



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
西秦岭甘南地区上二叠统毛毛隆组古环境演变:基于遗迹网络分析*
[党志英 , 张立军
, 张立军
, 张立军]
|
|

第一作者简介:党志英,女,1994年生,硕士研究生,主要从事沉积学和遗迹化石研究。E-mail: zydanghpu@126.com。
西秦岭地区是二叠系—三叠系之交深水相保存较为连续的地区,西秦岭甘南地区上二叠统毛毛隆组发育丰富的遗迹化石(12属17种)。遗迹网络分析作为新兴的研究方法,侧重于用相互连接的遗迹分类群来重建遗迹群落系统,这为研究遗迹群落的行为学和生态学提供了丰富的手段。笔者以西秦岭甘南地区上二叠统毛毛隆组发现的遗迹化石为数据源,运用遗迹网络分析方法,揭示出毛毛隆组遗迹群落具结构化特征,遗迹群落体现出环境驱动的遗迹组合面貌特征: Palaeophycus-Lockeia-Arenicolites-Dictyodora-Planolites-Protovirgularia-Chondrites组合(C1,近源浊流区),Palaeophycus-Lockeia-Arenicolites-Dictyodora-Protovirgularia-Phycodes-Chondrites-Helminthopsis组合(C2,浊流过渡区—浊流远源区),和Palaeophycus-Nereites-Scolicia-Protovirgularia-Helminthopsis组合(C3,远源浊流区)。该遗迹群落由C1演变至C3的过程,揭示了古环境从近源浊流区至远源浊流区的演变历程。
通讯作者简介:张立军,男, 1982年生,副教授,主要从事遗迹沉积学的教学与研究。E-mail: Ljzhanghpu@gmail.com。
About the first author: Dang Zhi-Ying,born in 1994,is a master degree candidate. She is engaged in researches of sedimentology and trace fossils. E-mail: zydanghpu@126.com.
The continuous Permian-Triassic strata in West Qinling represent deep-marine deposits. In this paper,abundant trace fossils(seventeen ichnospecies,twelve ichnogenera)have been identified in the Upper Permian Maomaolong Formation in the Gannan area of the western Qinling orogen. As a new ichnologic method,ichnonetwork analysis focuses on the reconstruction of the community system composed of interconnected ichnotaxa,which provides useful means for studying the behavior and ecology of ichnocoenosis. In this study,an ichnonetwork analysis is conducted with trace fossils data obtained from the first hand filed trip. The results show that ichnocoenosisis structured in the Maomaolong Formaiton,and three ichnoassociations were controlled by palaeoenvironments: Palaeophycus-Lockeia-Arenicolites-Dictyodora-Planolites-Protovirgularia-Chondrites (C1,proximal turbidite zone), Palaeophycus-Lockeia-Arenicolites-Dictyodora-Protovirgularia-Phycodes-Chondrites-Helminthopsis (C2,turbidite transition zone-distal turbidite zone),and Palaeophycus-Nereites-Scolicia-Protovirgularia-Helminthopsis (C3,distal turbidite zone). The evolution from C1 to C3 in the Upper Permian Maomaolong Formation from the Gannan area of the West Qinling reveals the palaeoenvironmental changes from proximal turbidite zone to distal turbidite zone.
网络分析作为多学科交叉研究的新兴领域, 主要是由相互连接的实体组成的类似于网络的系统, 近年来在信息科学和生态学中被广泛应用(Montoya et al., 2006; Bascompte, 2007)。在网络理论(Barabási, 2002; Lewis, 2009)的基础上, 遗迹网络分析研究方法应运而生(Baucon and Felletti, 2013a, 2013b)。基于遗迹分类群聚集在复杂网状结构(Ichnonetwork)中的假设, 网络理论被应用到遗迹组合分析中。在遗迹学中, 遗迹组合是(古)环境分析的主要基础, 一个遗迹学系统可以看作是一组相互联系的遗迹分类群, 其拓扑结构取决于生物与环境的相互作用。因此, 遗迹网络分析作为一种新兴的遗迹学研究方法, 为古环境分析提供了丰富的手段。西秦岭甘南地区二叠系海相地层中遗迹化石形态多样且保存完整, 为遗迹网络分析提供了重要的素材, 文中旨在将网络分析理论应用于恢复和判别该地区上二叠统毛毛隆组的遗迹群落类型, 进而重建其古生态学和古环境特征。
研究剖面地处西秦岭北部的甘肃省甘南藏族自治州合作市一带(图 1-a)。该地区南接松潘— 甘孜地块, 北接祁连山造山带, 西接柴达木古陆, 二叠纪时位于东古特提斯洋北部和华北板块南部(Shi, 2006)(图 1-b)。西秦岭北部地层主要为一套巨厚层浊流沉积的细碎屑岩和碳酸盐岩建造(左国朝等, 1984; 方国庆等, 1994; 罗根明等, 2007)。在西秦岭与南祁连造山带交接处的甘肃省夏河县发现了玄武岩与碳酸盐岩共生的海山建造, 该建造被认为是具有独特的喷气孔、杏仁体和枕状结构的洪泛型玄武岩, 揭示出西秦岭二叠纪时仍为多岛小洋盆环境(寇晓虎等, 2007; Kou et al., 2009)。同时, 在甘南地区二叠系发现了大套具明显的成层性、呈似层状或透镜体的砾屑灰岩, 其在平面上呈条带状分布, 与其他围岩呈韵律性分布或岩性过渡, 具滑塌构造, 反映了该地区位于大陆边缘、发育受一系列重力流影响的台前斜坡相沉积的特征(左国朝等, 1984; 李柒林等, 2002)。
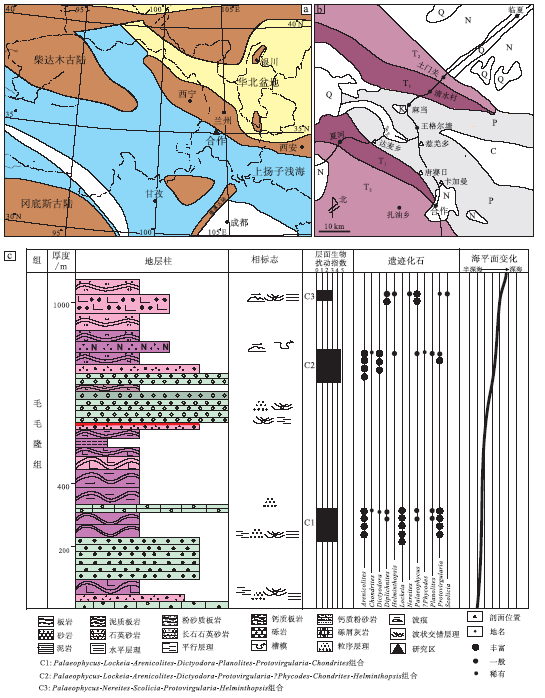 | 图 1 西秦岭甘南地区交通位置、古地理简图以及上二叠统毛毛隆组岩性柱状图(交通位置、古地理图据王鸿祯, 1985; 殷鸿福等, 1992; 有修改)Fig.1 Location, palaeogeographic map of Gannan area in West Qinling and stratigraphic column of the Maomaolong Formation of Upper Permian(location and palaeogeographic map modified from Wang, 1985; Yin et al., 1992 |
西秦岭甘南地区上二叠统毛毛隆组可以分为2个岩性段:下段以深灰色、灰色厚层到块状砾屑灰岩、砾岩及中细粒石英砂岩、薄层粉砂岩、板岩互层为主, 发育平行层理、交错层理、槽模和丰富的遗迹化石, 如Arenicolites, Chondrites, Dictyodora, Lockeia, Helminthopsis, Palaeophycus, Phycodes, Planolites, Protovirgularia; 上段以灰黑色、褐色中细粒砂岩及薄层板岩为主, 发育水平层理、波状层理等, 见较多遗迹化石, 如Nereites, Scolicia, Diplichnites, Helminthopsis, Palaeophycus, Protovirgularia(图 1-c)。甘南地区上二叠统毛毛隆组为一套具有斜坡沉积特征的深水沉积物, 是在沉积盆地不断裂陷的环境中形成的(左国朝等, 1984; 赵江天和杨逢清, 1992; 李柒林等, 2002)。
首先, 根据Baucon和Felletti(2013b)、Baucon等(2014, 2015)所描述遗迹网络分析方法, 将野外实测的遗迹化石数据集作为绘制遗迹网络的源数据。具体来说, 源数据是记录每个采样单元中每个遗迹群落是否存在的数据集, 绘制出的遗迹网络表示的是一组遗迹化石组合之间的关联关系。源数据分析的具体操作是将遗迹化石数据存入Excel电子表格, 其中遗迹分类群为列, 样品为行(图 2-a)。
 | 图 2 遗迹网络分析方法步骤(据Baucon et al., 2014, 2015)Fig.2 Drawing steps used for ichnonetwork analysis method(after Baucon et al., 2014, 2015) |
其次, 计算每个遗迹群落对的Jaccard索引。绘制遗迹网络的过程是基于遗迹分类群对之间的关联关系, 其思想是将遗迹分类群表示为节点, 并将一起出现的那些对连接起来, 每个关联关系的强度由链接权重来描述, 该链接权重对应于Jaccard索引(Jaccard, 1901, 1912; Hammer and Harper, 2006)。例如, 遗迹分类群A和遗迹分类群B的Jaccard指数是其中一个分类群或另一个分类群的相关发生数与样本数的比例(Jaccard, 1901, 1912; Hammer and Harper, 2006): J(A, B)=(A和B的样本数)/(A或B的样本数)。换句话说, Jaccard指数可以看作是一种关联的概率度量, 它表示如果样本中至少存在2种遗迹属中的1种, 则找到这2种遗迹属的概率(Jaccard, 1901, 1912; Hammer and Harper, 2006; Baucon and Felletti, 2013b)。计算Jaccard索引可以使用统计软件(Past 3.26)以矩阵形式(邻接矩阵)来保存和计算结果, 每组结果都是给定一对遗迹分类群对的Jaccard索引指数。例如, 将源数据导入统计软件(Past 3.26)中, 在软件中选择Edit→ Transpose, Statistics→ Similarity and distance indices→ Jaccard→ Copy, 再将结果进行保存(图 2-b)。
最后, 将邻接矩阵呈现为网络。遗迹网络图中的节点指代遗迹分类群, 其地理范围对应于一组特定的环境条件, 链接则指代遗迹之间的关联关系, 链接的节点为关联的遗迹分类群。笔者将上述保存的矩阵数据导入网络分析软件(Gephi 9.02)生成网络图形(图 2-c)。
Arenicolites isp. 为垂直于层面的U形潜穴, 潜穴直径1~8imm, 间距0.8~12imm, 管壁光滑, 局部可见衬里(图 3-a)。Arenicolites的产出环境广泛(Pemberton et al., 2001; Feng et al., 2019), 该遗迹属被认为是蠕虫类环节动物和小型甲壳动物的居住构造(杨式溥等, 2004)。
Chondrites intricatus呈树形分枝、扁平、上浮雕保存的潜穴, 直径为0.5~5imm, 分枝角度为25° ~40° (图 3-b)。Chondrites主要以内生迹形式保存, 属于广相遗迹化石, 通常指示缺氧/贫氧环境(Ekdale and Mason, 1988)。Chondrites isp. 短羽状分枝, 分枝较短且不密集, 具25° ~35° 的分枝角, 直径约0.2imm(图 3-c)。
 | 图 3 西秦岭甘南地区上二叠统毛毛隆组遗迹化石 a— Arenicolites isp.(Ar), Dictyodora cf. scotica(Di), 上浮雕保存; b— Chondrites intricatus, 上浮雕保存; c— Chondrites isp., 上浮雕保存; d— Dictyodora zimmermanni, 下浮雕保存; e— Diplichnites isp., 全浮雕保存; f— Diplichnites triassicus, 上浮雕保存; g— Lockeia siliquaria(Lo), Protovirgularia isp.(Pr), 下浮雕保存Fig.3 Trace fossils from the Upper Permian Maomaolong Formation in Gannan area of West Qinling |
Dictyodora cf. scotica为蜿蜒曲折的蛇曲形潜穴, 蛇曲幅度较高, 互相切穿, 二级弯曲, 直径3~10imm(图 3-a)。Dictyodora是一种呈网状三维结构的复杂遗迹, 属于Nereites遗迹相(Wetzel and Uchman, 2012), 其造迹生物可能是一种蠕虫或没有壳的软体动物(Benton and Trewin, 1980)。Dictyodora zimmermanni为不规则的蛇曲形潜穴, 个别单个的弯曲被拉长, 具有尖锐的转弯和锐角, 潜穴直径为0.7imm(图 3-d)。
Diplichnites triassicus沿层面上保留有多条直的印痕, 每条印痕由2排平行的、等距的、不等细的凹坑组成, 2排印痕的间距2~10imm。在所有的印痕中, 起点印痕最深, 沿着轴线逐渐变浅或消失(图 3-f)。Diplichnites isp. 由平行排列的一系列小凹坑组成, 运动路径垂直或倾斜状分布, 潜穴宽10~20imm, 小凹坑大小约1.5imm(图 3-e)。
Helminthopsis tenuis为简单弯曲的蛇形拖迹, 呈宽缓的不规则U形转弯, 直径约2imm(图 4-c)。该遗迹属的造迹者可能是多毛类动物(Fillion et al., 1990), 广泛分布在海相地层中(杨式溥等, 2004)。Helminthopsis isp. 呈简单的、不规则蛇曲形, 潜穴表面光滑无纹饰, 潜穴直径约3imm(图 4-b)。
 | 图 4 西秦岭甘南地区上二叠统毛毛隆组遗迹化石 a— Scolicia strozzii(Sc), Nereites missouriensis(Ne), 下浮雕保存; b— Helminthopsis isp., 下浮雕形式保存; c— Helminthopsis tenuis, 下浮雕保存; d— Palaeophycus isp., 下浮雕保存; e— Planolites isp., 上浮雕保存; f— Palaeophycus tubularis, 下浮雕保存; g— Phycodes isp., 下浮雕保存Fig.4 Trace fossils from the Upper Permian Maomaolong Formation in Gannan area of West Qinling |
Lockeia siliquaria呈杏仁形、果核形的铸型凸起, 表面光滑无饰, 长2~4imm, 宽0.5~1.0imm(图 3-g)。该遗迹属的造迹生物是双壳类(Uchman, 2004), Lockeia siliquaria被认为是双壳类动物在伸展阶段的足部铸型或贝壳的压痕铸型(Osgood, 1970; Seilacher, 1994)。
Nereites missouriensis不规则弯曲的链状潜穴, 由一系列椭圆形或卵形瘤状体组成, 且两侧首尾闭合, 2排足迹间距5~10imm(图 4-a)。该标本中心主动回填构造不明显, 可能是风化强烈而呈凹下的状态, 两侧的突起在遗迹的首尾都是闭合的(图 4-a), 推测应该是潜穴的一部分。Nereites missouriensis为广相型遗迹, 常见于显生宙的浊积岩(Crimes, 1987; Wetzel, 2002)。
Palaeophycus tubularis为表面光滑、不分枝、直到略弯曲的柱状潜穴, 相互交叉, 衬里较薄, 充填物与主岩相同, 直径2~6imm(图 4-f)。该遗迹属被认为是捕食性蠕虫或者滤食性动物的居住潜穴(Pemberton and Frey, 1982), 属于广相遗迹(Fillion et al., 1990)。Palaeophycus isp. 为水平的、不分枝的柱形潜穴, 衬里薄, 与主岩相同的洞穴充填, 直径5~12imm(图 4-d)。
Phycodes isp. 水平束装的柱状潜穴, 整体宽度12icm, 微微弯曲, 相互靠近或微微分开(图 4-g)。该遗迹属的造迹生物被认为是蠕虫状的环节动物、海鳃动物和海笔(Hä ntzschel, 1975; Bradley, 1980, 1981), 主要分布在浅海环境中(Seilacher, 1964), 但在微咸水环境和深水环境也有记录(Narbbonne, 1984)。
Planolites isp. 为直到微弯的柱形潜穴, 潜穴表面光滑无饰, 无衬里, 无分枝, 潜穴直径0.3~2imm(图 4-e)。Protovirgularia isp. 为直到略弯曲的柱状潜穴, 不分枝, 潜穴表面具有V字形纹饰, V形指向彼此相对, 且V形脊线分布较聚集, 潜穴宽1~4imm(图 3-g)。Protovirgularia被解释为双壳动物活动过程留下的痕迹, 广泛分布于陆相及海相地层中(Mángano et al., 1998; Uchman, 1998; Carmona et al., 2010)。
Scolicia strozzii为略弯曲的双叶脊, 具中沟, 横切面呈宽U字形, 山脊宽60imm(图 4-a)。该遗迹属被认为是海胆纲动物的爬行迹(Smith and Crimes, 1983), 在深海复理石相和浅海Cruziana遗迹相中均有出现(杨式溥等, 2004)。
遗迹网络最直接的特征是由不同遗迹分类群(节点)连接组成。节点数反映的是特定地层中存在的遗迹化石数量, 与遗迹多样性相对应(Buatois and Mángano, 2011), 而遗迹多样性通常被认为是环境压力的反映(Gingras et al., 2011)。同时, 节点的大小被称为节点度, 指代与节点关联的链接数(Wassermann and Faust, 1994), 而孤立的节点位置是没有直接拓扑意义的(Wassermann and Faust, 1994), 很可能代表受胁迫的生态系统(Bromley, 1996; Gingras et al., 2011)。例如, 在所研究的遗迹网络中, 孤立的节点是遗迹群落C3中发育的Diplichnites, 与网络的其余部分完全断开连接(图 2-c)。此外, 遗迹网络图形显示了遗迹群落之间的关系, 而且还记录了每个关系的强度, 连接是遗迹之间的关联关系, 每个关联关系的强度由链接权重来描述。遗迹分类群的环境范围之间的重叠程度越高, 相应的链接权重就越大(Baucon et al., 2014, 2015)。
在毛毛隆组剖面发现的遗迹化石, 依据遗迹化石的组分、分布特点、丰富程度以及网络拓扑分析, 可划分出3个遗迹组合:在毛毛隆组下段的灰黑色中厚层细粒石英砂岩和板岩层面上可观察到Palaeophycus-Lockeia-Arenicolites-Dictyodora-Planolites-Protovirgularia-Chondrites组合(C1)和Palaeophycus-Lockeia-Arenicolites-Dictyodora-Protovirgularia-Phycodes-Chondrites-Helminthopsis组合(C2)2个组合; 在毛毛隆组上段的深灰色砂岩底层面上可观察到Palaeophycus-Nereites-Scolicia-Protovirgularia-Helminthopsis组合(C3)。
3.2.1 C1组合:近源浊流区
 | 图 5 西秦岭甘南地区上二叠统毛毛隆组遗迹网络/遗迹组合图Fig.5 Trace fossils associations and ichnonetwork from the Upper Permian Maomaolong Formation in Gannan area of West Qinling |
遗迹群落组合C1与遗迹群落组合C2有着绝大部分的重叠, 与C3有着小部分的重叠(图 5-a), 这表明遗迹多样性和层面生物扰动指数(BPBI 3-4)强度较高。尤其是Arenicolites和Lockeia显示出高的生物扰动强度, 说明它们具有群居行为。Palaeophycus, Lockeia和Arenicolites连接的节点数最高, 为节点度最高的遗迹群落, 表明其与许多其他的遗迹群落相关联, 产出环境广泛, 属于穿相遗迹属。该遗迹组合主要反映了沉积物进食者和悬浮型进食者的觅食迹和居住迹。典型的深水遗迹化石仅有Dictyodora, 稀疏的Chondrites可能代表了暂时的缺氧(Ekdale and Bromley, 1984)。砂岩层面可见发育的水平层理、交错层理和槽模, 说明形成环境为含氧量较低、营养丰富的大陆斜坡环境, 属近源浊流区。
3.2.2 C2组合:浊流过渡区— — 浊流远源区
遗迹群落组合C2与遗迹群落C1和C3存在着大部分的重叠, 其中Dictyodora和Helminthopsis之间的链接权重非常大, 表明它们经常同时出现(图 5-a)。该遗迹群落的遗迹多样性和层面生物扰动指数(BPBI 4-5)均匀但强烈。例如, 在野外发现的灰黑色粉砂质板岩表层面上的Dictyodora分异度极高, 且相互交切, 形态分异度和规则度也很高, 潜穴管直径变化显著, 尤其是Dictyodora cf. scotica底部潜穴直径最大可达10imm。同时, 发现部分的槽模结构, 槽模表面还发育着Protovirgularia和Palaeophycus。该遗迹群落中沉积物进食者和悬浮型进食者的觅食迹广泛分布, 沉积进食策略的多样性表明在沉降较细的有机颗粒时所需的水动力较低。但是雕画迹不发育, 说明沉积底质有机质含量较高(Wetzel and Uchman, 1997), 因此该遗迹群落指示水动能较弱、底质有机质含量较高的大陆斜坡环境, 位于浊流过渡区— 浊流远源区一带。
3.2.3 C3组合:远源浊流区
遗迹群落组合C3仅与遗迹群落组合C2存在小部分的重叠, 其中Nereites和Scolicia之间的链接权重非常大, 表明这些遗迹分类群经常同时出现(图 5-a)。该遗迹群落中遗迹多样性和层面生物扰动指数(BPBI 3)大幅度降低, 这可能意味着环境压力因素(如底质稳定性、水能量、含盐度和含氧量)作用发生了显著的变化。该遗迹群落中以沉积物进食者Helminthopsis, Nereites和Scolicia的牧食迹为主, 说明沉积底质有机质含量较低(Wetzel and Uchman, 1997)。Helminthopsis, Nereites和Scolicia与遗迹群落中的其他节点没有联系, 表现出与其他遗迹群落环境的不相容性, 说明每个遗迹群落都有着其特定的环境偏好。该遗迹群落中的N. missouriensis一般代表着安静的深海环境, 可以指示营养流的波动(Wetzel and Uchman, 2012)。在薄层石英砂岩和泥岩互层, 发育水平层理和波状层理, 指示重力流沉积进入后期阶段, 水流作用减弱, 为安静的、水能量较低的、底质有机质含量较低的大陆斜坡环境, 属远源浊流区。
3.2.4 讨论
从上述分析可以看出, 这种新颖的方法是描述和分析遗迹学的有力工具, 能够辨别至关重要的遗迹群落、遗迹组合及其环境意义。遗迹学研究者应该把遗迹网络作为新的分析工具和研究手段, 并与沉积学相结合, 进一步增强遗迹学在古生物学中的重要作用。但是, 这种方法仍存在一定的局限性, 表现在遗迹网络无法绘制生物扰动强度, 而生物扰动强度是研究古环境至关重要的生态指标(Reineck, 1967; Gingras et al., 2011)。此外, 仅遗迹网络组成也无法代表相关的岩相和实体化石的信息, 而岩相和实体化石是古环境分析的重要信息。目前, 暂时仅能通过对生物扰动强度、岩相特征以及实体化石的半定量和定性观察, 再与遗迹学相结合的手段, 进行古环境和生态学的研究(Baucon et al., 2015)。虽然遗迹网络分析这一方法为笔者研究遗迹化石提供了新的思路, 但仍需要不断地进行完善和创新。
以遗迹化石为研究对象, 以遗迹网络分析为手段, 并结合沉积相特征, 探讨了甘南地区上二叠统毛毛隆组遗迹群落的生态环境变化。在上二叠统毛毛隆组可识别出遗迹化石12属17种, 他们包括: Arenicolites isp、Chondrites intricatus、Chondrites isp.、Dictyodora cf. scotica、Dictyodora zimmermanni、Diplichnites triassicus、Diplichnites isp.、Helminthopsis tenuis、Helminthopsis isp.、Lockeia siliquaria、Nereites missouriensis、Palaeophycus tubularis、Palaeophycus isp.、Phycodes isp.、Planolites isp.、Protovirgularia isp.、Scolicia strozzii。根据遗迹化石的组分、分布特点、丰富程度以及网络拓扑分析, 可划分3个遗迹化石组合, 即Palaeophycus-Lockeia-Arenicolites-Dictyodora-Planolites-Protovirgularia-Chondrites组合(C1), Palaeophycus-Lockeia-Arenicolites-Dictyodora-Protovirgularia-Phycodes-Chondrites-Helminthopsis组合(C2), 和Palaeophycus-Nereites-Scolicia-Protovirgularia-Helminthopsis组合(C3)。通过遗迹网络和岩相的综合分析, 认为研究区的古生态环境属于大陆斜坡沉积环境, 这一时期遗迹群落经历了从近源浊流区至远源浊流区的演变历程。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|