




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
寒武纪丝状蓝细菌生物膜主导和诱发的巨型鲕粒:以河南新安县石井剖面苗岭统徐庄组为例*
[梅冥相1, 2  ]
]
]
|
|

作者简介 梅冥相,男,1965年生,教授,博士生导师,主要从事沉积学和地层学研究工作;E-mail: meimingxiang@263.net。
多样化的产出环境和30×108年的分布历史表明,鲕粒成因一直是一个谜一样的沉积学难题,争论的关键问题是其究竟是有机(微生物)成因还是无机成因。来自于华北地台寒武系苗岭统徐庄组鲕粒滩相灰岩顶部的方解石巨鲕表现出同心状、放射—同心状和泥晶质的沉积组构,在鲕粒核心和鲕粒皮层中保存着精美的葛万菌( Girvanella)化石所主导的光合作用生物膜的钙化作用残余物,这为研究鲕粒形成与光合作用生物膜之间复杂的成因联系提供了一个较为直接的微生物证据,因为葛万菌可相对较为肯定地类比于近代的钙化织线菌( Plectonema)丝状蓝细菌化石。所以说,在伴随着后生动物辐射的寒武纪蓝细菌繁盛的方解石海中,尽管形成放射状鲕粒皮层的放射纤维状方解石的沉淀作用机理还没有得到完全彻底的了解,但是这些巨鲕确实表现出光合作用生物膜诱发、滋养并促进了鲕粒形成的直接证据,进一步支持了“鲕粒沉积可以作为一种不同的微生物岩体系新类型”的重要科学理念。
About the author Mei Ming-Xiang,born in 1965, graduated from China University of Geosciences in 1993,and obtained Ph.D. degree. Now he is a professor at School of Earth Sciences and Resource,China University of Geosciences(Beijing),and is engaged in researches on sedimentology and stratigraphy. meimingxiang@263.net.
Diversified forming environments and 3000 million years of history clearly express that ooid is an enchanting depositional grain whose origin remains a dilemma,and its formation is still actively debated that is reflected as the strongly difference between the organic and inorganic origins of ooids. Giant ooids from the top part of the Miaolingian Xuzhuang Formation at the Shijing section in Xin'an County of Henan Province are made of calcite and show concentric,radial-concentric,and micritic fabrics. Within both the core and the cortex of these ooids there are excellent calcified residues of photosynthetic biofilms reflected as the high-density preservation of the Girvanella,which provide a relative direct evidence of ooid formation that is genetically related to the photosynthetic biofilm,since it is relatively certain that Girvanella resembles closely recent calcified Plectonema. Therefore,although the exact forming mechanism of radial-fibrous calcites making up the cortex of radial ooids in the calcite sea of the Cambrian that is marked by cayanobacterial blooms and is coincided with metazoan radiation is not wholly understand,direct evidences of that the formation of these giant ooids is dominated,nourished and promoted by photosynthetic biofilms further support an important scientific idea, i.e. ooid deposits can be understood as a new differentiated type of microbialite system. Ultimately,our finding and study provide an important example for the further understanding and elucidating of the “ooid dilemma”.
鲕粒是圆形的或卵圆形的包覆颗粒(直径常常小于2 mm), 具有一个围绕着核心增生的平滑而且成层性较好的皮层, 核心常常由生物碎屑、球粒、硅质碎屑颗粒或岩石碎屑所组成(Davies et al., 1978; Simone, 1981; Tucker and Wright, 1990)。直径大于2 mm的鲕粒, 常被定义为“ 巨鲕” (Sumner and Grotzinger, 1993; 梅冥相, 2008, 2012a, 2012b; 李飞等, 2010; Trower and Grotzinger, 2010; 代明月等, 2014; Tang et al., 2015; Thorie et al., 2018)。传统观念认为, 鲕粒以平滑的圈层、以及“ 藻丝体” 等的缺乏所代表的非有机成因特征区分于有机成因的核形石, 其中平滑的圈层是区分这2种包覆颗粒的可靠标准(Davies et al., 1978; Simone, 1981; Tucker and Wright, 1990; Gerdes et al., 1994), 然而典型的放射鲕也常常缺乏平滑的圈层(梅冥相等, 2019c), 而且在寒武纪的核形石(Liu and Zhang, 2012; Han et al., 2015; Wilmeth et al., 2015; 齐永安等, 2016; 梅冥相等, 2019a, 2019b, 2020a)和鲕粒(Liu and Zhang, 2012; 代明月等, 2014; 陈百兵等, 2019; 梅冥相等, 2020b)中的“ 藻丝体” 还应该修订为丝状蓝细菌的鞘化石。
自从Sorby(1879)提出鲕粒的生长形成是像“ 雪球” 一样的机械过程这一认识以来, 有关鲕粒形成的机理几乎争论了150年, 主要表现为鲕粒的有机成因(Fabricius, 1977)和无机成因(Davies et al., 1978)的剧烈争论, 这在早期的综述中得到了系统的总结(Bathurst 1975; Simone, 1981)。Tucker和Wright(1990)的重要认识, 即“ 将鲕粒完全归为物理成因似乎为时过早” , 可能成为在新世纪之交研究鲕粒形成的有机或生物过程的推动力, 体现在以下几个创造了一些新概念的研究实例之中: (1)对现代微生物席中的多样化碳酸盐颗粒结构的研究, Gerdes等(1994)构筑了“ 底栖鲕粒” 的概念, 而且强调微生物席内的核形石和鲕粒是同一种微生物过程(生物膜聚集作用和文石结壳作用的交互过程)的变异体, 鲕粒与核形石的有效区分是前者具有平滑的皮层而后者具有扰动的皮层, 并认为这可能取决于核心的球状规则性; (2)Reitner等(1997)强调大盐湖(美国犹他州)中的现代文石鲕粒是有机矿化作用过程的产物, 而且发现与盐湖中较为罕见的文石放射鲕粒形成相关的有机质来源包括碎屑质的终年被风吹的有机质、昆虫(Ephydra)的残余物以及微生物多糖, 而细菌和蓝细菌没有直接卷入它们的形成作用; (3)Castanier等(1999)的研究表明, 在富营养之后, 细菌活度在固体产物形成早期和开始期很高, 而且生物矿物聚集体具有特别差的晶体结构而受到奢华的生物过程所掩盖, 这种条件适合于具有放射状内部结构(即放射鲕粒)的圆形体的形成; (4)在对北海现代微生物席的研究过程中, Brehm等(2006)多次观察到了不寻常的活着的生物微球, 这些生物微球由丝状蓝细菌(Phormidium sp.)、以及相关联的以一个特定方式排列的硅藻如Navicula属和异养细菌所构成, 而且在这些协同的培养基中发现了与化石鲕粒形式相似的碳酸盐沉淀作用, 从而提出了一个鲕粒形成的微生物过程的认识和假说; (5)Kahle(2007)在对巴哈马现代文石鲕粒的研究中, 认为围绕着核心的细菌生物膜、以及在生物膜中的可用于细菌的营养物供应, 是控制鲕粒形成的基本要素。但是, 也基于对巴哈马现代鲕粒的研究, Duguid等(2010)认为鲕粒的形成与微生物活动不存在直接关系, 重新强调了鲕粒形成的化学过程, 这增加了对鲕粒形成机理理解的困难, 并造成更加激烈的争论。
近年来, 更加深入的研究表明了鲕粒形成的有机过程尤其是微生物过程, 包括: (1)Cody和Noel(2012)对发育在澳大利亚南部Nullarbor平原的中新世沼泽相鲕粒的研究表明, 在表生背景之中这些球形纹层状颗粒的形成经历了土壤过程和微生物过程, 而且发现EPS、藻丝体、坡缕石和海泡石构成的纳米纤维微生物席主导了鲕粒皮层的微结构; (2)来自于日内瓦湖泊淡水鲕粒的研究表明了一个光合作用生物膜驱动鲕粒形成的本质性作用(Plé e et al., 2010); (3)Pacton等(2012)的进一步研究表明, 光合作用微生物不仅增强了围绕着鲕粒核心的早期碳酸盐沉淀作用, 而且控制了瑞士日内瓦湖泊中的淡水鲕粒整个皮层的形成, 微生物细胞外聚合物质在钙化之前首先透矿化为非晶质镁硅酸盐(amMg-Si), 诸如硫酸盐还原细菌之类的异养微生物在EPS矿化之前不可能在EPS的降解作用中起着明显的作用, 从而与犹他州大盐湖的鲕粒形成(见Reitner et al., 1997)存在着一些差异; (4)很多学者采用新技术对巴哈马现代鲕粒进行深入的和系统的多学科研究, 强调生物成因机制(微生物调节的有机矿化作用)在鲕粒形成中的巨大作用(Summons et al., 2013; Harris et al., 2019), 如: 微生物多样性(Edgcomb et al., 2013)、功能基因多样性(Diaz et al., 2014)、微生物活动的地球化学证据(Diaz et al., 2015)、鲕粒中有机质来源的分子信号(O'Reilly et al., 2017)、鲕粒形成的有机矿化作用(Diaz et al., 2017)以及鲕粒生长的底栖过程(Mariotti et al., 2018)等研究; 在此基础上, Diaz和Eberli(2019)对这些研究成果进行了高度总结, 并将巴哈马现代文石鲕粒理解为纹层状的有机沉积构造, 而且进一步认为其是一种微生物岩体系的新类型; (5)也正是基于巴哈马鲕粒滩分布的空间样式的研究(Rankey and Reeder, 2009, 2011, 2012; Harris et al., 2011, 2015)及鲕粒大小和形状演化的几何学模型(Trower et al., 2017; Sipos et al., 2018), 总结出鲕粒生长的旋转性旋回样式。然而, 由于鲕粒具有多样化的产出环境(从海洋、湖泊到沼泽)和多样化的矿物构成(如铁质鲕粒, 见Salama et al., 2013; 磷酸盐鲕粒, 见Pufahl and Grimm, 2003), 而且最为古老的鲕粒已经从27.2× 108年前(Awramik and Buchheim, 2009)追溯到30× 108年前(Siahi et al., 2017), 同时这些地球上最为古老的鲕粒还表现为多样化的同心状、放射— 同心状、泥晶和放射状鲕粒, 因此更增加了对鲕粒形成确切机理深入理解的困难。尽管鲕粒形成的微生物成因得到了强调, 但是目前确实难以将其统一为一个笼统的鲕粒生长模式。
与显生宙最高的大气圈CO2含量(Berner and Kothavala, 2001)、相对较低但是一直在上升着的大气圈O2含量(Berner et al., 2007; Campbell and Allen, 2008; Lenton et al., 2018; 但是, 还可能存在较大幅度的大气圈O2含量上升, 见Large et al., 2019)相对应的全球性温暖气候(温室效应条件)和方解石海(Sandberg, 1983; Hardie, 1996), 是寒武纪最为特别的环境特征。在这样的背景下, 最为特征的是与后生动物辐射一致的微生物碳酸盐岩丰度在寒武纪— 早奥陶世的增加(Riding, 2006a; Riding et al., 2019), 以及发生在大气圈二氧化碳分压实质上超过10 PAL阈值(Riding, 2006b; Kah and Riding, 2007)的普遍而且特别的蓝细菌活体鞘的钙化作用(Riding, 2011), 所以, 在寒武纪的微生物碳酸盐岩(梅冥相等, 2017, 2019a, 2019c, 2020b; Mei et al., 2020)、核形石(Liu and Zhang, 2012; Han et al., 2015; Wilmeth et al., 2015; 齐永安等, 2016; 梅冥相等, 2019b, 2020a)与鲕粒(Liu and Zhang, 2012; 代明月等, 2014; 梅冥相等, 2020b)中能够见到蓝细菌的钙化鞘化石。
来自于华北地台的河南省新安县石井剖面寒武系苗岭统徐庄组放射鲕粒滩相灰岩顶部的方解石泥晶鲕粒和泥晶— 放射鲕粒, 多为直径大于2 mm的巨鲕。在鲕粒核心和鲕粒皮层中发现了高密度保存的葛万菌化石所主导的光合作用生物膜, 从而提供了一个研究鲕粒形成与光合作用生物膜之间复杂联系的极为宝贵而且是较为直接的微生物活动证据, 为拓宽对寒武纪方解石海中主导着光养热带碳酸盐岩工厂(James, 1997; Schlager, 2003; Michel et al., 2019; 颜佳新等, 2019)中特别的放射— 同心状和泥晶质方解石鲕粒复杂的形成机理的了解以及对“ 鲕粒谜” 的进一步阐释, 提供了较为宝贵而且罕见的实例。
华北地台在寒武纪接受沉积的时间较晚, 大致开始于寒武纪第二世晚期, 而且寒武纪第二世的地层超覆在不同时代的前寒武纪地层之上, 在二者之间形成一个类似于北美地台(Peters and Gaines, 2012)的“ 巨型不整合面” (章雨旭, 2001)。所以, 在寒武纪, 华北地台发育一个特别的沉积序列(冯增昭等, 1990, 2004; Meng et al., 1997; Lee and Chough, 2011; 马永生等, 2017; Woo et al., 2019; 梅冥相等, 2017, 2019a, 2019b, 2019c, 2020a, 2020b; Mei et al., 2020), 即从寒武系第二统红层与碳酸盐岩混合沉积、经苗岭统鲕粒滩主导的碳酸盐岩台地序列变化到芙蓉统碳酸盐泥所主导的碳酸盐岩台地沉积序列。
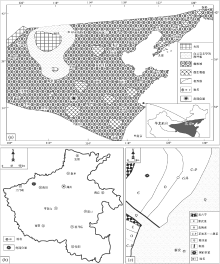
毛庄组、徐庄组、张夏组和崮山组大致构成了华北地台的苗岭统(彭善池, 2009; Peng et al., 2012; 彭善池和赵元龙, 2018)。徐庄组代表着一个碳酸盐岩台地转变的重要地层序列, 在由该组构成的三级层序中, 凝缩段单元表征的快速海侵事件终止之前发育红层的混合沉积, 意味着一个巨型的碳酸盐岩台地进入到成熟阶段(Meng et al., 1997; 马永生等, 2017; 梅冥相等, 2017, 2019a, 2019b, 2019c, 2020a, 2020b; Mei et al., 2020)。河南省新安县石井镇剖面的徐庄组(图 1; 图 2)构成一个三级沉积层序, 海侵体系域由一套具有红层性质的紫色砂质泥岩组成, 厚度约30 m, 其中所夹的碳酸盐岩透镜体和透镜层自下而上从白云岩经灰岩演变到鲕粒灰岩, 代表着一个沉积环境逐渐加深的退积序列; 1层厚度为5 m左右的深灰色和灰绿色陆棚相钙质泥岩夹泥灰岩地层组成该层序的凝缩段, 代表了1次快速海侵事件; 在凝缩段之上, 1套厚度近80 m的块状高能鲕粒滩相地层组成了该层序的高水位体系域, 其下部的早期高水位体系域发育较多的陆棚相钙质泥岩和深缓坡相中薄层泥灰岩, 上部的晚期高水位体系域(LHST)或强迫型海退体系域(FRST)则以发育具大型板状交错层理(图 2-b)的高能鲕粒滩相灰岩为主, 较为清楚地表明鲕粒滩的发育过程对应着基准面(或相对海平面)下降过程(Hunt and Tucker, 1992; Helland-Hansen and Gjelberg, 1994; 梅冥相和杨欣德, 2000; Schlager and Warrlichw, 2009; 梅冥相, 2010; Desjardins et al., 2012; Samanta et al., 2016; Mei and Liu, 2017)。该地层序列不符合经典的层序地层模式(Mitchum et al., 1977; Vail et al., 1977), 但符合Schlager和Warrlichw(2009)所倡导的下降阶段体系域模型, 即与较慢的侵蚀作用和基准面下降速率相对应的较高的沉积物生产作用。再者, 在徐庄组构成的三级沉积层序的高水位体系域的形成时期, 华北地台分布着一个面积大于100× 104 km2的鲕粒滩, 其代表着寒武纪类型多样的碳酸盐岩台地类型中(Pratt et al., 2012)的一种新的碳酸盐岩台地类型(图 1-a), 即鲕粒滩主导的浅海碳酸盐岩陆架, 向海一侧为缓坡、向陆一侧为潮坪(马永生等, 2017; 梅冥相等, 2019a, 2020a-b), 该沉积相分布也不符合经典的威尔逊碳酸盐岩相模式。下文将要介绍的巨鲕, 主要发育在该套块状鲕粒滩相灰岩的顶部, 地层厚度大致为2 m(图 2)。
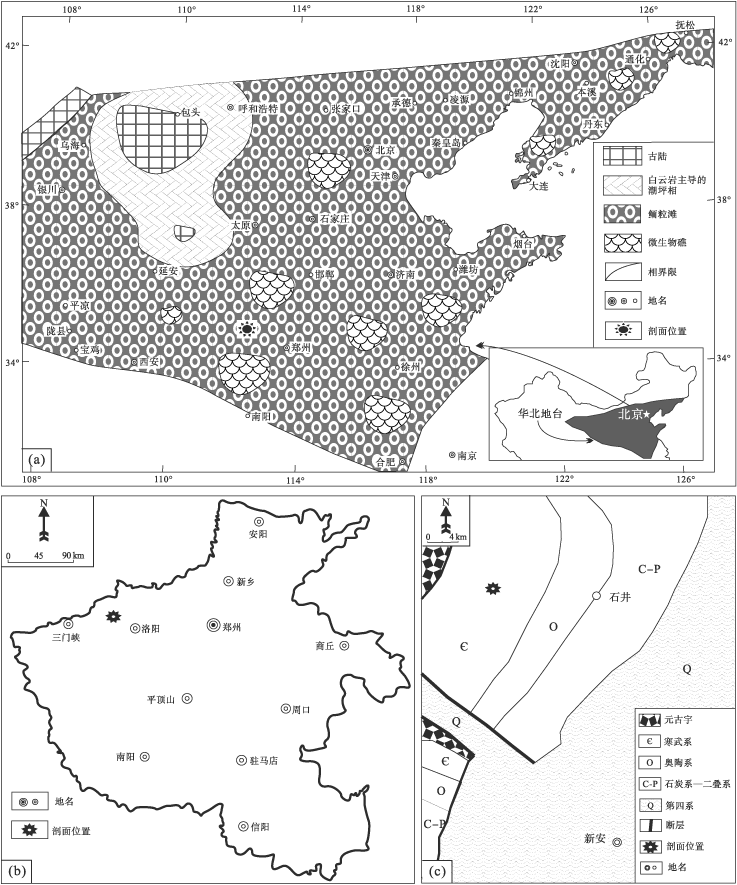 | 图 1 河南省新安县苗岭统徐庄组石井剖面位置及区域地质特征 a— 华北地台徐庄组沉积晚期的概略性古地理图, 修改自冯增昭等(1990, 2004); b— 河南省轮廓; c— 新安县附近地质简图Fig.1 Diagrams showing location of Shijing section for the Miaolingian Xuzhuang Formation in Xin'an County of Henan Province and regional geological characteristics |
 | 图 2 河南新安石井剖面苗岭统徐庄组沉积序列 TST代表海侵体系域, CS代表凝缩段, EHST代表早期高水位体系域, LHST代表晚期高水位体系域, FRST为强迫型海退体系域。照片所代表的沉积现象分别是: a— 徐庄组底部的层序界面(箭头所示), 以毛庄组顶部的鲕粒滩相灰岩与徐庄组底部的具有红层性质的紫色砂质泥岩地层之间的突然变化为特征; b— 徐庄组中部放射鲕组成的颗粒滩相灰岩中的双向板状交错层理; c— 徐庄组顶部的层序界面(箭头所示), 表现为徐庄顶部的块状鲕粒滩相灰岩地层与张夏组底部的陆棚相泥岩和泥灰岩地层之间的突然变化Fig.2 Diagram showing depositional succession for the Miaolingian Xuzhuang Formation at Shijing section in Xin'an County of Henan Province |
河南省新安县石井剖面(位置见图 1)的高水位块状鲕粒滩相灰岩(图 2-b, 2-c; 图 3-a, 3-b, 3-c), 大型双向交错层理发育(图 2-b; 图 3-a, 3-b)是其基本特征, 直径小于2 mm的放射鲕粒是其基本构成(图 3-d, 3-e), 也见到少量的泥晶鲕粒、三叶虫化石以及泥晶套包裹的可能由棘皮生物碎屑构成的假鲕。
 | 图 3 河南新安石井剖面苗岭统徐庄组上部高水位鲕粒滩相灰岩的基本特征 a— 块状鲕粒灰岩远照; b— 鲕粒灰岩中的双向板状交错层理; c— 鲕粒灰岩近照; d— 鲕粒灰岩的低倍显微照片, 其中(1)为放射鲕, (2)为泥晶— 放射鲕或泥晶鲕粒, (3)为三叶虫化石; e— 鲕粒灰岩的高倍显微照片, 其中(1)为泥晶套包覆的可能为棘皮生物碎屑组成的假鲕, (2)为放射鲕Fig.3 Images showing fundamental feature of high-stand grain-bank oolites in the upper part of Miaolingian Xuzhuang Formation at Shijing section in Xin'an County of Henan Province |
在厚度近80 m的高水位鲕粒滩相灰岩的顶部, 发育厚约2 m的粗鲕粒灰岩(图 4)。粗鲕粒的直径常大于2 mm, 多数在2~6 mm之间, 表现出“ 巨鲕” 的特征(Sumner and Grotzinger, 1993; Trower and Grotzinger, 2010; 李飞等, 2010; 梅冥相, 2008, 2012a, 2012b; 代明月等, 2014; Tang et al., 2015; Thorie et al., 2018) , 相对较为平滑的鲕粒皮层明显区别于核形石(Tucker and Wright, 1990; Liu and Zhang, 2012; Han et al., 2015; Wilmeth et al., 2015; 齐永安等, 2016; 梅冥相等, 2019a, 2019b; 梅冥相等, 2020a)。在宏观特征上与下伏的大套细粒放射鲕粒灰岩(图 3)形成明显的反差, 二者之间紧密的共生关系和较为相似的潮下高能环境, 代表着较为壮观的沉积学现象, 因为经典而传统的观念将方解石放射鲕粒解释为形成于高镁方解石而且多形成于低能环境(Davies et al., 1978; Simone, 1981; Tucker and Wright, 1990; Reitner et al., 1997; Flü gel, 2004; Woods, 2013; Siahi et al., 2017; Diaz and Eberli, 2019)。最为重要的是, 在巨鲕构成的颗粒灰岩之中, 局部发育厘米至分米级大小的小型均一石(图 4-b)和叠层石(图 4-c)生物丘, 但由于采样较为困难, 所以本次研究没有获得这些生物丘的显微观察样本, 有待于今后进一步采样来进行系统研究。
 | 图 4 河南新安石井剖面苗岭统徐庄组顶部巨鲕灰岩的基本宏观特征 a— 巨鲕颗粒灰岩的近照; b— 巨鲕灰岩中的小型均一石生物丘(箭头所指); c— 巨鲕灰岩中的小型叠层石生物丘Fig.4 Images showing macroscopically fundamental feature of giant-ooid oolites in the top part of Miaolingian Xuzhuang Formation at Shijing section in Xin'an County of Henan Province |
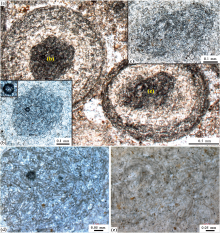
与大套鲕粒灰岩中主导性鲕粒为放射鲕粒(图 3-d, 3-e)明显不同, 图 4所示的石井剖面徐庄组顶部的巨鲕, 除了具有较大的直径(常常大于2 mm)之外, 主要表现为同心状、放射— 同心状、泥晶和放射状的鲕粒组构(图 5), 核心包括泥晶质的凝块(图 5-a)、三叶虫化石(图 5-b)和小型放射鲕粒(图 5-d, 5-e)。鲕粒皮层由相对较为平滑和断断续续的放射纤维状方解石纹层和泥晶纹层所组成: 放射纤维状方解石纹层的厚度相对较小(数十微米到200 μ m), 向鲕粒边缘变薄乃至消失; 泥晶纹层的厚度相对要大(可以达到毫米级别), 朝向鲕粒边缘泥晶纹层逐渐加厚并占据着主导。另外, 还可见到通过泥晶纹层包覆而形成的数毫米大小的葡萄石(图 5-c)、厚度超过1 mm的外部块状泥晶皮层主导的同心状放射— 泥晶鲕粒(图 5-d)和边缘明显受到磨损而破碎的巨鲕(图 5-e)。在显微组构中表现出的相对较为平滑的泥晶和放射纤维状方解石鲕粒皮层, 是其区别于核形石(Tucker and Wright, 1990; Liu and Zhang, 2012; Han et al., 2015; Wilmeth et al., 2015; 齐永安等, 2016; 梅冥相等, 2019a, 2019b; 梅冥相等, 2020a)的最为基本的特征, 另外, 单个鲕粒总是表现为从内向外由同心— 放射组构向主导性的致密泥晶组构变化的特点。
 | 图 5 河南新安石井剖面苗岭统徐庄组顶部巨鲕的基本微观特征 a— 泥晶质凝块为核心的同心状放射— 泥晶巨鲕, 大小为3 mm左右; b— 三叶虫化石碎屑核心的同心状放射— 泥晶巨鲕, 大小为5 mm左右, 在巨鲕之间填充着小型放射鲕粒; c— 泥晶纹层包覆着3个小型放射鲕和泥晶鲕粒的葡萄石, 大小为6 mm左右; d— 块状致密泥晶构成外侧皮层的同心状放射— 泥晶巨鲕, 大小为4 mm左右; e— 边缘明显受到磨损的同心状放射— 泥晶巨鲕, 大小为5 mm左右Fig.5 Images showing microscopically fundamental feature of giant-ooids in the top part of Miaolingian Xuzhuang Formation at Shijing section in Xin'an County of Henan Province |
在低倍显微镜下, 由泥晶凝块(图 5-a; 图 6-a)所构成的巨鲕核心多为次棱角状, 大小在0.5 mm左右, 与鲕粒皮层之间的边界较为清晰, 从而成为方解石放射鲕粒皮层的生长基质。在高倍显微镜下, 该核心表现为高密度保存的缠绕状葛万菌(Girvanella)化石(图 6-b至6-e), 包覆着该核心的鲕粒皮层内部以放射纤维状方解石皮层为主, 向外则变化为由泥晶质皮层所主导。在鲕粒核心泥晶团块中所见到的葛万菌化石由高度缠绕的丝状体所组成, 直径为10 μ m左右, 长度可以达到毫米级别, 相对均匀而且较薄(1~2 μ m)的泥晶壁构筑和成型了这些丝状化石的基本形态, 且泥晶壁以相对更加强烈的暗色而区别于填充在丝状体之间的泥晶。另外, 还见到了少量球形蓝细菌化石(图 6-b, 6-d), 直径为30 μ m左右, 具有较厚的泥晶壁和中空的亮晶填充物, 表明可能属于粘球菌之类的球形蓝细菌化石。
 | 图 6 河南新安石井剖面苗岭统徐庄组顶部巨鲕核心的特征性微观特征 a— 2个以泥晶团块为核心的同心状放射— 泥晶鲕粒的低倍照片; b, c— 为照片a中的巨鲕核心的高倍显微镜照片, 见到1个可能的球形蓝细菌化石(b), 表现为高密度保存的高度缠绕的丝状葛万菌化石; d, e— 为巨鲕核心的进一步放大, 除了更加清楚地丝状葛万菌化石以外, 那些很小的(1~2 μ m)的黑色棱角状小点可能代表着具有特殊意义的黄铁矿晶体的残余物Fig.6 Images showing distinctively microscopically fundamental feature for core of giant-ooids in the top part of Miaolingian Xuzhuang Formation at Shijing section in Xin'an County of Henan Province |
在巨鲕厚度大小不一而且形态多样的泥晶质皮层之中, 也发现了高密度保存的葛万菌化石(图 7; 图 8)。这些高密度保存葛万菌化石的鲕粒和葡萄石的泥晶质皮层, 在低倍显微镜下大致表现为数十到数百微米厚的纹层状(图 7-a; 图 8-d)、不规则团块状(图 7-d; 图 8-d)、块状纹层(图 8-a); 在高倍显微镜下, 则表现为由高度缠绕而且高密度保存的丝状葛万菌化石构成(图 7-b, 7-c, 7-e, 7-f; 图 8-b, 8-c, 8-e, 8-g)。此外, 该泥晶质皮层还表现出以下特征: (1)多发现于鲕粒的外侧, 具有朝向一侧加厚而不对称的特点(图 7-a; 图 8-a); (2)与较薄而且不连续的放射纤维状方解石纹层交互产出; (3)常在鲕粒最外部, 最厚而且最为特征, 表现为建设性的泥晶套(Riding, 2002)特征。
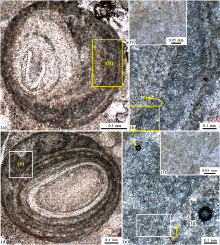
 | 图 7 河南新安石井剖面苗岭统徐庄组顶部巨鲕的纹层状和非纹层状巨鲕皮层 a— 1个以三叶虫化石碎片为核心的偏心巨鲕, 直径为2.5 mm左右, 纹层状的泥晶皮层向右侧加厚; b, c— 照片(a)中长方形所示部分的巨鲕皮层的局部放大, 表现为高密度保存的葛万菌化石; d— 1个以小型放射鲕为核心的偏心巨鲕, 在左上部的泥晶鲕粒皮层之中包含着不规则状的暗色泥晶团块和微亮晶物质; e, f— 照片(d)中长方形框所示部分的进一步放大, 表现为高密度保存的丝状葛万菌化石; g— 照片(e)中(箭头所指)所见到的1个可能的球形蓝细菌化石。另外, 在照片(c)和(f)所示的高倍显微镜照片中所见到的那些很小的 (1~2 μ m)的棱角状黑色小点可能代表着具有特殊意义的黄铁矿晶体的残余物Fig.7 Images showing laminated and irregular-like cortex of giant-ooids in the top part of Miaolingian Xuzhuang Formation at Shijing section in Xin'an County of Henan Province |
 | 图 8 河南新安石井剖面苗岭统徐庄组顶部块状泥晶纹层构成的巨鲕和葡萄石 a— 1个以块状毫米级别的致密泥晶纹层为特征的次圆形巨鲕, 大小为3 mm左右; b, c— 照片(a)中长方形所示部分的巨鲕皮层的局部放大, 表现为高密度保存的葛万菌化石; d— 1个较为典型的葡萄石, 以3个小型鲕粒为核心, 被泥晶皮层所包覆, 大小为6 mm左右, 也见图 5-c; e至g— 照片(d)中长方形框所示部分的进一步放大, 表现为高密度保存的丝状葛万菌化石。在照片(c)、(e)和(f)所示的高倍显微镜照片中所见到的那些很小的(1~2 μ m)的黑色棱角状小点可能代表着具有特殊意义的黄铁矿晶体的残余物Fig.8 Images showing giant-ooid marked by a massive dense-micrite contex and grapestone in the top part of Miaolingian Xuzhuang Formation at Shijing section in Xin'an County of Henan Province |
与巨鲕核心致密泥晶团块或凝块(低倍显微镜下)中的高密度保存的丝状葛万菌化石(图 6)相似, 在徐庄组巨鲕和巨型葡萄石的致密泥晶质皮层(图 7-a至7-d; 图 8-a至8-d)中, 也发育着高密度保存的丝状葛万菌化石。尽管都笼统归为葛万菌, 但作为一个化石的属, 还应该进一步描述为种, 但这已经超出了文中的研究范围, 有待于今后继续调查和研究并求教于古生物学家。然而, 那些相对较粗的缠绕状而且相对较粗的丝状体化石(图 8-e, 8-g)可能应该描述为Girvanella kasakiensis, 其余相对较细的丝状体化石大致可以归为Girvanella problematica(参见Liu et al., 2016)。同时, 在鲕粒皮层的泥晶质组构之中也见到球形蓝细菌化石(图 7-e, 7-g), 暗色致密泥晶构成的中空的球状化石可以类比于现代发育鞘的粘球菌之类的球形蓝细菌, 尽管它们的生物亲和性还需要进一步研究。因此, 徐庄组顶部的高密度保存着葛万菌化石的泥晶质巨鲕皮层, 表现出较为典型的微生物成因的基本特征。
在经典而传统的观念中, 放射鲕粒被解释为形成于高镁方解石而且多形成于低能环境(Davies et al., 1978; Simone, 1981; Tucker and Wright, 1990; Reitner et al., 1997; Flü gel, 2004; Woods, 2013; Siahi et al., 2017; Diaz and Eberli, 2019), 但是, 在直径小于2 mm的放射鲕粒(图 3-d, 3-e)所主导的徐庄组强迫型海退体系域的块状鲕粒灰岩中, 大型板状交错层理尤其是双向交错层理普遍发育(图 2-b; 图 3-a, 3-b), 表明这些块状放射鲕粒灰岩应该属于潮下高能沉积。由其所表征的大型鲕粒滩复合体分布面积大于100× 104 km2(图 1-a), 不仅表明1个由鲕粒主导的碳酸盐岩台地发育在华北地台的苗岭世(马永生等, 2017; 梅冥相等, 2019a, 2020a, 2020b), 而且指示这个巨大的鲕粒砂滩的产出和发育环境类似于今天巴哈马的鲕粒砂滩复合体(Rankey and Reeder, 2009, 2011, 2012; Harris et al., 2011, 2015, 2019)。这是1个在地质历史时期分布面积最大而且较为罕见的鲕粒滩。
河南省新安县石井剖面徐庄组顶部的巨鲕(图 4)集中发育在三级沉积层序强迫型海退体系域沉积块状鲕粒灰岩顶部(图 2-c), 局部与小型均一石(图 4-b)和叠层石(图 4-c)共生在一起, 间接地表明这些巨鲕是“ 纹层状的有机沉积构造” , 并遵循着微生物岩体系的一些形成特征。这是Diaz 和Eberli(2019)从巴哈马现代鲕粒所表现出的微生物活动特征推论出的一个重要科学理念。
石井剖面徐庄组顶部的巨鲕具有较为特别的同心状、放射— 同心状和泥晶质的沉积组构(图 5), 还包含少量的葡萄石(图 5-c), 总是留下方解石海(Sandberg, 1983; Hardie, 1996)中形成的方解石放射鲕(Siahi et al., 2017)的痕迹。但是, 更加发育的泥晶质皮层又表现出较为典型的皮壳粒(Cortoid; 由建设性泥晶套所包覆的颗粒; Gerdes et al., 1994; Flü gel, 2004; Woods, 2013)的基本特征。所以说, 不管是矿物构成还是沉积组构, 其均与现代的文石泥晶鲕粒有巨大的差异。
最为特征的是, 在这些巨鲕的致密泥晶质的凝块状核心(图 6)以及在那些纹层状(图 7-a; 图8-d)、不规则团块状(图 7-d; 图8-d)和块状致密泥晶(图 8-a)构成的巨鲕皮层之中, 发育着高密度保存的葛万菌化石。这些高度缠绕而且不分叉的丝状化石, 以下特征表明确实是较为典型的葛万菌化石(图 6; 图 8), 符合Nicholson和Etheridge(1878)提出的定义, 这些特征包括: (1)致密缠绕而且不分叉的丝状体; (2)丝状体的直径多为10 μ m左右, 长度可以达到数百微米; (3)较薄的(1~2 μ m)暗色致密泥晶壁成型了化石的轮廓, 在高倍显微镜下表现出比化石间的泥晶更暗的色调和更加致密的泥晶构成(图 7-c, 7-f; 图 8-c, 8-f, 8-g); (4)俯卧式的生长样式。
需要指出的是, 这样的葛万菌(图 6至图 8)属于葛万菌组合(丝状而且不分叉的组合)中最具有代表性的类型, 因为葛万菌组合的化石还包括束线菌(Subtifioria, 直线状的和电缆线状的)、环绕菌(Obruchevella, 线圈状环绕的)以及拉祖莫夫菌(Razumovskia; 柔软的、纤细的而且为卧伏式的细丝体)(Riding, 1991a, 1991b), 而且在华北地台寒武纪的凝块石(梅冥相, 2019d; Mei et al., 2020)和核形石(梅冥相, 2019a, 2020a)中还见到类似于葛万菌、但是确实是非缠绕的丝状蓝细菌鞘化石(可以类比于现代第谷菌(Tychonema; De los Rí os et al., 2015)的新的化石类型); 因此, 在徐庄组巨鲕的核心和皮层中见到的较为典型的葛万菌, 表明较为单调的化石组合类型是其基本特征之一。在早期的研究中, Vologdin(1962)将葛万菌(Girvanella)置于蓝细菌, 但是Johnson(1966)还是喜欢将其归到绿藻, 而Riding(1977)将Aldabra环礁的淡水水池中织线菌(Plectonema)的钙化鞘作为葛万菌的近代类比物, 且这样的解释一直沿用至今(Riding, 1991a, 1991b, 2011; Liu and Zhang, 2012; Liu et al., 2016; 梅冥相, 2017, 2019a, 2019b, 2019c, 2019d, 2020a, 2020b; Mei et al., 2020), 尽管类似于葛万菌(Girvanella)的缠绕状丝状蓝细菌鞘化石还被类比于现代的伪枝菌(Scytonema; 齐永安等, 2017)。图 7和图 8所示的徐庄组顶部巨鲕的核心和皮层中的葛万菌化石, 尽管准确的属种定名还需要进一步研究, 但是可以较为肯定的是该葛万菌化石确实代表着丝状蓝细菌鞘的钙化作用残余物。这些丝状蓝细菌鞘化石常具有1个厚度均匀(1~2 μ m)的暗色泥晶壁, 该暗色泥晶壁可能是丝状蓝细菌鞘的活体钙化作用残余物, 而且代表着直接与大气圈二氧化碳浓缩作用机制(CCMs)相联系的生态生理学机制(Kah and Riding, 2007; Riding, 1991a, 1991b, 2011), 而内部被亮晶方解石交代的管可能是蓝细菌细胞列(香毛簇)的降解作用残余物(Riding, 1991a, 1991b, 2011), 故这种蓝细菌活体鞘的钙化作用成型了葛万菌化石的基本轮廓。所以, 较为肯定的是, 徐庄组顶部巨鲕的致密泥晶质的凝块状核心(图 6)、以及由纹层状(图 7-a; 图 8-d)、不规则团块状(图 7-d; 图 8-d)和块状致密泥晶(图 8-a)构成的巨鲕皮层, 实际上就是光合作用生物膜(Riding, 2002; Flemming et al., 2016)的钙化作用残余物。
因此, 可以进一步肯定的是, 光合作用生物膜在驱动徐庄组顶部巨型鲕粒的形成和生长过程中具有本质性的作用, 类似于今天的日内瓦淡水湖泊中的方解石鲕粒(Plé e et al., 2010; Pacton et al., 2012)。但是, 又表现出以下特殊性和复杂性: (1)紧接着葛万菌生物膜核心的放射纤维状方解石的沉淀作用(图 6-a)以及与鲕粒葛万菌主导的泥晶皮层交互的放射纤维状方解石鲕粒皮层(图 5, 图 7, 图 8), 尽管其确切机制还需要进一步调查, 但是还是反映了在富有机质和富营养物条件所激发的放射纤维状方解石的沉淀作用特征(Castanier et al., 1999; Richter et al., 2011), 类似于犹他州大盐湖中的现代文石放射鲕粒的形成作用(Reitner et al., 1997); (2)朝向一侧(图 7-a, 7-d)以及从里向外(图 6-a; 图 8-a)加厚的巨鲕和葡萄石(图8-d)的葛万菌主导的光合作用生物膜皮层(图 5)所展示的不规则皮层的特点, 包括图 5-e所示的鲕粒边缘明显的磨损作用, 表明巨鲕形成过程中底栖(滚动)过程的贡献, 类似于Mariotti等(2018)所强调的现代巴哈马文石鲕粒形成的底栖过程的贡献。该底栖过程可能得益于与鲕粒共生的小型均一石(图 4-b)和叠层石(图 4-c)生物丘的障积作用, 进一步表明巨型鲕粒生长也经历了许多学者所强调的鲕粒生长所要求的动荡与停息阶段的交互过程(Rankey and Reeder, 2009, 2011, 2012; Trower et al., 2017; Sipos et al., 2018; Harris et al., 2011, 2015, 2019; Diaz and Eberli, 2019), 同时也表明形成巨型鲕粒的海底环境还可能生长着一些特别的多重状蓝细菌生物膜构成的微生物席, 进一步说明了寒武纪海洋中的蓝细菌繁荣所激发的多样化微生物席或微生物膜的建造物特征。
最为重要的是, 不能够简单地将徐庄组顶部光合作用生物膜主导的方解石巨鲕直接理解为蓝细菌的建造物, 其更可能为一个复杂的蓝细菌(丝状蓝细菌为主, 也可能包含一些球形蓝细菌(图 6-b; 图 7-e, 7-g))主导的生物膜的早期石化作用(Dupraz et al., 2009, 2011)的产物, 因为并非所有蓝细菌所主导的微生物膜或微生物席均能够钙化, 而且今天的绝大多数微生物膜或微生物席确实没有形成坚硬的岩石(Stal, 2012)。虽然还存在很多细节问题需要进一步调查和研究, 但构成鲕粒核心与皮层的光合作用生物膜的细胞外聚合物质(EPS; Decho, 2010; Dupraz et al., 2009, 2011; Stal, 2012; Decho and Gutierrez, 2017)、以及由EPS所主导的蓝细菌细胞外基质(ECM; Helm and Potts, 2012)复杂的钙化作用, 是徐庄组顶部方解石巨鲕形成的主要作用机制。光合作用生物膜的钙化作用主导的巨鲕经历了复杂的作用过程, 包括: (1)蓝细菌鞘可能的活体钙化作用(Kah and Riding, 2007; Riding, 1991a, 1991b, 2011), 这可能首先发生并成型了图 6至图 8所示的葛万菌化石; (2)蓝细菌丝状体之间的间隙中致密泥晶和微亮晶基质所代表的钙化作用, 尽管该作用过程可能经历了接近活体的蓝细菌微生物席的EPS构成的钙化作用(Ka
虽然对巴哈马现代海相文石鲕粒近年来的持续性研究(详见Harris 等(2019)以及Diaz和Eberli(2019)的高度概括和总结)表明了微生物和细胞外聚合物质(EPS)在鲕粒成因和发育中的作用, 并且这些包覆颗粒可以看作是“ 纹层状的有机沉积构造(laminated organosedimentary structures)” 而且遵循着微生物岩体系的一些形成特征, 但是还是缺乏相应可靠的微生物证据, 所以说, 明确发生在紧密关联的时空条件下的2种鲕粒形成的沉淀作用机制— — 生物诱发的和生物影响的机制在纳米级别到显微尺度下的具体贡献, 现在还是一个挑战。笔者在寒武纪巨型鲕粒中典型的光合作用生物膜的钙化作用残余物之中所发现的高密度保存的丝状蓝细菌化石和少量的球形蓝细菌化石, 为了解方解石海中光合作用生物膜(嵌入在自我生产的EPS中的蓝细菌群落; Decho, 2010; Flemming et al., 2016; Decho and Gutierrez, 2017)滋养和诱发寒武系苗岭统放射— 同心状方解石巨鲕的形成, 提供了较为罕见而且可靠的微生物证据。
陈百兵等(2019)基于超微组构对放射鲕粒中泥晶方解石残余物的微生物成因(蓝细菌死亡之后的灰泥化)的解释, 及宋文天和刘建波(2020)强调的放射鲕粒皮层中的同心状组构, 均间接地表明了放射鲕粒中残留着相应的泥晶薄膜代表的光合作用生物膜的钙化作用残余物, 类似于图 7和图 8所示的放射— 同心状巨鲕的高密度保存着丝状蓝细菌的光合作用生物膜的钙化作用残余物, 从而成为放射鲕粒的生长受到光合作用生物膜诱发和滋养的证据, 尽管还存在许多细节问题需要进一步研究。朝向一侧(图 7-a, 7-d)以及从里向外(图 6-a; 图 8-a)加厚的巨鲕和葡萄石(图8-d)的葛万菌主导的光合作用生物膜皮层(图 5)所展示的不规则皮层, 包括图 5-e所示的鲕粒边缘具有明显的磨损作用, 表明了巨鲕形成作用中的底栖(滚动)过程的贡献, 进一步说明巨型鲕粒形成初期可能产生于水柱中的受到“ 海雪” (生物膜的残余物)提供的有机基质对放射纤维状方解石沉淀作用的滋养和激发, 但在海底的底栖滚动作用期间则生长和发育了更厚的丝状蓝细菌主导的生物膜。因此, 这些巨型鲕粒的形成, 也间接地说明了为什么较大的鲕粒多以同心鲕为主(郭芪恒等, 2020)的原因。
河南新安石井剖面徐庄组顶部的巨鲕, 发育和生长在高能鲕粒颗粒滩相灰岩之上, 组成三级沉积层序强迫型海退体系域, 与鲕粒核心和皮层中的高密度保存着葛万菌化石的光合作用生物膜的钙化作用残余物一起, 形成了较为壮观的沉积学现象, 为拓宽“ 鲕粒谜” 的阐释和了解提供了一个方解石海中的与光合作用新陈代谢活动存在着直接联系的方解石鲕粒的典型实例。
尽管矿物构成存在着差异, 徐庄组顶部的巨鲕与现代文石鲕粒共享着以下2个方面的基本特征, 包括: (1)鲕粒可以看作是“ 纹层状的有机沉积构造” 而遵循着微生物岩体系的一些形成特征; (2)鲕粒形成与成型总体性的样式符合“ 旋转性旋回的样式” 。需要强调的是, 徐庄组巨鲕皮层的增生作用确实偏离了晶体生长的理论, 表现在泥晶质皮层具有明显的光合作用生物膜的钙化作用残余物的基本属性, 而且放射纤维状方解石纹层的沉淀作用受到富有机质和营养物环境的控制, 从而进一步表明生物矿物聚集体具有特别差的晶体结构而受到复杂的生物过程所掩盖。但是, 除了矿物构成与现代海相文石鲕粒不同以外, 徐庄组顶部的巨鲕还具有以下特殊性而表明了鲕粒形成作用的复杂性: (1)尽管鲕粒皮层的增生作用主要由光合作用生物膜及其代表的底栖过程所主导, 但是该光合作用生物膜的钙化作用还与硫酸盐还原反应存在着联系; (2)鲕粒皮层中的放射纤维状方解石皮层经历了特别的沉淀作用, 代表着一个特别的生物化学过程; (3)鲕粒的同心— 纹层状组构不能解释为像现代文石鲕粒那样的方解石晶体的切线状排列的结果; (4)鲕粒较为平滑的皮层更多是磨损作用的产物, 表明了鲕粒的最终成型也受到物理过程的驱动。
就像形成这些巨型鲕粒核心与皮层的光合作用生物膜的钙化作用以及成型了在生物膜中高密度保存的丝状蓝细菌的可能的活体鞘钙化作用那样, 了解和剖析徐庄组巨鲕形成与成型的所有相关作用过程最大的挑战是依据产物的组构和组分特征去推断作用过程的时候会存在着许多偏差, 因为要穿越成岩作用过滤器; 再者, 不同的海水条件(现代的文石海与寒武纪的方解石海)及其所产生的现代文石泥晶鲕粒与寒武纪的方解石鲕粒之间确实存在着形成机理和生长样式的巨大差异, 尽管共享着同一个作用特征。无论如何, 本次研究提供了较为罕见而且可靠的微生物证据来了解寒武纪方解石海之中光合作用生物膜滋养和诱发放射— 同心状方解石巨鲕的形成, 这为今后的深入研究提供了重要的研究线索和思考途径。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|