1 球粒状磷块岩
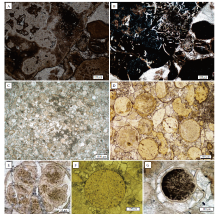
球粒状磷块岩手标本下呈浅灰色— 灰黑色颗粒状, 可见白云石条带或纹层(图 3-A, 3-B), 常称为假鲕状磷块岩。矿石由卵圆状、浑圆状磷质颗粒组成, 颗粒含量通常大于50%, 粒径小于1 mm, 大多数颗粒集中在0.1~0.2 mm之间; 镜下颗粒内因混入较多有机质成分和残余黄铁矿而呈现不透明黑色, 可见球粒外包壳结构和球粒内似核形石状圈层构造(图 3-C); 颗粒间胶结物多为碳酸盐矿物, 主要为微晶白云石颗粒(图 3-D); 此外矿石内含少量石英碎屑和海绿石颗粒(图 3-E), 磷质球粒和海绿石颗粒内出现存在普遍的黄铁矿聚集(图 3-E, 3-F)。含白云石条带球粒磷块岩主要由磷质球粒定向排布组成的磷质条带和微晶白云石条带或纹层构成, 白云石条带或纹层内同样含少量磷质球粒(图 3-D)。扫描电镜和能谱分析研究表明, 球粒状磷块岩主要由磷质球粒和白云石胶结物组成(图 3-G, 3-H, 3-I, 3-J), 磷质球粒内部可见由2~50 μ m大小不一的多面体形态黄铁矿晶体组成的黄铁矿莓状体(图 3-K); 超微晶体形态显示球粒状磷质颗粒内部由长度0.5~2 μ m的长柱状磷灰石晶体呈同心放射状排布(图 3-L), 表现出花环状磷灰石晶体集合体的形态特征。
 | 图 3 瓮安陡山沱组球粒磷块岩矿石结构特征 A、B— 瓮安陡山沱组A矿层含白云质条带/纹层球粒磷块岩; C— 球粒状磷块岩中磷质球粒含大量有机质, 单偏光; D— 球粒状磷块岩中磷质条带/纹层与白云质条带/纹层, 磷质条带/纹层由磷质球粒组成, 白云质条带/纹层由泥晶白云石组成, 正交偏光; E— 磷质球粒间的自生海绿石矿物, 单偏光; F— 磷质球粒内莓状黄铁矿, 单偏光; G— 球粒状磷块岩扫描电镜照片, 背散射成像; H、I、J— 图G打点位置相关矿物能谱曲线; K— 球粒状磷块岩中莓状黄铁矿簇, 背散射成像; L— 磷质球粒内磷灰石晶体超显微结构形态, 二次电子成像Fig.3 Structural characteristics of spherulitic phosphorite of the Doushantuo Formation in Weng'an area |
3.1.2 泥晶磷块岩与磷质页岩
泥晶磷块岩一般为深灰色至黑色, 矿石致密、均匀, 矿层呈薄层或中薄层产出(图 4-A, 4-B, 4-C), 通常与含碳质泥岩、页岩或粉砂岩共生, 岩层内常见水平纹层(图 4-D), 此外矿层内常见半自形— 自形黄铁矿。泥晶磷块岩中磷灰石产出形态可分为2种: 一种作为石英或泥质碎屑的基质成分呈不规则凝胶状产出(图 4-E, 4-F); 另一种为磷质泥晶细纹层, 通常与矿石内泥质成分呈互层状产出, 并含粉砂屑石英颗粒(图 4-G)。值得注意的是, 瓮安B矿层底部与丹寨地区磷矿层中部分泥晶磷块岩可见球状生物球粒保存(图 4-F, 4-G), 直径200~800 μ m, 球粒成分为磷酸盐, 球粒边缘可见明显的包壳结构, 其形态特征与瓮安生物群中疑似胚胎化石相似, 但未见球粒内的分裂发育。扫描电镜与能谱分析结果表明泥晶磷块岩中主要矿物成分为磷灰石与碎屑石英(图 4-H, 4-J, 4-K), 组成泥晶磷块岩的凝胶状或纹层状磷灰石晶体为超微不规则片状磷灰石集合体(图 4-I)。磷质页岩在沉积构造、矿物组成和矿石结构上与泥晶磷块岩相同, 但P2O5含量较低(10%~18%), 在宏观剖面上磷质页岩往往与泥晶磷块岩共生, 肉眼难以区分。
 | 图 4 贵州省遵义、瓮安和丹寨地区陡山沱组泥晶磷块岩矿石结构特征 A— 陡山沱组顶部含磷层序, 泥晶磷块岩呈中薄层状与页岩、磷质页岩互层产出, 遵义松林剖面; B— 陡山沱组顶部灰黑色中薄层磷块岩矿层, 丹寨番仰剖面; C— 深灰色中薄层泥晶磷块岩层, 遵义松林剖面; D— 黑色页岩中的水平层理, 遵义松林剖面; E— 泥晶磷块岩偏光显微镜下结构照片, 单偏光, 磷质呈凝胶状赋存于石英、泥质碎屑基质中, 遵义松林剖面; F— 泥晶磷块岩, 单偏光, 磷质呈凝胶状产出于基质中, 可见磷酸盐化球状生物化石, 瓮安ZK511钻孔; G— 泥晶磷块岩, 单偏光, 磷质以纹层状产出, 可见磷酸盐化球状生物化石, 丹寨番仰剖面; H— 泥晶磷块岩扫描电镜照片, 背散射成像, 遵义松林剖面; I— 泥晶磷块岩中磷灰石晶体显微结构, 二次电子成像, 遵义松林剖面; J、K— 图H打点位置相关矿物能谱曲线Fig.4 Structural characteristics of micritic phosphorite of the Doushantuo Formation in Zunyi, Weng'an and Danzhai areas, Guizhou Province |
3.1.3 含生物化石磷块岩
此类磷块岩类型中含有丰富、多类型的生物化石, 其中ZK032钻孔样品中主要包含多细胞藻类化石和疑似胚胎动物化石。藻类化石中可以观察到多细胞结构, 由成百上千的小分裂细胞聚集而成, 体积较大, 整体形态呈网格状分布(图 5-A); 此外化石体内常见被碳酸盐岩矿物充填的微型管道结构(图 5-B), 被认为是由黄铁矿或其他晶体颗粒在基质中受有机质降解释放的气体压力推动而留下的作用痕迹, 被称为异质体拖拽轨迹(AITs)(She et al., 2016; 杨晓光和韩健, 2017)。疑似胚胎动物化石呈圆球粒状, 化石直径0.3~1.0 mm(图 5-C, 5-D), 球粒化石含磷大于70%, 可见球粒外包壳、表膜和囊内部结构, 壳体容易出现内陷和褶皱现象, 使壳体表明呈现“ 脑纹状” 和“ 瘤状” 等多种规则的装饰(图 5-E); 疑似胚胎动物球体化石内有分裂生长(图 5-E), 颗粒内部可见微粒磷质颗粒(图 5-F), 推测为胚胎分裂后的微粒化石; 球体化石之间胶结物一般为亮晶白云石(图 5-D)。化石除保存了有机体组织和细胞的外观结构外, 其内部充填物构造复杂, 常见空隙和各种不同的生物学结构, 比如疑似胚胎化石中卵裂球、卵裂构造和围卵腔等(图 5-G)。
 | 图 5 瓮安陡山沱组含生物化石磷块岩矿石结构特征 A、B— 瓮安陡山沱组B矿层磷块岩内多细胞藻类生物化石, 分别为单偏光和正交偏光, ZK032; C、D— 瓮安陡山沱组B矿层磷块岩内疑似胚胎生物球粒化石结构和形态特征, 单偏光, ZK032; E— 疑似胚胎生物球粒化石, 可见内部分裂结构, 单偏光, ZK511; F— 疑似胚胎生物球粒化石, 可见内部微粒磷质颗粒, 单偏光, ZK032; G— 疑似胚胎生物球粒化石, 可见球粒外缘管腔构造, 单偏光, ZK511Fig.5 Structural characteristics of phosphrite with biological fossils of the Doushantuo Formation in Weng'an area |
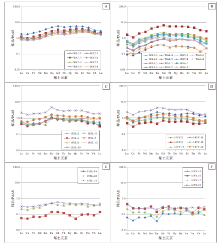
瓮安ZK032、遵义松林剖面和丹寨番仰剖面陡山沱组磷块岩稀土元素分析结果见表 1和表2。瓮安陡山沱组A矿层球粒磷块岩稀土元素总量(Σ REE)为168.6× 10-6~428.7× 10-6, 平均值225.85× 10-6; B矿层磷块岩样品稀土元素总量明显低于A矿层, Σ REE值为23.9× 10-6~368.1× 10-6, 平均值109.0× 10-6。松林和番仰陡山沱组剖面Ⅳ 段泥晶磷块岩和磷质页岩样品Σ REE值相似, 分别为76.8× 10-6~356.5× 10-6(平均值132.26× 10-6)和77.0× 10-6~406.4× 10-6(平均值185.33× 10-6), 均高于两地区Ⅲ 段和Ⅳ 段泥晶白云岩样品中稀土元素含量(松林和番仰泥晶白云岩Σ REE值分别为4.5× 10-6~60.9× 10-6和11.0× 10-6~52.7× 10-6)。瓮安A矿层球粒磷块岩表现出无Ce负异常(Ce/Ce* =0.98~1.13, 平均值1.07)和Eu无异常到弱正异常(Eu/Eu* =1.07~1.21, 平均值1.15); B矿层WA3-3泥晶磷块岩样品为弱负Ce异常(Ce/Ce* =0.93)和Eu正异常(Eu/Eu* =1.39), 其余样品则表现出明显的Ce负异常(Ce/Ce* =0.53~0.80, 平均值0.68)和弱Eu正异常(Eu/Eu* =1.11~1.24, 平均值1.19)。松林剖面陡山沱组Ⅳ 段磷块岩和磷质页岩样品为弱Ce负异常到无异常(Ce/Ce* =0.76~1.19, 平均值0.95)和较明显的正Eu异常(1.33~1.93, 平均值1.60), Ⅲ 段泥晶白云岩样品则表现出不稳定的Ce异常(Ce/Ce* =0.72~1.14)和Eu正异常(1.08~1.52)。番仰剖面陡山沱组Ⅳ 段泥晶磷块岩(包括磷质页岩)和泥晶白云岩均存在Ce负异常, Ce/Ce* 值分别为0.68~0.91(平均0.77)和0.62~0.96(平均0.85), 磷块岩Eu异常值除14FY-3样品为负异常外(Eu/Eu* =0.79)外均为弱正异常(Eu/Eu* =1.08~1.38, 平均值1.25), 而泥晶白云岩值表现为明显Eu负异常(Eu/Eu* =0.43~0.81, 平均值0.56)。PAAS(澳大利亚后太古代页岩)标准化稀土元素配分曲线显示瓮安、松林和番仰陡山沱组磷块岩和磷质页岩样品均有中稀土微弱富集的“ 帽型” 分布特征(图 6-A, 6-B); 松林和番仰陡山沱组泥晶白云岩稀土元素配分曲线则表现出较为平坦分布的趋势特征(图 6-C, 6-D, 6-E, 6-F)。
 | 图 6 贵州瓮安、遵义和丹寨地区陡山沱组磷块岩、磷质页岩和泥晶白云岩样品稀土元素PAAS标准化稀土元素配分曲线 A— 瓮安ZK032剖面A矿层磷块岩; B— 瓮安ZK032剖面B矿层磷块岩; C— 遵义松林剖面磷块岩和磷质页岩; D— 丹寨番仰磷块岩和磷质页岩; E— 遵义松林剖面泥晶白云岩; F— 丹寨番仰剖面泥晶白云岩Fig.6 PAAS-normalized REE patterns of phosphorites, phosphatic shales and dolomicrites in Zunyi, Weng'an and Danzhai areas, Guizhou Province |
磷块岩中的稀土元素通常是在沉积成磷过程中以类质同象的形式取代磷灰石的Ca2+进入矿物晶格中, 因此磷块岩中的稀土元素地球化学特征往往可以反映其沉积水体氧化还原状态(Jarvis et al., 1994; Joosu et al., 2015)。然而磷块岩沉积过程中陆源碎屑的输入、胶结物胶结与成岩改造作用往往会对磷块岩中稀土元素配分模式造成不同程度的影响(Chen et al., 2003; 肖朝益等, 2018)。通过岩相观察, 瓮安地区磷块岩中陆源碎屑矿物含量极低, 而遵义松林和丹寨番仰地区磷块岩样品中含有一定量的陆源泥质或粉砂质硅酸盐矿物成分, 陆源碎屑矿物内所包含的母岩稀土元素在沉积和成岩阶段均会对原生沉积磷块岩中稀土元素地球化学特征造成干扰(Reynard et al., 1999; 解启来等, 2003)。
通过对样品中代表陆源碎屑含量的Al元素和Σ REE值之间的相关性判断陆源输入对原生磷块岩稀土元素值造成的影响, 相关性越高则表明干扰越大。瓮安、丹寨和番仰陡山沱组样品中Al2O3含量与Σ REE值均无明显相关性(图 7-A), 说明其稀土元素值受陆源碎屑影响较小。瓮安地区磷矿层磷质颗粒或生物球粒均为白云石胶结, 其胶结作用通常发生在沉积物水界面附近, 与磷灰石成磷环境相似, 前人对瓮安磷块岩磷质球粒和白云石胶结物中稀土元素配分特征研究, 表明2种配分曲线存在一致性(Chen et al., 2003; 解启来等, 2013)。“ 帽型” 稀土元素配分模式是大多数海相沉积型磷块岩的普遍配分模式, 这可能是由于成磷过程中有机质或铁锰氧化物优先吸附中稀土元素(MREEs)并释放在成磷水体中导致的(Kidder and Eddy-Dilek, 1994; Haley et al., 2004)。然而在成岩作用过程中, 中稀土元素(MREEs)由于与Ca2+有相似的离子半径而优先进入磷灰石晶格, 因此成岩改造强烈的磷块岩往往具有中稀土极度富集特征, 稀土元素配分曲线呈现明显的“ 上凸型” 配分模式(Reynard et al., 1999; Morad and Felitsyn, 2001; Joosu et al., 2015, 2016)。研究区磷块岩样品稀土元素配分曲线多为中稀土微弱富集的“ 帽型” 配分模式, 而非中稀土明显富集的“ 上凸型” , 且磷灰石晶体多为原生超微几何体形态, 粒柱状重结晶磷灰石晶体较为少见, 指示磷块岩样品原生稀土元素信息得以良好保存。此外Ce异常与DyN/SmN的负相关性可以检测成岩作用对磷块岩稀土元素配分的影响程度(Shields and Stille, 2001), 而研究区磷块岩和白云岩样品中Ce/Ce* 值与DyN/SmN值无明显负相关(图 7-B), 同样证明了成岩作用未对磷块岩稀土元素特征产生显著影响。因此, 通过以上岩相学与地球化学分析表明, 研究区磷块岩样品稀土元素特征基本保存了原生沉积水体地球化学信息。
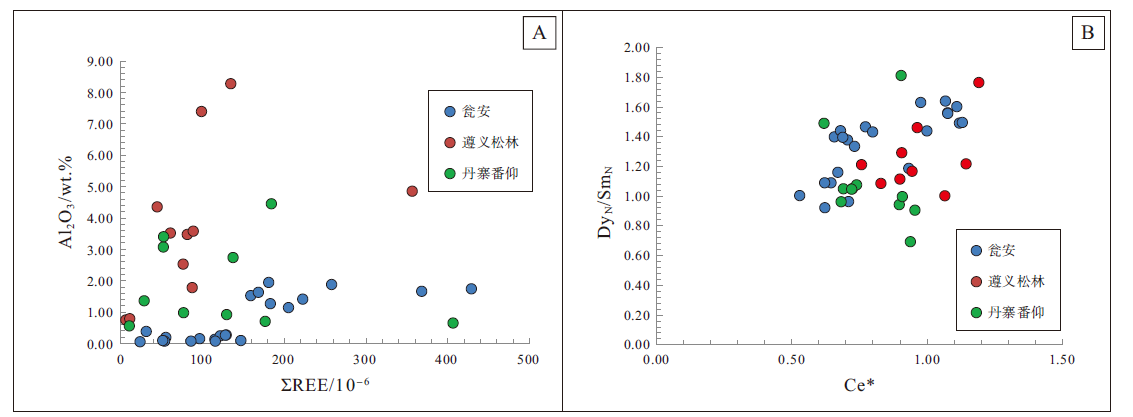 | 图 7 贵州瓮安、遵义和丹寨地区陡山沱组样品Σ REE-Al2O3(A)和Ce* -DyN/SmN(B)元素关系Fig.7 Element correlations of Σ REE-Al2O3(A)and Ce* -DyN/SmN in the Doushantuo samples in Zunyi, Weng'an and Danzhai areas, Guizhou Province |
正常海水地球化学条件下, 海洋中的磷酸盐浓度往往难以达到饱和, 因此磷灰石矿物很难在海水中以无机沉降的形式直接产出(Arning et al., 2009)。成磷作用是磷酸盐在氧化还原作用交替的沉积物— 水界面附近, 经过有机质降解释放、生物化学作用聚集和微生物吸附使孔隙水中磷浓度达到饱和并形成磷灰石自生沉降的过程(Fö llmi, 1996; Arning et al., 2009; Pufahl and Groat, 2017)。不同成磷作用模式下形成的磷块岩往往存在不同的结构特征, 磷灰石晶体矿物形态、矿物组合类型和成矿环境是解析成磷作用模式的重要依据。
瓮安地区A矿层磷块岩以球粒状磷块岩为主, 矿石内普遍存在的次生含铁矿物黄铁矿和海绿石则表明“ Fe-氧化还原泵” 模式是诱导沉积水体中磷质富集的关键机制(Algabri et al., 2020)。Fe-氧化还原泵模式是发生在氧化还原界面附近的一种富磷循环机制, 氧化水体环境下铁氢氧化物(FeOOH)高效吸附海洋中的磷酸盐后沉降海底, 在次氧化到还原环境中磷酸盐解吸附并被释放到孔隙水中形成聚集, 而氧化还原界面附近的铁氢氧化物通过不断的吸附、解吸附作用有效阻止了磷酸盐的逸出, 从而使磷酸盐浓度不断提升并形成沉降(Compton et al., 2000; Neslon et al., 2010)。在这一成磷作用系统中, 震旦纪之前深部底层海水的铁化缺氧环境提供了大量的铁质来源(Canfield et al., 2008; Li et al., 2010; Lyons et al., 2014)。进入氧化水体的溶解铁吸附磷质并以铁磷复合物(FeOOH· PO4)形式沉降进入沉积物, 在氧化还原界面附近部分还原作用形成富含Fe2+和Fe3+的自生海绿石矿物, 在硫酸盐还原带Fe3+被完全还原为Fe2+并形成莓状黄铁矿, 铁氢氧化物(FeOOH)的还原过程同时释放了大量磷酸盐, 为磷灰石的自生沉积提供了先决条件。球粒状磷块岩内稀土元素PAAS标准化配分曲线以及表现出的无Ce负异常同样指示了成磷作用处于氧含量较低的沉积环境, 氧化还原界面下的贫氧水体是磷灰石沉积的主要场所。Fe-氧化还原泵模式成磷环境一般在沉积物— 水界面以下的孔隙水中才能达到磷酸盐饱和浓度, 但是在震旦纪古海洋极端富磷的海水条件下, 其成磷作用可能在滨岸浅海带海底氧化还原界面附近的水柱中进行(She et al., 2013; Algabri et al., 2020), 然而这一模式在深部缺氧海洋环境中难以使磷酸盐在开阔海水中直接聚集沉降(Nelson et al., 2010)。磷酸盐成矿方面, 磷质球粒内的花环状磷灰石晶体集合体与前人在硅铁建造(BIFs)中的花环状磷灰石相似, 被认为是纳米级短柱状磷灰石晶体和无定型磷灰石胶状集合体成岩演化而来, 是微生物作用聚集磷酸盐成矿的产物(Sun et al., 2014)。磷质颗粒内的核形石环带结构同样指示微生物如蓝细菌及其胞外聚合物(EPS)粘附作用下增生加积形成, 球粒内普遍分散的有机质可能为蓝细菌及EPS残余(She et al., 2013, 2014)。磷质球粒间的白云石胶结物和磷质纹层间的白云质条带, 为黄铁矿和磷质球粒沉积后, 水体环境中具有抑制白云石沉积的硫酸盐耗尽而引发的沉积产物(Pufahl and Groat, 2017), 其诱因可能为蒸发环境中石膏等蒸发矿物的沉积造成沉积水体中Mg/Ca值的提升和硫酸盐的祛除(Vernhet and Reijmer, 2010; Zhang et al., 2019), 也可能是海水含磷浓度和沉积环境的交替变化造成的沉积分异(柴华等, 2014)。
贵州陡山沱组泥晶磷块岩和磷质页岩中常见原生磷质细纹层, 通常与陆源细碎屑泥质或粉砂质沉积物共生, 是低能静海沉积环境下形成的自生富磷沉积物(Garrison and Kastner, 1990; Soudry, 1992; Arning et al., 2009)。与球粒状磷块岩不同, 泥晶磷块岩和磷质页岩中次生矿物成分以陆源碎屑沉积的石英和黏土矿物为主, 几乎不含指示“ Fe-氧化还原泵” 模式成磷作用的标志次生矿物如海绿石, 然而矿层与富含有机质的碳质页岩共生, 且有生物化石保存, 指示成磷作用可能与沉降的有机质降解释放磷酸盐有关。有机质沉降模式是在松散的泥砂质沉积物— 水界面以下通过一系列微生物参与的氧化还原反应进行的, 有机质中降解使其包含的磷迅速释放在孔隙水中(如硫酸盐还原细菌降解作用), 并使孔隙水中的磷酸盐浓度急剧提升达到饱和沉降(Schulz and Schulz, 2005; Arning et al., 2009; Goldhammer et al., 2010; Bailey et al., 2013)。泥晶磷块岩中不规则片状磷灰石集合体的组成结构, 指示了有机质释放磷质进入孔隙水中、并使磷酸盐浓度过饱和迅速沉降并受上覆沉积物压实的沉积过程(Joosu et al., 2016)。遵义松林、丹寨番仰地区陡山沱组Ⅳ 段泥晶磷块岩, 和与其共生的磷质页岩层表现出的Ce负异常, 均显示成磷作用发生在氧化还原界面附近次氧化到氧化的沉积环境。
含生物化石磷块岩主要分布于瓮安地区B矿层, 生物化石在丹寨地区泥晶磷块岩中也有少量保存。这些生物化石为有机体(动物休眠卵和胚胎、藻类以及疑源类)死亡后早期成岩阶段在富磷水体中迅速磷酸盐化的产物(Xiao and Knoll, 1999; 刘鹏举等, 2009; 袁训来, 2009)。这种由生物遗体磷酸盐化形成磷块岩的生物成矿模式同样是促使成磷作用运行的主要机制, 但由于生物作用规模的限制一般出现在生物异常繁盛且水体富磷的浅水海岸沉积区(杨瑞东, 1999)。瓮安B矿层含生物化石磷块岩中低Ce负异常值, 指示了氧化状态的富磷水体环境, 而正是海洋中丰富的磷酸盐和较为氧化的水体条件, 为高等生命的演化爆发提供了有利场所。
新元古代末期全球性成磷事件, 被认为是地质历史时期全球范围内首次出现的大规模富磷沉积物沉积事件(Cook and Shergold, 1984; Cook, 1992)。本次成磷事件在时空分布上与新元古代一些列重大地质变革、特别是新元古代末期氧化事件(NOE)存在密切的耦合关系: “ 雪球地球” 后气候转暖, 冰川迅速融化, 大量陆源风化磷酸盐等营养成分进入海洋系统, 伴随大气及海洋氧含量不断增加, 造成大洋生产力繁盛与生命演化爆发, 同时在氧化的海洋中引发了广泛的富磷沉积物发育(Brasier and Callow, 2007; Papineau, 2010; Pufahl and Hiatt, 2012; She et al., 2013, 2014; Boyle et al., 2014; Drummond et al., 2015; Pufahl and Groat, 2017)。与古元古代全球首次成磷事件(约2.2— 1.8 Ga)中磷块岩沉积被限制在极浅的、光合作用影响的含氧海洋绿洲中相比, 本次成磷事件伴随深部海洋的不断充氧, 成磷沉积作用规模更大、范围更广泛(Nelson et al., 2010; Pufahl and Hiatt, 2012)。
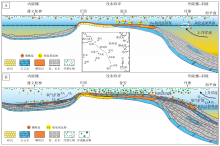
华南地区震旦系纪磷块岩广泛发育在扬子地台上, 尤其是分布于贵州省的开阳、瓮安磷矿、遵义磷矿和丹寨磷矿, 代表了浅水海岸至深水陆棚相磷块岩沉积环境, 是目前发现的世界上唯一在新元古代末期在整个陆架台地上磷矿层连续完整发育的地区。陡山沱早期仅在黔中古陆周缘浅水海岸存在磷矿层沉积, 瓮安地区A矿层磷块岩成磷作用模式为Fe-氧化还原泵系统下的微生物聚集粘结成磷, 无Ce负异常指示沉积水体处于较低的含氧水平; 遵义松林地区直到陡山沱组Ⅲ 段泥晶白云岩层开始出现不稳定的Ce负异常, 但无磷矿层发育。这表明成磷事件早期海洋含氧量普遍较低, 仅在浅部海水沉积区出现氧化带, 并在氧化还原界面附近通过铁氢氧化物的富磷循环和微生物聚集作用形成磷灰石沉降(图 8-A); 而陆棚— 斜坡沉积相区深部海水在陡山沱早中期仍为缺氧状态, 氧化还原界面不足以到达深部海水底层, Fe氢氧化物和有机质吸附的磷酸盐, 在氧化还原界面以下分解释放重新进入海水, 在广阔的海水中磷酸盐浓度难以达到饱和, 并不能形成磷灰石的聚集沉降, 因此进入深海的磷酸盐会伴随上升洋流重新回到表层海水循环利用(图 8-A)。
 | 图 8 贵州瓮安、遵义和丹寨地区成磷事件早期(A)和成磷事件晚期(B)陡山沱组磷块岩成磷作用模式Fig.8 Phosphogenesis models during early(A)and late(B)phosphogenic event from Doushantuo phosphorites in Zunyi, Weng'an and Danzhai areas, Guizhou Province |
陡山沱晚期开始出现普遍成磷现象, 瓮安地区B矿层发育泥晶磷块岩与含生物化石磷块岩, 成磷作用也由原先的“ Fe-氧化还原泵” 模式向“ 有机质沉降” 模式和“ 生物成矿” 模式转变, 成磷环境也逐渐由贫氧向次氧化、氧化状态演化, 指示了陡山沱组沉积从早期到晚期海洋氧含量的增加; 同时遵义松林、丹寨番仰陡山沱组Ⅳ 段发育了泥晶磷块岩沉积, 且Ce负异常同样指示相对氧化的水体环境, 表明深部海水不断充氧, 氧化还原界面扩散水体较深的陆棚— 斜坡沉积相区(图 8-B), 这与新元古代氧化事件过程中深海通氧过程形成耦合。陡山沱组矿层磷块岩中生物化石丰度、数量明显提升, 并逐渐出现多细胞高等生物, 推测是震旦纪古海洋沉积环境中氧气含量不断增加和丰富的磷酸盐供给引起的, 而多细胞生物的不断演化发展同时也永久改变了海洋含氧结构 (Lenton et al., 2014)。陡山沱早中期深部海水磷酸盐的循环利用过程, 进一步增强了大洋表层生产力, 使表层浮游植物不断光合作用产生氧气, 而海洋中充足的营养物质来源和不断升高的氧含量, 为生物的演化提供了物质基础和适宜环境, 形成了以瓮安动物群为代表的高等多细胞生物演化, 且多细胞动物的快速埋藏减少了对海水氧气因对有机质降解而造成的消耗, 从而促使氧气向深海扩散。伴随深部海水进一步氧化, 直到Gaskiers冰期后大洋底部逐渐全部被氧化(Hiatt et al., 2015), 成磷作用逐渐由向古陆周缘浅水海岸深部海水延伸, 最终形成大规模磷矿床沉积。
因此, 陡山沱组磷块岩成磷作用和沉积分布, 很好地记录了新元古代氧化事件海洋系统氧化还原条件转变进程, 海洋氧气含量提升不仅影响了磷块岩成磷模式和沉积水体环境, 并且扩展了磷块岩沉积范围, 而磷酸盐的聚集和海洋增氧为生物的演化和爆发提供了良好的生存条件, 生态系统的发展同时也造成了海洋氧化状态的转变, 形成了氧化事件、成磷作用和生态系统演化的相互制约和密切耦合关系。
1)瓮安地区A矿层球粒磷块岩成磷作用是通过Fe-氧化还原泵模式和微生物聚磷作用形成的磷灰石自生沉积, B矿层和遵义、丹寨地区泥晶磷块岩则通过有机质降解释放磷酸盐进入孔隙水中富集成矿, 磷酸盐化生物遗体形成的含生物化石磷块岩则是生物成磷作用的产物。
2)贵州省震旦纪成磷事件早期成磷作用仅分布于浅水海岸, 成磷作用晚期则扩散至深水陆棚— 斜坡, 稀土元素地球化学特征显示磷块岩海洋水体环境逐渐由贫氧向氧化状态演化。
3)陡山沱组磷块岩成磷作用转换和沉积分布, 记录了新元古代末期氧化事件下的海洋增氧过程, 伴随海水氧含量不断提升, 磷块岩分布范围逐渐向深部海水沉积区扩展, 成磷环境逐渐富氧, 同时海洋中磷酸盐和氧含量的提升造成的生态系统演化也促进了深海通氧。
(责任编辑 郑秀娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|