







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
广西隆安都结剖面下石炭统都安组上部鲕粒类型及其地质意义*
[巩恩普 , 杨臻元, 黄文韬, 关长庆, 张永利, 苗卓伟, 王立芙, 李骁, 王俊杰]
, 杨臻元, 黄文韬, 关长庆, 张永利, 苗卓伟, 王立芙, 李骁, 王俊杰]
, 杨臻元, 黄文韬, 关长庆, 张永利, 苗卓伟, 王立芙, 李骁, 王俊杰]
|
|

第一作者简介 巩恩普,男,1958年生,教授,主要从事古生物学与地层学相关的教学与科研工作。E-mail: gongep@mail.neu.edu.cn。
鲕粒是一类特殊的沉积颗粒,为古气候和古海洋环境的重要指示器。为了深入认识此类特殊颗粒的成因机制、形成环境及地质意义,对广西隆安地区都结剖面下石炭统都安组上部含鲕粒地层开展了古生物学、沉积学和岩相学研究。研究区共识别出5种主要的鲕粒类型: 放射状纹层鲕粒(O1)、规则同心放射状纹层鲕粒(O2)、不规则同心放射状纹层鲕粒(O3)、泥晶鲕粒(O4-A和O4-B)和复合鲕粒(O5)。各类鲕粒的显微组构和沉积环境指示其具有不同的形成过程,其中水动力条件影响和控制着鲕粒的发育和分布情况。研究区含鲕粒地层形成于维宪期末—谢尔普霍夫期,恰好对应早石炭世晚期冰川作用的开始。受冰川作用影响,全球海平面频繁波动,研究区地处低纬度地区并以浅滩和潮坪沉积环境为主,为鲕粒的形成提供了适宜的水体条件,即温暖、动荡的浅水环境。此外,含鲕粒岩层内广泛发育钙质微生物和微生物席,说明微生物活动在研究区较为常见,可能与鲕粒的形成过程具有一定的关联。
, Yang Zhen-Yuan, Huang Wen-Tao, Guan Chang-Qing, Zhang Yong-Li, Miao Zhuo-Wei, Wang Li-Fu, Li Xiao, Wang Jun-Jie
About the first author Gong En-Pu,born in 1958,professor,is mainly engaged in researches of paleontology and stratigraphy. E-mail: gongep@mail.neu.edu.cn.
Ooids,a typical type of sedimentary grains,are an important proxy for paleoclimate and paleoceanography. To better understand the formation mechanisms,depositional environments and geological significance of these carbonate grains,studies relating to paleontology,sedimentology,and petrography were carried out on the ooid-bearing limestones in the upper Du’an Formation of the Lower Carboniferous in Dujie section of Long’an area,Guangxi. Five ooid types have been identified: Radial ooids(O1),regular concentric-radial ooids(O2),irregular concentric-radial ooids(O3),micrite ooids(O4-A,O4-B),and composite ooids(O5). The microscopic fabric and depositional environments of these ooids uncovered different forming processes,indicating that the formation and distribution of the ooids were influenced by the hydrodynamic conditions. These ooids formed in the late Visean to Serpukhovian,corresponding to the onset of the ice age in the late Mississippian. Influenced by changes of ice volume,sea level changed frequently,resulting in shallow shoal and tidal flat environment in the study area. Dujie area was located in the low latitude,providing shallow,warm,and agitated waters for the ooid formation. Furthermore,calcimicrobes and microbial mats were abundant in the ooid-bearing limestone,indicating that the microbial activities were common and might be related to the formation of these ooids.
鲕粒是圆形、椭圆形或卵形碳酸盐岩颗粒, 表面光滑, 粒径一般小于2 mm, 具有核心和皮层, 核心一般为生物碎屑、似球粒、硅质颗粒或者岩屑等, 外部皮层具有很好的纹层结构(Davies et al., 1978; Simone, 1980; Diaz and Eberli, 2019)。富含鲕粒的地层往往具有较高的孔隙度和渗透率, 是良好的油气聚集场所, 常被视为油气勘探的重点(马永生等, 2005; 周彦等, 2007; 王炜等, 2011; 李开开等, 2018)。此外, 鲕粒分布范围广泛, 各地质时期均有发育, 被认为是古气候和古海洋环境的重要指示器, 多形成在温暖动荡的浅水环境(Opdyke and Wilkinson, 1990; Heydari and Moore, 1994; Duguid et al., 2010; Li et al., 2013, 2015, 2017; 李飞等, 2015; Diaz and Eberli, 2019), 如大巴哈马浅滩(Harris et al., 2019)。但是, 并非所有温暖的动荡水体中都发育鲕粒, 这也使得地质学家对此类特殊颗粒的成因机制产生了极大的兴趣。在300多年的研究历史中, 针对这些广泛发育的碳酸盐岩颗粒, 地质学者开展了大量研究工作, 并在此基础上提出了各种成因机制和模型(Davies et al., 1978; Ferguson et al., 1978; Duguid et al., 2010; Diaz et al., 2015, 2017; Mariotti et al., 2018)。但时至今日, 鲕粒的成因机制和形成过程仍存在较大的争议, 主要集中在非生物成因和生物成因方面(Duguid et al., 2010; 梅冥相, 2012; Diaz et al., 2014, 2015, 2017; Tan et al., 2017; Trower et al., 2017; 周瑶琪等, 2017; Mariotti et al., 2018; 宋文天和刘建波, 2020)。
一直以来, 鲕粒的物理— 化学成因机制被很多学者所接受, 并认为鲕粒的形成需要满足一定的条件: 存在合适的核心; 较浅的温暖水域; 碳酸盐饱和或过饱和; 动荡的水体, 有利于CO2的排出(Davies et al., 1978; Sumner and Grotzinger, 1993; Duguid et al., 2010; Rankey and Reeder, 2012; Diaz and Eberli, 2019)。Duguid等(2010)在研究现代巴哈马鲕粒时, 提出了一个新的鲕粒物理— 化学成因模型(非生物成因), 认为鲕粒的形成过程与微生物活动没有直接的关系(图 1-a)。
 | 图 1 现代海相鲕粒成因机制模型简图(据Duguid等, 2010; Diaz and Eberli, 2019, 有修改) a— 鲕粒的物理— 化学成因模型; b— Diaz等(2015, 2017)提出的鲕粒生物成因说模型示意图Fig.1 Cartoon depicting the growth models of modern marine ooids (modified from Duguid et al., 2010; Diaz and Eberli, 2019) |
他们将鲕粒的形成过程划分为2个阶段: 活跃阶段和静止阶段。在活跃阶段, 具有高Mg/Ca值的非晶质碳酸钙(ACC: Amorphous calcium carbonate)在鲕粒最外圈皮层表面形成; 在静止阶段, 这些非晶质碳酸钙重结晶成针状文石, 进而形成新的皮层。但是, Duguid等(2010)并未对非晶质碳酸钙(ACC)的形成过程和机制进行阐述和解释。
近年来, 地质学家在鲕粒生物成因说的相关研究上也取得了较大的突破: Brehm等(2006)在实验室内培养微生物群落, 人工合成了鲕粒; Li等(2017)根据鲕粒皮层中的稀土元素特点证实鲕粒的形成过程有微生物的参与; Diaz等(2015, 2017)提出鲕粒在形成过程中最外层的ACC是微生物作用的结果(图 1-b), 并明确指出鲕粒是不同于叠层石和凝块石的一类特殊微生物岩。
虽然关于鲕粒的成因机制仍存在一定的争议, 但这并不影响其成为有效的古环境和古气候指示器。石炭纪是晚古生代冰期的主要阶段, 全球古气候和古环境发生了重大变革, 同时也是鲕粒发育的重要时期(Opdyke and Wilkinson, 1990; Isbell et al., 2003; Fielding et al., 2008; Montañ ez and Poulsen, 2013)。出露在广西都结地区石炭系都安组上部的“ 都结藻灰岩” 被认为是密西西比亚纪晚期冰川作用的产物, 是一套浅水碳酸盐沉积物, 含有大量微生物席、核形石和鲕粒等特殊沉积(邝国敦等, 1999; 苗卓伟等, 2016; Huang et al., 2020)。文中根据前人的研究理论和成果, 对“ 都结藻灰岩” 中发育的各类鲕粒开展了系统的室内外研究, 并结合其显微组构和沉积环境分析各类鲕粒的形成过程及其对晚古生代冰期的指示意义。
研究区位于广西隆安县都结乡, 大地构造位于“ 滇黔桂盆地” (赵自强和丁启秀, 1996), 又称“ 南盘江盆地” (Enos et al., 1998), 范围大致包括弥勒— 师宗— 普安断裂带东南、 紫云— 罗甸— 南丹— 都安断裂带西南和宁明— 南宁断裂带以北的地区。 晚古生代, 持续拉张作用使盆地内形成了一系列台、 盆相间的古地理格局(焦大庆等, 2003)(图 2)。
 | 图 2 中国南方早石炭世岩相古地理(据焦大庆等, 2003; 有修改)Fig.2 Palaeogeography of the early Carboniferous in South China(modified from Jiao et al., 2003) |
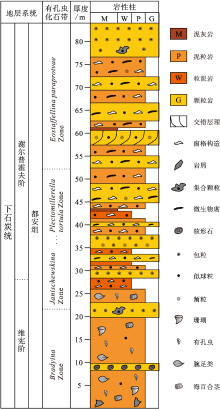
都结地区石炭系保存完好, 出露连续, 属桂西南沉积小区, 自下往上依次发育隆安组、都安组、大埔组、黄龙组和马平组(邝国敦等, 1999)。都安组主要为灰色、浅灰色厚层灰岩、厚层生物屑灰岩、有孔虫灰岩、藻灰岩和介壳灰岩等, 常夹白云质灰岩或白云岩; 生物化石丰富, 主要为浅水底栖生物, 如珊瑚、腕足类、䗴、有孔虫、海百合和藻类等。都安组上部发育一套富含微生物碳酸盐岩的远岸孤立台地浅滩相— 环潮坪相沉积, 分布范围很广, 因在都结地区厚度最大, 也被前人称为“ 都结藻灰岩” (邝国敦等, 1999); 生物化石以藻类为主, 可见少量底栖动物化石, 如珊瑚类的Kizilia、Carcinophyllum、Palaeosmilia等, 非䗴有孔虫Plectogyra sp.、Palaeotextularia sp.、Glomospira sp.、Cribrospira sp.和Bradyina sp.等, 䗴类化石Eostaffella ikensis、E. paraprotvae和Eostaffella sp.等(邝国敦等, 1999)。古生物学特征指示“ 都结藻灰岩” 形成于维宪期末— 谢尔普霍夫期(表 1, 图 3)。
| 表 1 广西隆安地区下石炭统地层格架(据邝国敦等, 1999; 王向东等, 2019; 有修改) Table 1 Stratigraphic framework of the lower Carboniferous in Long’ an area, Guangxi(modified from Kuang et al., 1999; Wang et al., 2019) |
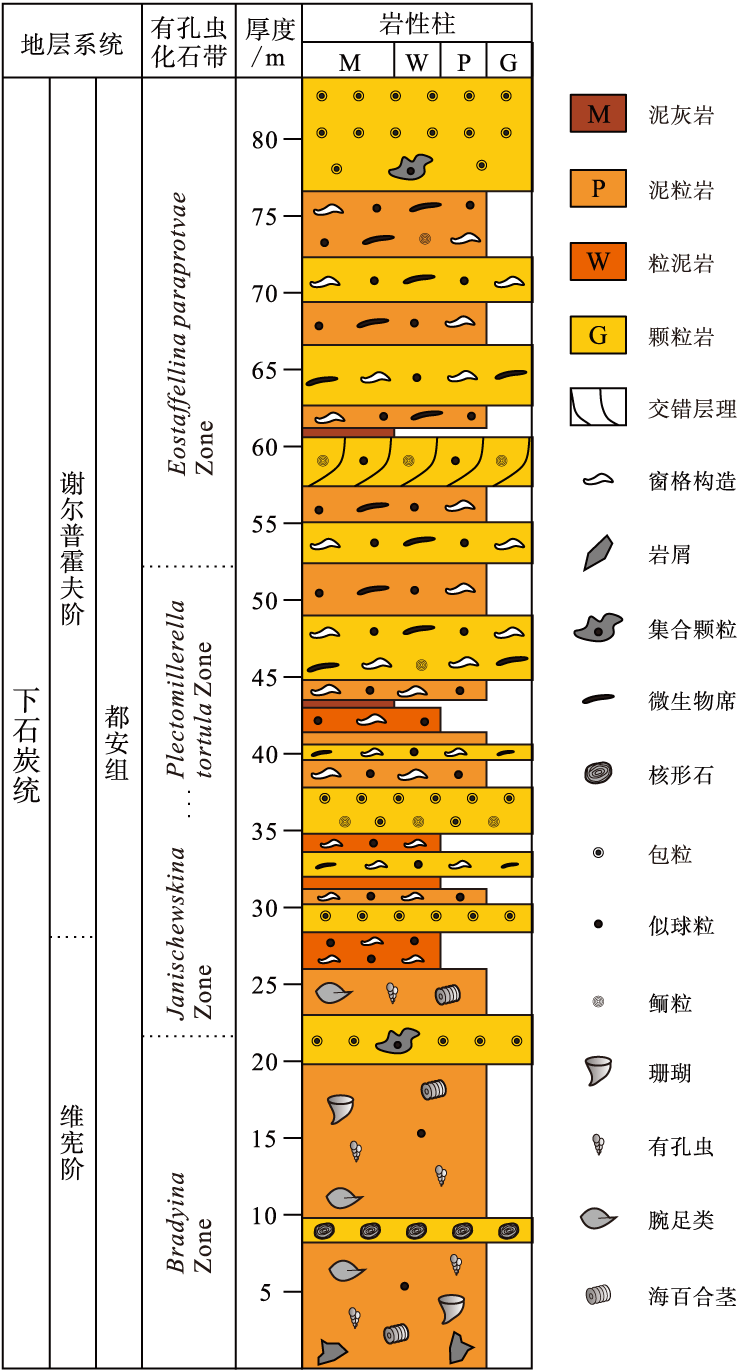 | 图 3 广西隆安地区都结剖面下石炭统都安组岩性柱状图Fig.3 Lithologic column of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
都结地区下石炭统中未见大量陆源碎屑物, 一般不含砂岩夹层, 是远离古陆的远岸台地和孤立台地沉积(邝国敦等, 1999)。邝国敦等(1999)认为发育在都安组顶部的“ 都结藻灰岩” 形成于远岸孤立碳酸盐岩台地浅滩环境, 局部为极浅水的潮坪环境。苗卓伟等(2016)在对“ 都结藻灰岩” 内的微生物岩研究中, 识别出8种微相类型, 包括含亮晶胶结物的泥晶包壳生屑灰岩、含腕足似球粒灰岩、无纹层似球粒颗粒灰岩、团块灰岩、细密纹层状粘结岩、内碎屑鲕粒灰岩、鲕粒灰岩和白云质鲕粒灰岩, 并认为都结剖面微生物岩的沉积环境是在台地边缘浅滩相、开阔台地相和局限台地相3种沉积相带之间往复变化的(苗卓伟等, 2016)。
在此基础上, Huang等(2020)对都结地区都安组上部地层开展了详细的岩相学和同位素地球化学分析, 结果表明“ 都结藻灰岩” 可能与维宪期末— 谢尔普霍夫期开始的冰川作用有关, 是该时期海平面下降的产物。“ 都结藻灰岩” 可划分为3个岩相组合: 底部以厚层状的含岩屑生物碎屑泥粒岩和生物碎屑粒泥— 泥粒岩为主(图 4-a, 4-b), 代表了正常浪基面以下中等水动力条件的潮下带环境; 上覆团块泥粒— 颗粒岩(图 4-c, 4-d)、似球粒— 鲕粒泥粒岩(图 5-a, 5-b, 5-c)、内碎屑鲕粒灰岩(图 5-b, 5-d)和核形石泥粒— 颗粒岩, 是一套正常浪基面之上的浅滩沉积物, 形成于较强水动力条件的浅水潮下带; 富含窗格状构造的潮间带沉积物是剖面上最常见的岩石类型(图 6), 中间夹薄层状潮上带沉积层(图 7), 指示潮坪环境(Huang et al., 2020)。剖面上可见多个由浅滩沉积与潮坪沉积组成的旋回, 此类沉积旋回在华南其他地区、欧洲和北美等地均有发育, 为该时期海平面频繁变化的产物(Veevers and Powell, 1987; Dvorjanin et al., 1996; Wang et al., 2013; Huang et al., 2020)。
 | 图 4 广西隆安地区都结剖面下石炭统都安组典型潮下带微相类型 a— 生物碎屑粒泥— 泥粒岩的野外照片; b— 生物碎屑粒泥— 泥粒岩的显微镜下照片(单偏光); c— 团块泥粒— 颗粒岩的野外照片; d— 团块泥粒— 颗粒岩的显微镜下照片(单偏光); 生物壳体(S), 海百合(C), 有孔虫(F), 泥晶套(ME)Fig.4 Typical subtidal microfacies types of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
 | 图 5 广西隆安地区都结剖面下石炭统都安组高能鲕粒沙坝沉积物 a— 潮间带沉积物向潮下带沉积物过渡的野外照片; b— 发育交错层理的似球粒— 鲕粒泥粒岩(下部)和内碎屑鲕粒灰岩(上部)的光面照片; c— 似球粒— 鲕粒泥粒岩的显微镜下照片(单偏光), 形成于受潮汐作用的沙坝环境; d— 内碎屑鲕粒灰岩的显微镜下照片(单偏光), 形成于相对高能的鲕粒沙坝环境, 下部的似球粒— 鲕粒泥粒岩和潮间带沉积物被波浪或风暴打碎后形成碎屑; 鲕粒(O), 似球粒(P), 岩屑(L)Fig.5 Sediments of ooid sandbar with high energy of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
 | 图 6 广西隆安地区都结剖面下石炭统都安组典型潮间带微相类型 a— 潮间带沉积物的光面照片; b— 纹层状似球粒泥粒岩的显微镜下照片(单偏光); c— 窗格孔藻粘结岩的显微镜下照片(单偏光); d— 内碎屑— 似球粒泥粒岩的显微镜下照片(单偏光); 岩屑(L), 似球粒(P), 窗格状构造(FS)Fig.6 Typical intertidal microfacies types of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
 | 图 7 广西隆安地区都结剖面下石炭统都安组潮上带水平层状叠层石 a— 潮上带水平层状叠层石的野外照片; b— 水平层状叠层石的光面照片, 发育微型帐篷状构造(红色箭头)Fig.7 Stromatolites from supratidal zone of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
综上可知, 都结地区都安组顶部的“ 都结藻灰岩” 形成于远岸浅水孤立台地环境, 主要包括正常浪基面之上的极浅水潮下带(浅滩环境)和潮坪环境(潮间带和潮上带环境), 是维宪期末— 谢尔普霍夫期海平面下降的产物。
“ 都结藻灰岩” 中鲕粒十分发育, 类型丰富, 主要分布在浅滩和潮坪沉积物中。通过对其显微组构的观察统计, 根据鲕粒纹层特征, 将研究区鲕粒划分为5种主要类型(表 2): 放射状纹层鲕粒(O1)、规则同心放射状纹层鲕粒(O2)、不规则同心放射状纹层鲕粒(O3)、泥晶鲕粒(O4-A和O4-B)和复合鲕粒(O5)。
| 表 2 广西隆安地区都结剖面下石炭统都安组鲕粒类型及其特征 Table 2 Ooids and their characteristics of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
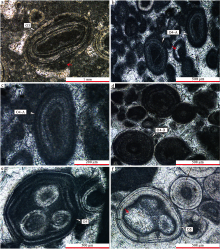
放射状纹层鲕粒(O1)在剖面内整体含量不高, 但在潮间带和潮下带环境都有发育, 可与规则同心放射状纹层鲕粒和泥晶鲕粒伴生。在潮间带沉积物中, 放射状纹层鲕粒零散分布, 偶见局部集中分布现象(图 8-a)。其粒径较小, 多为0.2~0.8 mm, 呈规则的圆形— 椭圆形(放射皮层较厚), 也可见受核心形态控制的其他外形(放射皮层较薄)(图 8-a)。纹层保存完好, 呈放射状, 厚度相对均一, 纹层内可见暗色泥晶物质残余。核心类型单一, 以泥晶方解石为主, 其他类型核心少见。在具交错层理的浅水潮下带似球粒— 鲕粒灰岩中, 放射状纹层鲕粒和规则同心放射状纹层鲕粒是主要的鲕粒类型(图 8-b), 粒径多在0.2~0.5 mm之间, 且部分泥晶化严重, 纹层结构模糊不易辨认。
 | 图 8 广西隆安地区都结剖面下石炭统放射状纹层鲕粒和规则同心放射状纹层鲕粒的显微特征(单偏光) a— 潮间带沉积物中厚皮层圆形和薄皮层不规则放射状纹层鲕粒(O1); b— 具交错层理的潮汐沙坝内发育的放射状纹层鲕粒(O1)和规则同心放射状纹层鲕粒(O2); c— 内碎屑鲕粒灰岩内的规则同心放射状纹层鲕粒(O2), 皮层内含多个明显的泥晶纹层(红色箭头); d— 鲕粒泥晶核心溶蚀后被亮晶方解石充填; e— 规则同心放射状纹层鲕粒, 受成岩作用影响部分同心纹层消失, 呈现放射状(红色箭头); f— 图e红色方框内放大照片, 皮层内同心纹层数大于10; P为似球粒, D为白云石Fig.8 Thin-section photographs(plane-polarized light)of radial ooids and regular concentric-radial ooids of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
规则同心放射状纹层鲕粒(O2)是剖面上丰度最高的鲕粒类型, 发育环境包括浅水潮下带和潮间带, 在所有含鲕层位都有发现。此类鲕粒多呈圆形— 椭圆形, 粒径变化较大, 0.2~1.5 mm不等。皮层较厚, 放射状亮层被极薄的泥晶方解石暗层隔开(图 8-c, 8-d), 其中亮层连续性较好, 同一层厚度相对均匀, 单层厚10~50 μ m, 部分可达80 μ m; 而暗层较薄, 单层厚5~10 μ m, 少数可达30 μ m, 同一层的连续性较亮层差。核心以泥晶方解石为主, 未见生物碎屑核心, 部分鲕粒核心溶蚀消失, 被亮晶或微亮晶方解石充填(图 8-d)。不同环境中的规则同心放射状纹层鲕粒的粒径和皮层圈数变化较大: 在具交错层理的浅水潮下带似球粒— 鲕粒灰岩中, 粒径最小, 多在0.2~0.5 mm之间, 同心纹层数也最少, 一般小于5圈, 多为1~3圈(图 8-b); 在含内碎屑鲕粒灰岩中, 此类鲕粒粒径多在0.2~0.8 mm之间, 同心纹层数增多, 3~8圈不等(图 8-c, 8-d); 在潮坪沉积环境中, 规则同心放射状纹层鲕粒零散分布在富含泥晶物质的粒泥岩中, 可与不规则同心放射状纹层鲕粒伴生, 窗格状构造发育, 粒径在0.8~1.5 mm之间, 同心纹层数最多, 可大于10圈(图 8-e, 8-f)。
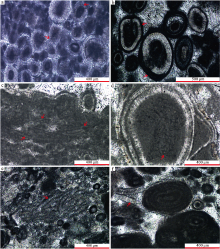
研究区不规则同心放射状纹层鲕粒(O3)含量相对较少, 发育环境也较为局限, 主要出现在富含泥晶质的潮坪相粒泥岩中, 窗格状构造发育, 与似球粒和少量规则同心放射状纹层鲕粒伴生。粒径相对较大, 多在0.5~2 mm之间(图 9-a), 也可见大于2 mm的个体。
 | 图 9 广西隆安地区都结剖面下石炭统都安组鲕粒的显微特征(单偏光) a— 不规则同心放射状纹层鲕粒(O3), 见不规则的泥晶纹层(红色箭头); b— 泥晶鲕粒(O4-A), 富有机质核心溶蚀后形成的偏心鲕粒(红色箭头); c— 图b右侧泥晶鲕粒(O4-A)的放大照片, 纹层结构保存完好; d— 泥晶化鲕粒, 皮层结构消失或模糊; e— 复合鲕粒 (O5), 核心为鲕粒形成的葡萄石; f— 复合鲕粒, 核心被亮晶方解石充填(红色箭头)Fig.9 Thin-section photographs(plane-polarized light)of ooids of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
鲕粒核心主要为泥晶方解石, 未见其他类型核心。鲕粒的外形受核心的形态影响比较大。皮层由亮暗相间的纹层组成, 具有较好的韵律性, 亮层厚度相对较小, 为5~20 μ m, 且亮层越厚, 连续性越好; 暗层为富含有机质的泥晶方解石, 层厚变化较大, 为5~50 μ m, 部分位置可达100~200 μ m。
都结地区泥晶鲕粒(O4)可划分为2个亚类: 泥晶鲕粒O4-A(图 9-b, 9-c)和泥晶鲕粒O4-B(图 9-d)。
泥晶鲕粒O4-A主要分布在潮坪相含鲕似球粒泥粒岩内, 呈椭圆形, 粒径较小, 多为0.1~0.5 mm。核心以泥晶方解石为主, 未见生物碎屑核心, 且富含有机质的泥晶方解石核心常被亮晶方解石取代, 形成偏心鲕(图 9-b)。鲕粒的外形受核心形态影响较大。该类鲕粒皮层由亮暗相间的泥晶纹层组成, 纹层保存完好(图 9-c), 亮层由微晶— 微亮晶方解石组成, 单层厚度5~10 μ m, 最厚达15 μ m; 暗层由泥晶方解石组成, 厚度多小于10 μ m。多数泥晶鲕粒和同层的其他颗粒(如正常鲕粒)均被泥晶方解石包壳。
泥晶鲕粒O4-B零散分布于浅水潮下带浅滩相沉积物中, 与各类似球粒、团块和生物碎屑伴生, 呈圆形— 椭圆形, 粒径0.2~0.8 mm, 皮层结构模糊, 难以辨认(图 9-d)。
复合鲕粒(O5)在剖面内也十分常见, 主要发育在潮坪相沉积物中, 在浅水潮下带浅滩沉积物中零星可见, 粒径变化较大, 0.5~3 mm不等, 主要受核心大小和形态的影响。核心多为泥晶方解石胶结的葡萄石(图 9-e), 也可见富含有机质的核心溶蚀后被亮晶方解石取代现象(图 9-f)。复合鲕粒的皮层相对较薄, 由保存完好的亮暗相间纹层组成, 在不同鲕粒中亮暗层厚度、连续性和保存状态差别较大, 亮层一般厚5~30 μ m, 暗层为泥晶方解石, 厚5~60 μ m。
目前, 虽然鲕粒的成因机制仍存在争议, 但越来越多的人开始接受鲕粒的生物成因说(Brehm et al., 2006; 梅冥相, 2012; Li et al., 2013, 2015, 2017; Diaz et al., 2014, 2015, 2017; 邢延路和冯李强, 2015; 倪胜利, 2017; Tan et al., 2017; Mariotti et al., 2018; 陈百兵等, 2019; 郭芪恒等, 2019; 蒋裕强等, 2019)。Diaz等(2017)对现代鲕粒的研究证实, 微生物在鲕粒的形成过程中扮演着重要的角色。鲕粒的形成过程可以划分为2个主要阶段: 静止阶段和活跃阶段(Diaz et al., 2017)。在静止阶段, 微生物所分泌的胞外聚合物(EPS: Extracellular polymeric substance)在鲕粒的最外层聚集生长。随后, 在活跃阶段, EPS中的各类酸性官能团(如: COO-、
1)放射状纹层鲕粒。鲕粒的放射状纹层是晶体沿垂直或近垂直于核心表面或纹层的方向排列所形成的, 多形成于水动力条件较弱的浅海环境(Davies and Martin, 1976; Davies et al., 1978; Land et al., 1979), 也可出现在高盐度的水体环境(Friedman et al., 1973; Halley, 1977; Flü gel, 2010)。在研究区, 放射状纹层鲕粒主要出现在潮间带和浅水潮下带沙坝沉积物中(图 10)。具交错层理的似球粒— 鲕粒灰岩中颗粒具有很好的分选性和磨圆度, 代表了具有较高水动力条件的潮汐沙坝环境(图 5-b, 5-c; 图 8-b)。周瑶琪等(2017)在模拟海相鲕粒形成过程中观察到: 在静水条件下, 碳酸钙发生沉淀并形成垂直鲕粒表面生长的晶体; 而在水动力条件增强时, 晶体方向发生变化, 不能形成稳定的放射状圈层。因此, 强水动力条件的沙坝环境可能不利于放射状纹层的形成(Davies and Martin, 1976; 周瑶琪等, 2017)。其内部发育的放射状纹层鲕粒个体较小, 与大量规则同心放射状纹层鲕粒伴生, 部分具同心纹层残余结构(图 11-a), 推测其原始结构很可能是同心放射状纹层, 经后期磨蚀、微生物侵蚀或成岩等作用而不易辨认, 部分表现出放射状纹层结构。潮间带沉积物中放射状纹层鲕粒零散分布, 放射状结构保存相对完好(图 8-a), 可能代表了原始纹层结构, 形成于潮间带低洼处。相对平坦的潮间带常发育若干洼地, 为鲕粒放射纹层的形成创造了极浅的安静水体, 而强烈的蒸发作用和间歇性的海水补给不仅有利于维持CaCO3过饱和状态, 也成为CaCO3的主要来源(余素玉等, 1987)。
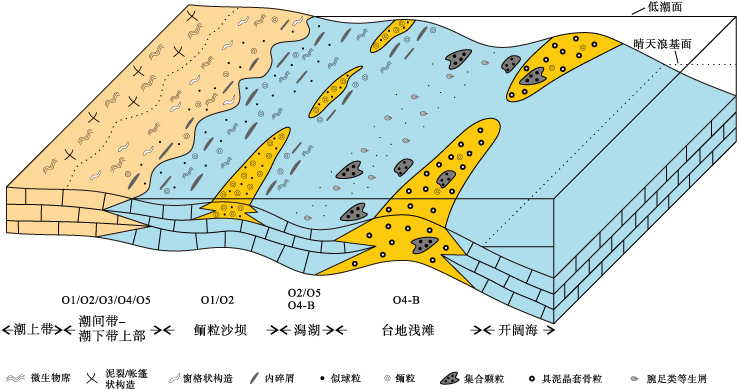 | 图 10 广西隆安地区下石炭统都安组沉积环境及鲕粒分布Fig.10 Depositional environment and distribution of ooids of the lower Carboniferous Du’ an Formation in Long’ an area, Guangxi |
 | 图 11 广西隆安地区都结剖面下石炭统都安组鲕粒形成中微生物作用的显微镜下照片(单偏光) a— 似球粒— 鲕粒灰岩中放射状纹层鲕粒皮层内残余的同心纹层结构(红色箭头); b— 鲕粒表面形成的致密微生物泥晶套(红色箭头); c— 钙质微生物Ortonella(红色箭头)和微生物席内疑似鲕粒雏形(白色箭头); d— 鲕粒核心内的疑似微生物菌丝(红色箭头); e、f— 与鲕粒伴生的钙质藻类(红色箭头)Fig.11 Thin-section photographs (plane-polarized light) showing microbial activities during formation of ooids of the lower Carboniferous Du’ an Formation in Dujie section of Long’ an area, Guangxi |
2)规则同心放射状纹层鲕粒。规则同心放射状纹层鲕粒是“ 都结藻灰岩” 中最常见的一类鲕粒类型, 皮层由放射状亮晶纹层与同心泥晶纹层组成, 构成亮暗纹层组。
从潮下带浅滩环境到潮坪环境, 规则同心放射状纹层鲕粒含量逐渐降低, 粒径却逐渐增加, 表现出与沉积环境的显著联系。在水动力较强的潮下带浅滩环境, 交错层理发育, 鲕粒含量最高, 但粒径最小, 同心纹层数也最少(图 8-b; 图 11-a)。这可能是因为在持续高能的浅滩, 虽然温暖动荡的水体环境有利于鲕粒的生长, 但颗粒滚动时间增加, 加剧了鲕粒等颗粒物质之间的碰撞和磨蚀作用, 不易形成粒径较大的鲕粒(齐永安等, 2014; Mariotti et al., 2018)。而在潮间带和潮下带下部环境, 间断性的水体搅动不仅为鲕粒的形成创造了有利条件, 也减弱了磨蚀作用, 致使粒径有所增加。鲕粒皮层中富含有机质的泥晶纹层被认为可能形成于鲕粒中碳酸盐晶体生长停滞时期, 是微生物在其表面生长形成胞外分泌物, 并诱导方解石沉淀的结果(Tan et al., 2017)。放射状亮晶纹层被多期次的泥晶薄层隔开, 指示了此类鲕粒生长过程包括了多个较显著的“ 静止— 活跃” 生长周期。
3)不规则同心放射状纹层鲕粒和复合鲕粒。不规则同心放射状纹层鲕粒和复合鲕粒粒径最大, 分布范围相对局限, 主要发育在具有中等水动力条件的水体环境。其皮层中的泥晶纹层较厚, 形态不规则, 常出现加厚现象(图 9-a, 9-e), 这可能是多种因素作用的结果: 此类鲕粒在形成过程中可能经历了较长的停滞期, 在微生物作用下形成了较厚的泥晶纹层, 在此过程中颗粒滚动有限, 使得微生物或胞外分泌物在鲕粒表面分布不均匀; 当水动力增强, 鲕粒在随后的活跃阶段没有经历充分的碰撞和磨蚀, 并被搬运至相对低能环境沉积、埋藏。此外, 复合鲕粒的核心多为含鲕粒的葡萄石(图 9-e), 可能是鲕粒被搬运至相对静水环境后, 微生物对其进行侵蚀、钻孔、融合和粘结的结果(Mariotti et al., 2018)。
4)泥晶鲕粒。 研究区的泥晶鲕粒主要分布在潮间带环境和潮下带浅滩环境。 研究表明泥晶鲕粒主要有鲕粒泥晶化作用、 低沉积速率环境(如外斜坡环境)中微晶方解石的生长和微生物作用3种成因(Flü gel, 2010; Woods, 2013)。 研究区浅滩相发育的泥晶鲕粒(O4-B)内部结构模糊或消失(图 9-d), 与团块和磨圆较好的生物碎屑伴生, 此类泥晶鲕粒可能是正常鲕粒泥晶化的结果。 而在部分潮间带沉积物中分布的泥晶鲕粒(O4-A), 其内部结构完整, 同心纹层结构清晰(图 9-b, 9-c), 区别于泥晶化鲕粒。 同时, 潮坪环境的沉积速率相对较快, 因此也排除了研究区泥晶鲕粒在低沉积速率环境下的成因机制。 通过比较发现此类泥晶鲕粒结构特点与Woods(2013)识别出的微生物泥晶鲕粒一致, 可能是微生物作用的产物。
通过对各类鲕粒分布情况和形成过程的分析, 笔者认为研究区的鲕粒不仅受控于古海洋环境, 同时微生物活动可能在其形成过程中也扮演了重要的角色。
1)形成微生物泥晶鲕粒O4-A。依据上述分析中泥晶鲕粒O4-A皮层完好的纹层结构及其发育环境, 并类比Woods(2013)提到的微生物泥晶鲕粒, 推测O4-A可能为一种由微生物作用形成的鲕粒。除以上证据外, 与O4-A伴生的其他类型鲕粒和颗粒表面常被1层致密微晶方解石包裹形成泥晶套(图 11-b), 泥晶套厚度不均一, 与内部鲕粒界线清晰, Woods(2013)认为此类泥晶套属于“ 建造型” 泥晶套, 是微生物在颗粒表面生长、诱导或捕获泥晶方解石形成的, 与核形石纹层相似, 区别于泥晶化作用形成的“ 破坏性” 泥晶套(泥晶套与颗粒之间界线模糊)。同时, 鲕粒常与钙质微生物伴生, 并在微生物席内可见疑似鲕粒雏形颗粒(图 11-c), 与现代微生物席内的鲕粒类似(Gerdes et al., 1994), 这也说明此类泥晶鲕粒的形成可能与微生物活动有关。
2)提供鲕粒核心。泥晶方解石(如似球粒)是研究区鲕粒最主要的核心类型, 其他核心类型(如生物碎屑)十分少见。在保存完好的泥晶核心内部可见疑似微生物菌丝的纤维状物质(图 11-d), 这说明研究区部分鲕粒的泥晶核心可能是微生物作用的产物(如微生物席碎屑)。
3)诱导CaCO3沉淀。虽然鲕粒的放射状纹层多被认为是碳酸盐晶体自然生长的结果, 但生物因素可能也直接或间接地影响了其形成过程(Davies and Martin, 1976; Davies et al., 1978)。在微生物席中, 微生物通过光合作用可以大量消耗水体中的CO2, 同时释放出OH-, 改变水体微环境, 提高pH值和C
4)鲕粒泥晶纹层的形成。研究区内鲕粒皮层中暗色泥晶纹层十分显著, 尤其是在不规则同心放射状纹层鲕粒和复合鲕粒中最为突出, 此类泥晶纹层可能是鲕粒在静止阶段微生物群落在其表面生长并诱导碳酸盐沉淀或捕获水体中的碎屑物质形成的(Li et al., 2017; Mariotti et al., 2018)。
研究区的各类鲕粒发育在以潮坪沉积物为主的“ 都结藻灰岩” 中, 形成于受潮汐作用影响的浅水孤立台地(邝国敦等, 1999), 发育在都安组上部, 时代为维宪期末— 谢尔普霍夫期, 是维宪期末全球性海平面下降的产物(苗卓伟等, 2016; Huang et al., 2020)。这一时期, 在世界多地古海平面表现出了相似的变化规律, 如: 古纬度较高的阿根廷西部盆地(Gonzá lez, 1990; Limarino et al., 2006; Perez Loinaze et al., 2010); 古纬度较低的美国内华达州东南部的Arrow Canyon Range地区(Bishop et al., 2009)、蒙大拿中部大雪山地区(Ahern and Fielding, 2019)、中国南方(Wang et al., 2013; Huang et al., 2020)和英国多个盆地(Wright and Wagonerstone, 2001; Fielding and Frank, 2015), 以及冈瓦纳大陆的其他地方(Veevers and Powell, 1987; Smith and Read, 2000)。与此同时, 在南美洲巴西多个盆地(Caputo et al., 2008)、阿根廷中西部(Limarino et al., 2006; Perez Loinaze et al., 2010)和中国西藏南部(Garzanti and Sciunnach, 1997)等中高纬度地区地层中大量发育冰成岩, 并伴随着δ 13C显著的正向漂移(Grossman et al. 2008; Qie et al., 2011; Huang et al., 2020; Yang et al., 2020)。因此, 多数学者也认为本次全球性海平面下降可能是该时期冰川开始广泛发育的结果(Bishop et al., 2009; Qie et al., 2011; Wang et al., 2013; Chen et al., 2016; Ahern and Fielding, 2019; Huang et al., 2020)。维宪期末大规模的海退事件, 迫使研究区由相对深水的潮下带环境转变为浅水的潮下带浅滩环境和潮坪环境, 水动力条件显著增强, 为鲕粒的形成提供了动荡水体。石炭纪华南地区位于古低纬度地区, 属热带— 亚热带气候(Wang et al., 2013)。维宪期末— 谢尔普霍夫期, 虽然中高纬度地区冰川开始大量发育, 但对低纬度地区浅层海水温度影响不大, 主要表现为频繁的海平面变化。研究区特殊的古地理位置使其在冰川作用的海退期为“ 都结藻灰岩” 内各类鲕粒的形成提供温暖、动荡和CaCO3过饱和的浅水环境。
1)广西都结地区下石炭统都安组上部“ 都结藻灰岩” 中鲕粒十分发育, 类型丰富。根据显微组构、核心及纹层特征, 共识别出5种主要类型: 放射状纹层鲕粒、规则同心放射状纹层鲕粒、不规则同心放射状纹层鲕粒、泥晶鲕粒和复合鲕粒。
2)各类鲕粒的显微组构和分布情况主要受水动力条件影响。在动荡的浅滩环境, 鲕粒悬浮、滚动和磨圆较充分, 常形成小粒径的规则同心放射状纹层鲕粒, 而在部分潮坪或局限环境则发育放射状纹层鲕粒、不规则同心放射状纹层鲕粒和泥晶鲕粒。
3)在广西都结地区下石炭统都安组上部, 钙质微生物和微生物席十分发育, 常与各类鲕粒伴生。结合现代鲕粒研究成果, 推测微生物活动在研究区鲕粒形成过程中可能起到了一定的作用, 如提供泥晶核心、形成同心泥晶纹层、催化或诱导CaCO3沉淀等。
4)维宪期末— 谢尔普霍夫期, 冰川作用的开始导致了全球海平面的大幅下降和频繁波动, 可能是促使“ 都结藻灰岩” 形成的主要原因。而研究区位于低纬度地区, 在一定程度上为各类鲕粒的形成提供了有利的水体环境。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|