




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国遗迹学研究十年进展
[胡斌1, 3  , 齐永安
, 齐永安1, 2, 3 , 宋慧波1, 3 , 牛永斌1, 3 , 张立军1, 3 , 郑伟1, 3 , 王长征1, 3 ]
, 齐永安, 牛永斌|
|

通讯作者简介 宋慧波,女,1979年生,教授,博士生导师,主要从事古生物地层学、遗迹学和沉积学教学与研究工作。E-mail: songhuibo2005@hpu.edu.cn。
第一作者简介 胡斌,男,1952年生,教授,博士生导师,主要从事古生物地层学、遗迹学和沉积学的教学和研究工作。E-mail: hub@hpu.edu.cn。
近十年来,中国遗迹学的研究取得许多新进展和新认识,重要成果概述如下: (1)在显生宙海相和过渡相沉积中新识别出84个遗迹属和211个遗迹种,新建28种遗迹组合及26种遗迹组构;在中生代和新生代陆相冲积扇、河流与湖泊沉积中识别出44个遗迹属和107个遗迹种,新建22种遗迹组合及10种遗迹组构;(2)论述了寒武纪底质革命对早期底栖生物的进化和生态效应的影响;(3)提出了3种古氧相(常氧相、贫氧相和厌氧相)的遗迹化石响应关系及其形成的沉积背景;(4)在华北下二叠统太原组灰岩中发现了大量成因与微生物相关的碳酸盐岩微形体,在动藻迹( Zoophycos)潜穴充填物中识别出球状、杆状、簇状、网状、瓶状和似脑球状6种微形体,描述了由造迹者与共生微生物一起进行一系列生命活动行为所形成的复杂精美的生物成因构造;(5)揭示了复杂遗迹化石动藻迹( Zoophycos)从浅阶层迁移至深阶层与从浅海迁移至半深海—深海同步的演化规律;(6)提出了滨—浅海和湖泊环境中风暴与浊流事件沉积序列中以及P/Tr界线附近遗迹化石的组成与分布特征,并论证了P-Tr事件前后遗迹化石及其造迹生物生态系统的演变规律,为晚二叠世末生物大灭绝后的生态系统复苏过程提供了可靠的遗迹学信息;(7)在华东、华北和西部地区多处新发现侏罗纪和白垩纪大量兽脚类、蜥脚类和鸟脚类恐龙足迹群;(8)在华北地台中元古界和上二叠统—下三叠统碎屑岩中识别出多种微生物成因沉积构造;(9)详细分析了碎屑岩和碳酸盐岩储集层中的生物扰动作用和遗迹组构及其对储集层物性的影响,建立了生物扰动变化三维地质模型,并提出了生物扰动型储集层的表征与研究方法。上述研究成果极大地促进了中国在遗迹分类学、遗迹相、遗迹组构、地微生物以及遗迹学在事件沉积和油气储层应用研究等方面的发展和进步。
, Qi Yong-An, Niu Yong-BinAbout the corresponding author Song Hui-Bo,born in 1979,is a professor and Ph.D. supervisor of Henan Polytechnic University. She is mainly engaged in teaching and researches of paleontology and stratigraphy,ichnology and sedimentology. E-mail: songhuibo2005@hpu.edu.cn.
About the first author Hu Bin,born in 1952,is a professor and Ph.D. supervisor of Henan Polytechnic University. He is mainly engaged in teaching and researches of paleontology and stratigraphy,ichnology and sedimentology. E-mail: hub@hpu.edu.cn.
Many new advances and understandings have been made in the study of Chinese ichnology in recent ten years,and key results are outlined below: (1)84 ichnogenera and 211 ichnospecies have been identified and 28 types of ichnoassemblages and 26 types of ichnofabrics have been established in the Phanerozoic marine and transitional deposits;44 ichnogenera and 107 ichnospecies have been identified and 22 types of ichnoassemblages and 10 kinds of ichnofabrics have been established in the Mesozoic and Cenozoic terrestrial(alluvial fan,fluvial and lacustrine)deposits;(2)The impact of the Cambrian Substrate Revolution on the evolution of early benthic organisms and ecological effects was discussed;(3)The response relationship of 3 kinds of palaeo-oxygenation(aerobic,dysaerobic and anaerobic)facies to ichnofossils and their deposition backgrounds was proposed;(4)Many carbonate micro-shapes(single mineral and aggregates)related to microbes were found in limestones of the Lower Permian Taiyuan Formation of the northern China,and six kinds of carbonate micro-shapes were distinguished by observing Zoophycos burrow fillings,which are spheroid-like,rhabditiform,areatus,net-like,vase-like and cerebrum-like. Meanwhile,it was described that many complex and delicate biogenetic structures formed by a series of life activities of trace-makers with symbiotic micro-organisms;(5)The sedimentary setting evolution of a complex ichnofossil Zoophycos was revealed, i.e.,the bathymetrical shift of Zoophycos from the littoral-neritic to bathyal-abyssal environments synchronized with the tiering shift from shallow to deep;(6)It was suggested that the composition and distribution characteristics of ichnofossils in a sedimentary sequence of storm and turbidity currents in the littoral-neritic and lacustrine environments and nearby P-Tr boundary,and the ecological system evolution law of ichnofossils and their trac-makers before and after the P-Tr event was demonstrated,which provides a reliable ichnologic information for the ecosystem recovery process after the P/Tr great extinction;(7)A large amount of theropod,sauropod and ornithopod dinosaur tracks were found in the Jurassic and Cretaceous of many places in the eastern,northern and western China;(8)Many types of microbially induced sedimentary structures were distinguished in the clastic deposits of the Mesoproterozoic and Upper Permian-Lower Triassic in the North China Platform;(9)The bioturbation and ichnofabrics in clastic and carbonate rock reservoirs and their impact on reservoir physical properties were analyzed in detail,a three-dimensional geological model of bioturbation changes was established,and the manifestation and research methods of bioturbation type reservoirs were put forward. These results have greatly promoted the development and progress of ichnological field including ichnotaxon,ichnofacies,ichnofabrics,geomicrobiology and application in the event deposits and reservoir geology in China.
近十年来, 中国遗迹学者做了大量卓有成效的研究工作, 据国内外期刊相关文章的不完全统计, 在中国中元古代至新生代古近纪海相和过渡相地层中识别出的遗迹化石计有211个遗迹种, 陆相地层中有107个遗迹种。限于篇幅, 文中仅总结了7个方面的主要内容, 分别是: (1)寒武纪遗迹组合、组构与底质革命, (2)奥陶纪— 泥盆纪遗迹组合、组构、时空演化及生态习性, (3)石炭纪— 二叠纪遗迹组合、组构及古氧相与地微生物学, (4)三叠纪海相地层及P/Tr界线附近遗迹化石组成与分布特征, (5)陆相遗迹化石组合与组构, (6)微生物成因沉积构造, (7)遗迹学在事件沉积和油气地质勘探方面的应用研究等。
近十年来, 中国寒武纪的遗迹化石研究主要集中在华北地区, 尤其是在河南省和山西省境内的寒武系中做了较多的遗迹化石调研工作, 豫西洛阳、渑池、登封和鲁山等地寒武系中的遗迹化石研究较详。共识别出遗迹化石19个遗迹属和33个遗迹种, 建立了6类遗迹组构, 在遗迹化石的沉积背景分析以及其与沉积底质的关系方面取得许多新的研究进展和认识。
辛集组为一套风暴作用影响下的浅海相含磷含海绿石碎屑岩沉积, 主要由红色中厚层石英砂岩及极薄层泥岩与砂岩互层组成。遗迹化石主要产出在砂岩、极薄层泥岩与薄层砂岩互层的沉积序列中, 常见2类: 第1类是机会(r-选择)遗迹化石Skolithos, 属Skolithos遗迹相; 第2类是均衡(K-选择)遗迹化石, 包括Gordia, Palaeophycus, Planolites和 Taenidium等, 属Cruziana遗迹相(白万备等, 2018)。Skolithos主要发育在风暴作用衰减期形成的平行层理砂岩中, 它是一种不受沉积相控制、高度局限、低分异度的机会(r-选择)遗迹化石。从产状上看, Skolithos仅孤立分布在层面上, 不与其他遗迹化石共生, 反映出风暴期沉积速率高、环境不稳定、不断迁移的砂质底质不利于大多数造迹生物的殖居, 只有少量的喜沙悬食生物才能够在这种恶劣的环境中掘穴。Cruziana遗迹相主要发育在风暴间歇期形成的极薄层砂岩与泥岩互层层面上。风暴间歇期的环境稳定, 适合造迹生物的生活, 留下的遗迹化石具有均衡(K-选择)遗迹化石特点, 即遗迹分异度高、丰度一般、造迹生物采取高度专业化的行为方式。华北地区辛集组发育的遗迹化石主要以层面觅食迹和进食迹为主, 在同一层面可见多种大小不同的遗迹化石共生, 如Palaeophycus tubularis, Palaeophycus striatus与具有新月形回填纹构造的Taenidium isp.出现在同一层面上, 也可见相互交叉重叠现象, 为不同造迹生物共同扰动留下的构造。
此外, 苗兰云和朱茂炎(2014)在河南省石门沟、阳坡和山西省水峪等地寒武系辛集组近底部层位采集遗迹化石计有8个遗迹属和12个遗迹种。这一遗迹化石组合属非典型的Cruziana遗迹相, 形成于盐度正常的潮下带砂质软底环境。
豫西地区馒头组由灰黄色、灰红色、暗紫色含粉砂钙质泥岩夹含泥微晶灰岩逐渐过渡为粉砂质泥岩、粉砂岩与灰岩互层夹砂岩, 由潮上带泥坪沉积逐渐演变为潮间带混合坪和碎屑岩— 碳酸盐混积台地沉积。该组碎屑岩中的遗迹化石主要发育在潮间带混合坪沉积中, 并以固底控制的遗迹化石为特征(李妲等, 2016)。沉积物表面的抓痕及爬痕类固底遗迹化石主要有Rusophycus, Beaconichnus, Cruziana, Diplichnites, Dimorphichnus, Monomorphichnus 和Qipanshanichnus等。潜穴类固底遗迹化石包括Palaeophycus striatus, Bergaueria hemispherica, Beaconites antarcticus, Treptichnus isp.等。馒头组固底遗迹化石以表面爬痕、抓痕、沉积物表面粘附的潜穴和浅阶层水平潜穴为主, 缺乏深掘的垂直潜穴和强烈生物扰动, 对沉积物起混合作用的生物扰动较少。潜穴类固底遗迹化石的围岩颜色均一无杂色, 潜穴边缘清晰分明, 不发育衬壁或加强边缘, 充填物与围岩界限明显, 并且充填物为上覆的粗粒砂质沉积物, 无变形或坍塌现象, 显示开放式居住觅食潜穴特征。这反映了当时具有一定固结程度且不发育混合层的沉积物特征, 表明沉积物的混合程度较低。
豫西地区朱砂洞组沉积环境为潮上带蒸发坪、潮间坪和局限台地。野外露头观测显示, 生物扰动构造与围岩因差异风化在岩石表面呈斑驳形态, 前者呈灰黑色— 深灰色, 风化后凸出于层面之上, 后者颜色较浅, 呈浅灰色— 灰白色, 风化后凹进层面, 二者界线清晰可辨(齐永安等, 2014a)。单个潜穴光滑, 无衬壁, 直径差别不大, 通常为3~10 mm, 少数可达15 mm, 分枝直径相比主枝略小。依据野外照片, 采用Image J2x软件辅助对图像进行分析, 逐层统计Thalassinoides所占地层百分比, 将其扰动指数划分为BI=2~5。当指数BI=2时, 潜穴直径为3~4 mm, 扰动构造在剖面上呈顺层面的连续条带状, 单位面积内扰动部分占10%~20%; 当指数BI=3时, 潜穴直径为4~6 mm, 扰动构造在剖面上多呈平行于层面的断续条带状, 层面上多呈单个微曲管状或少数发育Y形分枝, 单位面积内扰动部分占31%~55%; 当指数BI=4时, 潜穴直径为5~12 mm, 扰动构造在剖面上多呈方向排列不定的Y或T形分枝, 扰动深度相比中等程度略微增加, 层面上也多呈排列间距不定的Y或T形分枝, 潜穴之间常见交叉叠覆, 单位面积内扰动部分占61%~84%; 当指数BI=5时, 潜穴直径为8~13 mm, 扰动构造在剖面或层面上绝大多数发育Y或T形分枝, 少数呈单个管状, 潜穴之间完全交叉叠覆, 沉积界面改造强烈, 单位面积内扰动区域可达90%以上。Thalassinoides这种交互式巷道潜穴系统代表着深层生物扰动者的行为, 它们能够将沉积物自下向上通过潜穴内溶质运移到层面上, 并通过增加沉积物— 水界面的面积进入沉积物中。造迹生物沿着沉积物— 水界面向下掘穴过程中, 能够使海水与沉积物进行物质交换, 改变沉积物孔隙水的化学组成。
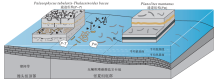
河南省洛阳、登封和渑池等地张夏组中的遗迹化石研究程度较高, 依据沉积学和遗迹学特征, 本组近滨碳酸盐岩中识别出6类遗迹组构, 每一类遗迹组构在垂向上连续叠加, 重复出现, 代表了沉积环境的高频交替循环变化(齐永安等, 2012): ① Planolites montanus 遗迹组构, 分布在张夏组底部浅灰色平板状薄层微晶灰岩与深灰绿色页岩薄互层中和中下部厚5.3 m薄层微晶灰岩与土黄色灰泥岩薄互层中, 由单一的遗迹化石Planolites montanus组成, 且严格分布在灰岩层面上, 形成在平均浪基面之下和平均风暴浪基面之上的潮下低能较深水环境, 但常遭受强风暴流的影响(图 1); ②Palaeophycus tubularis-Thalassinoides bacae遗迹组构, 产于该组中下部灰白色微晶灰岩夹极薄层浅土黄色泥质灰岩中, 遗迹分异度高, 能识别出的遗迹化石较多, 有Palaeophycus tubularis, Thalassinoides bacae, Thalassinoides horizontalis, Skolithos linearis, Arenicolites isp.和Planolites isp.等, 形成在与广海连通性较好并时常受到风暴流影响的无鲕粒碳酸盐岩台地较浅水区(图 1); ③ Planolites montanus-Skolithos linearis遗迹组构, 分布在剖面中部, 形成在深灰色薄层鲕粒灰岩与浅灰色薄层微晶灰岩互层沉积中, 垂向生物扰动主要是由Skolithos引起的, 平面上的生物扰动主要是Planolites引起的, Skolithos也有一定的贡献, 形成于高、低能交替的滨岸浅滩或开阔台地滩间坪或滩前沉积中(图 2); ④ 模糊生物扰动遗迹组构, 总体特征是扰动量大, 生物扰动构造与围岩的颜色和岩性相似, 但与围岩的边界模糊, 单个遗迹化石几乎无法识别, 偶尔可见高角度倾斜的Palaeophycus heberti和水平分布的Planolites montanus, 形成于风暴浪基面附近、偶尔被远源风暴干扰的深水开阔台地沉积中(图 2); ⑤ Planolites beverleyensis遗迹组构, 由单种遗迹化石Planolites beverleyensis组成, 呈直或微弯曲的简单管状潜穴, 水平或近水平分布, 在层面上任意方向延伸, 无定向性, 潜穴之间常叠覆但未见分叉现象, 在整个地层中扰动强度变化不大, 扰动指数为1~2, 为多毛类留下的深阶层机会遗迹化石, 形成于高能动荡鲕粒滩环境中(图 2); ⑥ Thalassinoides bacae遗迹组构, 由单一的遗迹化石Thalassinoides bacae组成, 发育厚衬壁的三维潜穴系统, 在地层中多密集分布, 偶尔排列成簇状, 化石产状多样, 以高角度倾斜和垂直于层面分布为主, 生物扰动量变化较大, 形成在局限台地潮下低能环境中(图 2)。遗迹组构的演化指示了沉积环境的变化: 从平均浪基面之下的潮下深水低能环境变浅为平均浪基面之上的潮下浅水区, 开始发育鲕粒滩, 从滩前或滩间高低能交替环境加深过渡为碳酸盐岩靠海一侧的深水区, 最终以高能浅滩结束(局部发育滩间洼地和滩后局限台地)。整个过程反映了寒武纪中期底栖生物开始在碳酸盐岩台地殖居的状态。
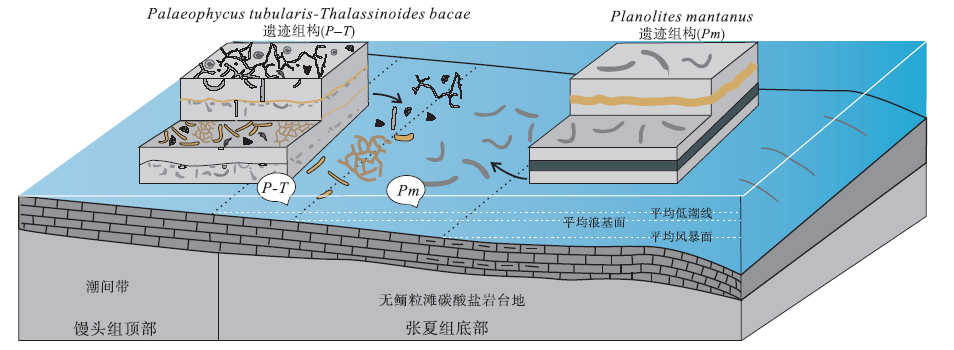 | 图 1 豫西张夏组下部遗迹组构与沉积环境分布模式(据齐永安等, 2012)Fig.1 Distribution model of ichnofabrics and sedimentary environments of the lower part of the Zhangxia Formation in western Henan Province(after Qi et al., 2012) |
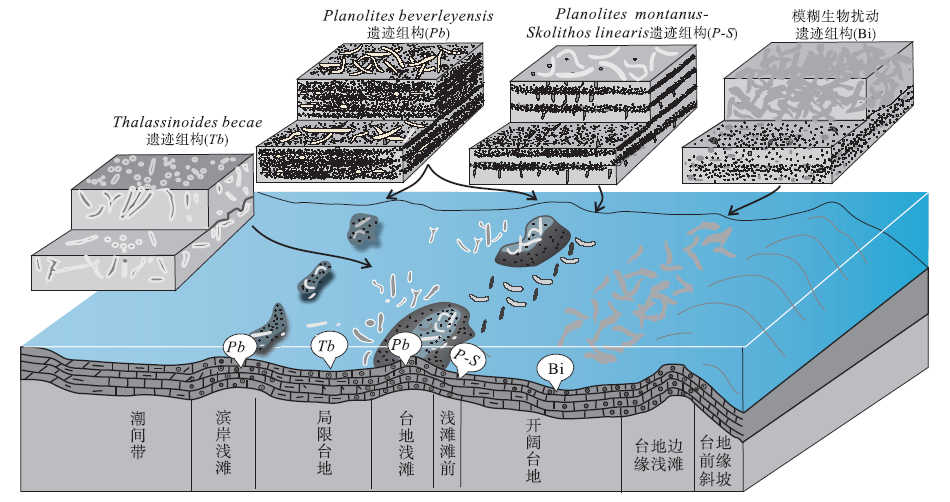 | 图 2 豫西张夏组中部遗迹组构与沉积环境分布模式(齐永安等, 2012)Fig.2 Distribution model of ichnofabrics and sedimentary environments of the middle part of the Zhangxia Formation in western Henan Province(after Qi et al., 2012) |
从下到上, 张夏组遗迹组构内部和遗迹组构之间均有一定的规律性: (1)每个遗迹组构内部生物扰动量均呈现增强趋势; (2)遗迹组构间阶层类型从属种单一且稀少的简单浅阶层类型到遗迹分异度和扰动强度增大的简单浅— 中阶层类型, 然后经过由模糊生物扰动为主的复杂浅— 中阶层类型, 最后过渡到简单深阶层类型; (3)沉积底质从张夏组底部的固底底质到中部的富含水分的混合底; (4)遗迹化石从不发育衬壁到发育有厚衬壁; (5)机会种遗迹化石在顶部出现等。这些规律综合表明在内栖生物向碳酸盐岩台地殖居的过程中, 生物扰动在数量和垂向上不断增强, 改变了沉积底质性质, 也带动了造迹生物行为习性的改变, 同时, 生物对沉积环境的适应性有了很大的提高。
地球早期生物扰动混合沉积物的可靠证据始于埃迪卡拉纪Nama组合带(Muscente et al., 2018)。在整个埃迪卡拉纪, 后生动物仅生活在沉积物表层, 穿透沉积物的能力较低, 偶尔对沉积物有些扰动(Chen et al., 2013, 2018; Meyer et al., 2014)。底栖生物在微生物席上行走留下的足迹与微生物席下掘穴开采形成的潜穴紧密相连, 揭示了埃迪卡拉纪两侧对称动物复杂的席下矿工行为, 但它们对原始沉积组构的改造可以忽略不计, 沉积— 水界面与下伏沉积物之间没有明显的营养物质交换, 还不能算是Bromley(1996)所界定的真正意义上的沉积物混合(Buatois et al., 2016)。生物扰动对沉积物的真正混合首次发生在前寒武纪— 寒武纪界线及其后的寒武纪幸运期, 以Treptichnids记录的三维分布的掘穴行为为代表(Mangano and Buatois, 2014), 但后生生物仅对原始沉积组构有轻微的改造, 与埃迪卡拉纪相比只是稍微加强了一些, 遗迹化石仅仅穿透沉积物表层数厘米深, 通常是平行于层面分布。
到寒武纪第二期, 中到高能的近滨砂岩中大量发育以Skolithos为代表的管状垂直潜穴, 对沉积物的扰动深度甚至可达1 m, 但它们是永久型居住结构, 混合沉积物的能力仍然不显著。随着寒武纪第三期以沉积进食潜穴为代表的遗迹化石多样性的增加, 沉积物的混合才变得越发普遍(Mangano and Buatois, 2014)。后生生物对沉积物的完全混合(扰动均质化)首次出现在寒武纪第四期, 以Thalassinoides形成的强烈扰动构造为代表(齐永安等, 2014b; Zhang et al., 2017)。这种交互巷道式潜穴代表着深层生物扰动者的行为, 它们能够自下向上将潜穴内溶质运移到层面上, 也可以通过增加沉积物— 水界面的面积将海水带入沉积物中, 促进了海水与沉积物之间的交换。生物扰动对沉积物的这种完全混合也发生在寒武纪古丈期, 表现为碳酸盐松软沉积物中留下的形态模糊的强烈扰动构造(齐永安等, 2012)。生物扰动对沉积物的混合不仅改变了沉积物的原始物理化学信息, 促进了海水与沉积物的生物地球化学循环, 而且诱导了沉积底质革命, 扮演了显生宙早期生态系统工程建造者的角色。
寒武纪是一个生物进化、生态学和地球生物学变化明显的时期。不同学者从不同的角度阐明了在这个生命历史关键时期发生的突破, 即寒武纪革命, 包括农艺革命(Seilacher and Pflüger, 1994; Seilacher, 1999)、寒武纪底质革命(Bottjer et al., 2000)和寒武纪信息革命(Plotnick et al., 2010)。这些概念现在经常被用于对寒武纪化石记录的观测和解释, 以及对埃迪卡拉纪和显生宙地球生物学特征的对比分析。
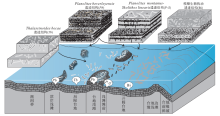
Seilacher(1999)将农业的发展和耕作效率的提高运用到寒武纪早期沉积底质的演化研究中。他刻画了微生物席底质(简称席底)特征: 浅海底质被微生物席或微生物膜覆盖或绑定, 隔绝了海底沉积物与海洋水体的任何交换, 沉积物— 水界面(微生物席)稳化固结, 缺乏明显的垂向生物扰动和生物对沉积物的混合, 导致低氧、低含水量的底质和沉积物。广泛发育的微生物席有助于以席底为基础的食物链和营养网的建立, 后生动物仅生活在沉积物/水界面很浅处, 即有微生物席的位置; 营养结构仅由初级生产者和分解者2个阶层组成。值得注意的是, 由于没有肉食捕食动物的存在, 这时期的环境很安稳, 被称为“ 埃迪卡拉花园” 。席底动物的进食策略和生活方式包括席包壳生物— 吸附在微生物席底表面、席黏附生物— 在微生物席底周围生长的生物、席抓扒生物— 啃食席底的生物、席消费者— 能够从微生物席的外部消化吸收的生物体、席牧食者— 生物在席底上面形成牧食迹或觅食迹、席下矿工— 在微生物席下建造巷道并且以最底层微生物的分解为生, 形成进食或者觅食构造(图 3)。生物扰动混合底质(简称混合底)则强调了垂向生物扰动作用的重要性, 包括深阶层的垂直潜穴(如Skolithos, Arenicolites, Diplocraterion, Lingulichnus, Syringomorpha)和中阶层水平结构(如Cruziana等)对沉积物的扰动改造。生物对沉积物的垂向扰动有利于水体与海底沉积物之间的流体交换, 扩大了适宜居住的底栖带, 增强了养分的循环利用(图 3)。农艺革命意味着在显生宙尽管压力环境局部存在(例如, 高盐潮坪、缺氧陆架、深海平原), 微生物持续丰富, 但广泛的席底在海洋环境中不再普遍存在(Seilacher, 1999)。
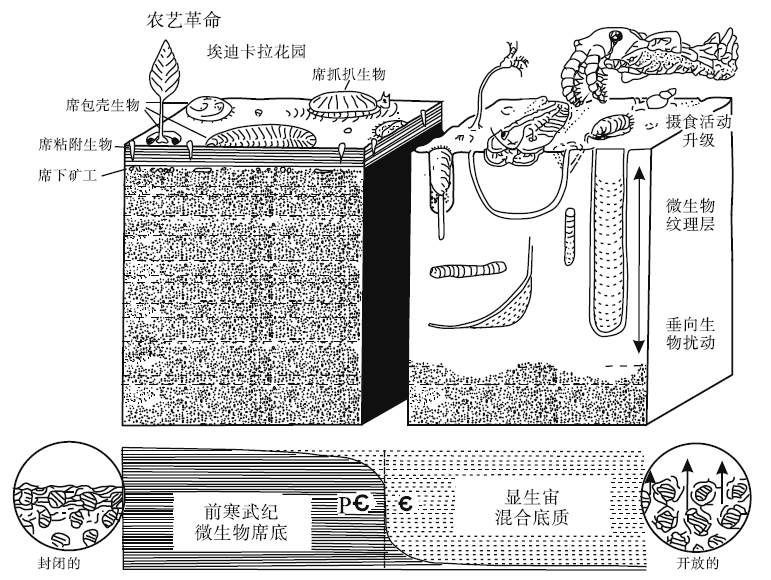 | 图 3 从前寒武纪微生物席底到显生宙生物扰动混合底质的转变(据Seilacher, 1999)Fig.3 Change from the Precambrian microbial matground to Phanerozoic bioturbated mixground(after Seilacher, 1999) |
寒武纪底质革命主要强调底质变化如何影响早期底栖生物的进化和生态效应, 可以被理解为底栖生物尤其是底栖生物群落对农艺革命诱发的底质进化的响应, 可进一步发展为与硬底的演化和复杂的有机体— 底质相互作用的联系, 底质的变化导致了早期后生动物的灭绝、适应以及不同的进化响应(Bottjer et al., 2000)。寒武纪信息革命详述动物对地形信息或环境信号的变化做出的反应, 侧重于海洋环境的信息内容和生物体获取和处理这些信息的能力, 强调协同进化。埃迪卡拉纪遗迹化石显示了行为简单的非定向觅食策略, 与之形成鲜明对比的是寒武纪幸运期遗迹化石表现出复杂的定向策略, 如趋向性、趋避性和趋触性, 也出现了系统分枝进食结构, 反映出造迹动物已发育复杂的感觉和神经系统(Plotnick et al., 2010; Mangano and Buatois, 2014)。
自2010年以来, 中国奥陶系、志留系和泥盆系中的遗迹化石研究主要集中在四川盆地、新疆塔里木盆地、准噶尔盆地以及河南、山西、陕西、贵州和广西等地。在奥陶系— 泥盆系海相及潮坪相中发现较多的遗迹化石, 据不完全统计, 已鉴定出51个遗迹属和107个遗迹种, 建立了4种遗迹相、19种遗迹组合及11种遗迹组构, 在遗迹化石及其沉积环境和生态学等研究方面取得了许多新进展。
在遗迹相研究方面, 张立军等(2011)对四川北川甘溪和广元后高坪、广西横县六景和桂林杨堤、贵州独山和贵阳乌当等地泥盆纪遗迹学与沉积学进行了系统研究, 鉴定和描述遗迹化石15属24种。根据遗迹化石之间的共生组合关系、实体化石特征和其他相标志, 识别出4种遗迹相: ① Skolithos遗迹相, 主要发育于浪控型海岸体系、潮控型海岸体系和障壁岛— 潟湖沉积体系的高能沉积区; ② Rhizocorallium遗迹相, 主要发育于潮控型海岸体系和障壁岛— 潟湖沉积体系的低能沉积区; ③ Cruziana遗迹相, 发育在浅海陆棚、具有丰富的生物及食物的砂泥岩和灰岩为主的低能沉积区; ④ Zoophycos遗迹相, 沿泥岩和泥灰岩发育的滨外沉积区分布。华南泥盆纪遗迹相从早泥盆世到晚泥盆世的演替规律是从Skolithos遗迹相→ Rhizocorallium遗迹相→ Cruziana遗迹相→ Zoophycos遗迹相→ Rhizocorallium遗迹相。早泥盆世晚期以后华南泥盆纪同时期的遗迹相自西南向东北的空间变化规律则表现为Zoophycos遗迹相→ Cruziana遗迹相→ Rhizocorallium遗迹相→ Skolithos遗迹相。
在塔里木盆地, 在上奥陶统柯坪塔格组下段中— 厚层细砂岩和粉砂岩薄层中识别的遗迹化石有5属9种, 这些遗迹化石可归于Cruziana遗迹相, 属浅海碎屑岩沉积相带(丁奕等, 2015)。在柯坪一带志留系依木干他乌组见有较多遗迹化石, 鉴定出6属7种, 解释为潮坪沉积环境(白忠凯等, 2017)。另外, 在靠近滇黔桂古陆半封闭海湾环境的重庆秀山地区, 于下志留统龙马溪组上部的灰色页岩中发现属于潮坪沉积环境的双菌迹(Bifungites)遗迹化石(黄再琴和王约, 2011)。
有关遗迹组合的研究, 在中国四川盆地、新疆准噶尔盆地和华北盆地都做了很好的工作。四川龙门山地区早— 中泥盆世地层中发现遗迹化石较多, 研究较详, 已识别出15个遗迹属(张立军和龚一鸣, 2013; Zhang, 2014; Zhang and Zhao, 2015)。在甘溪剖面下泥盆统的滨浅海相地层中, 根据遗迹化石组成及其沉积特征, 建立了8种遗迹组合: ① Planolites-Palaeophycus遗迹组合, 主要由觅食迹组成, 指示低能的砂质近滨环境; ② Rusophycus-Phycodes遗迹组合, 主要是觅食迹和停息迹, 形成于低能的近滨环境; ③ Chondrites-Palaeophycus遗迹组合, 主要是居住迹和觅食迹, 反映了贫氧低能的近滨下部环境; ④ Skolithos-Cylindrichnus遗迹组合, 主要由居住迹组成, 形成于高能的前滨环境; ⑤Skolithos和 Skolithos-Diplocraterion遗迹组合, 主要为居住迹和觅食迹, 反映了低能、食物丰富的近滨上部环境; ⑥Chondrites-Palaeophycus遗迹组合, 主要为居住迹、觅食迹和化能型遗迹, 反映了低能、贫氧的滨岸潟湖环境; ⑦Rusophycus遗迹组合, 主要为觅食迹和居住迹, 反映了低能、食物丰富的远滨上部环境; ⑧Zoophycos遗迹组合, 主要为觅食迹和牧食迹, 反映了受风暴影响的远滨下部环境。此后, 荆锡贵等(2018)在研究下泥盆统平驿铺组、白柳坪组、甘溪组、谢家湾组和养马坝组二台子段以及中泥盆统养马坝组养马坝段和观雾山组遗迹化石时, 又新建2种遗迹化石组合: ① Skolithos-Thalassinoides遗迹组合, 这一组合与上述第④组合(Skolithos-Cylindrichnus遗迹组合)的化石组成类似, 同样出现于高能砂质沉积环境中, 为水动力较强的无障壁滨岸前滨环境; ② Zoophycos-Palaeophycus遗迹组合, 产生于正常浪基面以下、风暴浪基面以上的风暴沉积环境。姜涛等(2018)在该套地层中专门研究了Chondrites(丛藻迹), 识别出8种类型, 其他遗迹化石还有Phycodes palmatus, Palaeophycus striatus, Cruziana rouaulti, Lockeia siliquaria和Palaeophycus tubularis等。基于这些遗迹化石在地层中的产状、分布及组合特征, 又划分出2种遗迹组合: ①Chondrites-Phycodes遗迹组合, 大多分布于下泥盆统平驿铺组灰黑色粉砂质泥岩中, 主要由Chondrites type-C, Chondrites filiformis, Phycodes palmatus和Palaeophycus striatus等组成, 产生于相对较深水、食物充足、氧含量相对较低的浅水陆棚沉积环境; ②Chnodrites-Lockeia遗迹组合, 发育于下泥盆统白柳坪组砂质沉积为主的岩层中, 由Chondrites type-C, Chondrites isp., Chondrites intricatus, Cruziana rouaulti, Lockeia siliquaria和Palaeophycus tubularis等组成, 产于氧含量充足的常氧相浅水陆棚沉积环境。
在西准噶尔地区晚泥盆世— 早石炭世深海沉积中已识别出大量遗迹化石, 鉴定出16属21种(Fan and Gong, 2016a)。基于这些遗迹化石的沉积背景分析, 建立了3种遗迹组合: ① Helminthopsis-Helminthoidichnites-Arenituba遗迹组合, 产于受密度流影响的沉积中; ② Zoophycos-Nereites-Phycosiphon遗迹组合, 出现在典型的斜坡沉积中; ③ Phycosiphon-Nereites遗迹组合, 发育在受底流影响的斜坡下部或盆地底部泥质沉积中。
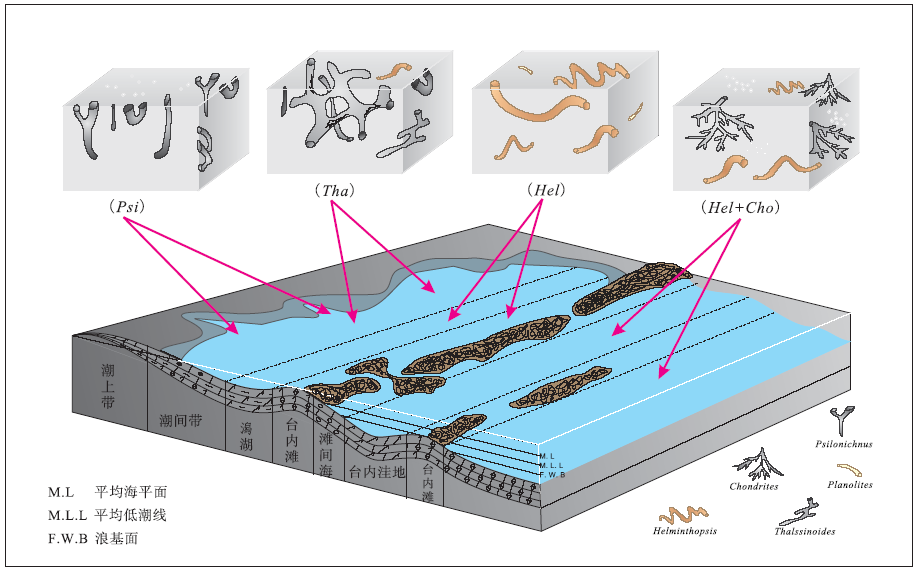
在豫西北奥陶系马家沟组碳酸盐岩中共识别出三大生态类型的遗迹化石, 共10个遗迹属和17个遗迹种。按其古生态和沉积学特征, 可划分出3种遗迹组合(牛永斌等, 2015): ① Psilonichnus-Thalassinoides 遗迹组合, 代表局限碳酸盐岩台地内的潮坪环境; ② Helminthopsis-Gordia 遗迹组合, 反映局限碳酸盐岩台地的潟湖或开阔碳酸盐岩台地的滩间海和台内洼地等低能、静水的沉积环境; ③ Chondrites-Scolicia遗迹组合, 指示开阔碳酸盐岩台地内洼地中氧化还原界面附近的沉积环境。
近几年来, 中国奥陶系和泥盆系碳酸盐岩和碎屑岩中的生物扰动与遗迹组构研究受到关注。豫西北奥陶系马家沟组豹斑灰岩中“ 豹斑” 构造形态多样, 经研究是由多种生物潜穴构成, 按其形态可划分为Psilonichnus, Chnodrites, Thalassinoides, Helminthopsis及Planolites等5种生物潜穴类型(董小波和牛永斌, 2015b)。不同类型生物潜穴的出现主要受到沉积环境的制约。生物潜穴充填物按照白云石化程度可被划分为泥晶白云石充填物及粉— 细晶白云石充填物2种类型, 其中泥晶白云石主要存在于Psilonichnus及Chnodrites这2种潜穴中, 而粉— 细晶白云石则主要存在于Thalassinoides, Helminthopsis及Planolites之中; 潜穴充填物的白云石化过程可被分为白云石化前的准备阶段、白云石的形成阶段及白云石的后期调整阶段, 不同潜穴的白云石化状态差异就是在此过程中形成的。陈曦等(2011)研究陕西韩城— 旬邑地区中奥陶统马家沟组豹斑白云岩时明确表示, 掘穴生物活动形成的潜穴是造成不均匀白云岩化、形成豹斑的重要原因。基于对奥陶系马家沟组中生物扰动构造形态、类型及其分布特征的研究, 已在马家沟组三段海相碳酸盐岩中识别出遗迹化石7属10种(牛永斌等, 2018b), 建立了4种遗迹组构(图 4): ① Psilonichnus遗迹组构, 主要为Psilonichnus upsilon, 遗迹形态包括“ I” 形、“ Y” 形、“ U” 形和“ J” 形等, 发育在潮上带与潮间带上部(高潮坪)低能的沉积环境; ② Thalassinoides遗迹组构, 以Thalassinoides callianassa, Thalassinoides horizontalis为特征, 另外还有少量的Psilonichnus upsilon和Helminthopsis isp., 代表潮间带中— 下部和台内滩等较高能的沉积环境; ③Helminthopsis遗迹组构, 遗迹化石主要由Helminthopsis abeli, H. isp., H.hieroglyphica, Thalassinoides horizontalis, Palaeophycus tubularis, Planolites montanus组成, 产生于局限碳酸盐岩台地内的潮下带— 潟湖和开阔碳酸盐岩台地内的滩间海等较低能的沉积环境; ④ Helminthopsis-Chnodrites遗迹组构, 常见遗迹化石包括Helminthopsis abeli, H. hieroglyphica, Chnodrites intricatus, Palaeophycus tubularis, Planolites montanus和Gordia molassica等, 形成于开阔碳酸盐岩台地内滩间海— 台内洼地等低能沉积环境。刘梦瑶等(2020)进一步精细解析了山西省兴县中奥陶统马五5亚段中豹皮灰岩, 认为豹皮状灰岩主要与遗迹化石Thalassinoides潜穴系统有关, 白云石斑块沿生物潜穴分布, 潜穴内部为白云石, 围岩部分为方解石, 白云石斑块保存了完整的生物潜穴形貌, 表明生物潜穴内部优先发生了白云石化作用, 因此生物扰动作用是豹斑灰岩/豹斑白云岩形成的重要因素。奥陶纪海相碳酸盐岩地层中大量发育的Thalassinoides三维潜穴系统为白云石化流体提供了良好的运移通道, 为回流渗透作用提供了有利条件, 促进潜穴充填物白云石化, 从而形成了大量豹皮状的生物扰动白云岩(图 5)。
 | 图 4 豫西奥陶系马家沟组三段遗迹组构及其沉积环境分布模式(据牛永斌等, 2018b)Fig.4 Ichnofabrics and their sedimentary environments of the Member 3 of the Ordovician Majiagou Formation in western Henan Province(after Niu et al., 2018b) |
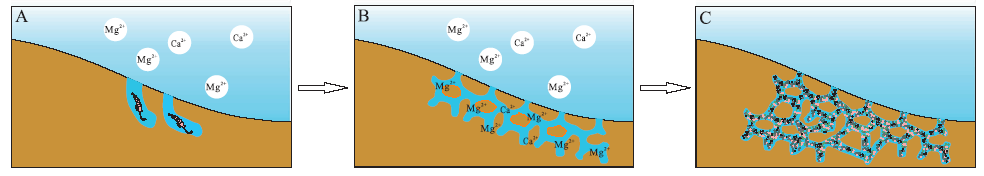 | 图 5 生物扰动对白云岩回流渗透的促进作用(据刘梦瑶等, 2020) A— 造迹生物在沉积物中掘穴; B— 造迹生物的掘穴作用形成三维连通的潜穴系统, 白云石化流体沿潜穴迁移; C— 潜穴充填物发生白云石化作用Fig.5 Role of bioturbation in promoting reflux permeability of dolomite stone(after Liu et al., 2020) |
在新疆西准噶尔地区晚泥盆世— 早石炭世海相沉积中也发现大量生物扰动构造, 已识别出18种遗迹化石, 建立了5种遗迹组构(Fan and Gong, 2016b): ① Chondrites-?Phymatoderma 遗迹组构, 产生在下临滨— 滨外浅海钙质、泥质粉砂岩、钙屑灰岩和钙质泥岩沉积中; ② Rhizocorallium commune遗迹组构, 出现在高能下临滨环境, 常位于钙质风暴岩之上; ③ 海葵居住潜穴(Dolopichnus gulosus)遗迹组构, 产自滨外过渡带或浅海沉积环境; ④ Teichichnus遗迹组构, 发育在滨外过渡带或浅海细粒沉积及厚层状中— 细粒风暴岩中; ⑤ Zoophycos遗迹组构, 见于滨外过渡带远端风暴岩(极细砂岩和粉砂岩)及浅海至斜坡上部中厚层状硅化凝灰质粉砂岩中。
Zhang等(2015a)基于全球不同地区291篇论文(1821-2015)和不同时代地层(寒武系至古近系)中180件动藻迹(Zoophycos)样品数据的分析, 构建了显生宙动藻迹的数据库, 从地球生物学角度系统归纳总结后, 发现了显生宙动藻迹3个方面的宏演化规律及其环境背景。 (1)显生宙动藻迹的丰度和纬度分布主要受控于显生宙的生物兴衰和潘基亚超大陆的聚散, 与全球气候和温度的变化并没有关系。显生宙的生物灭绝事件和潘基亚超大陆的聚合不仅导致海生无脊椎动物分异度的降低, 也导致动藻迹丰度分布的最低值。浅海区面积减少和生物之间的生存竞争的加剧, 中— 新生代浮游生物的繁盛是导致动藻迹造迹者向半深海— 深海迁移的主要生物— 环境背景。(2)从古生代至新生代, 动藻迹的环境分布表现为从浅阶层迁移至深阶层, 从浅海迁移至半深海— 深海, 显生宙动藻迹这种阶层和水深分布趋势体现了动藻迹的造迹生物对非宜居生境的适应辐射。显生宙海洋生产力的升高, 不仅导致了滨— 浅海区浅阶层生存环境竞争的增加, 也促使生物向非宜居的半深海— 深海和深阶层的适应辐射, 随着中— 新生代海底营养条件的改善, 最终导致了动藻迹在白垩纪— 新生代半深海— 深海区和深阶层的繁盛。(3)显生宙动藻迹的形貌结构由简单到复杂、由粗放到集约、由低级到高级的变化, 清楚地反映了动藻迹的造迹生物智慧和文化的进化, 表明生物圈、生态系和生物类群以及生物文化的发展演替均遵从进化论揭示的基本规律。
广西、四川和豫西南淅川地区上泥盆统浅海相碳酸盐岩中发育丰富的根珊瑚迹(Rhizocorallium), 在该遗迹化石的边缘管和蹼纹不同区域中具有形态大小不同的莓状黄铁矿颗粒, 反映出潜穴内部微区环境的变化过程。研究显示, 根珊瑚迹潜穴中莓状黄铁矿的存在可能与硫酸盐还原菌有关, 其造迹生物的生命活动中与之共生的微生物使原始沉积物中的氧化剂被逐渐还原, 造成潜穴内部氧含量下降, 并且靠近中部的蹼纹形成环境较之边缘管更近封闭, 含氧量进一步降低, 从而出现从边缘管向蹼纹内部莓状黄铁矿的单晶颗粒逐渐缩小的趋势。由此推断, 根珊瑚迹可能是造迹者精心构筑的觅食— 花园, 根珊瑚迹的造迹者在边缘管和蹼纹中, 由于食物供应和氧化还原条件的不同, 培植不同的微生物, 与其形成一个互利共生的群落(Zhang et al., 2016; Zhang and Zhao, 2016; 赵曌和张立军, 2017; Zhao et al., 2019)。
此外, 在华南古热带区广西来宾一带浅海灰岩中常见保存完好的动藻迹(Zoophycos), 其造迹者以浅海底质沉积物中富营养物为食, 这些有机营养物就是与其共生的微生物(硫酸盐还原菌)。基于对动藻迹蹼状构造中丰富黄铁矿莓球粒的分析, 可以认为, 这些动藻迹不仅仅是造迹者运移过程中产生的生物成因沉积构造, 而且是由造迹者与共生微生物一起进行一系列的生命活动行为所形成的复杂精美的生物成因沉积构造(Zhang et al., 2015b)。
中国石炭纪— 二叠纪海相沉积中含有丰富的遗迹化石, 以往研究程度较高, 近10年在新疆准噶尔盆地西北部、柴达木盆地南缘和湖南中部(石炭纪)、晋中南(晚石炭— 早二叠世)、豫西和内蒙古(早二叠世)、广西和安徽巢湖(早— 中二叠世)以及四川广元和重庆(晚二叠世)等地区又取得许多新的研究成果。在潟湖、浅海陆棚、半深海及深海盆地沉积环境中识别遗迹化石计有23个遗迹属51个遗迹种, 建立了7种遗迹组合及10种遗迹组构, 并开展了遗迹化石与古氧相和地微生物相关性的探索, 为中国石炭纪— 二叠纪地层的沉积环境分析提供了遗迹学方面的重要信息。
新疆准噶尔盆地西北部下石炭统塔尔巴哈台组上部含丰富的植物茎干化石和遗迹化石, 已识别的遗迹化石有8个遗迹种并含有Nereites遗迹相的典型分子, 绝大多数都是半深海— 深海浊积岩中的常见属种, 说明塔尔巴哈台组上部地层形成于半深海— 深海沉积环境(纵瑞文等, 2014)。在柴达木盆地南缘下石炭统石拐子组和上石炭统缔敖苏组中发现了大量Chondrites遗迹化石, 研究认为下石炭统石拐子组产生于台洼沉积环境, 上石炭统缔敖苏组形成于局限碳酸盐岩台地沉积环境(牛永斌等, 2008)。在湘中下石炭统中发现13个遗迹属17种不同类型的遗迹化石, 建立了Fucusopsis, Skolithos, Palaeophycos, Rhizocorallium和 Chondrites等5个遗迹组合, 分别产于潮坪— 潟湖、障壁砂坝、滨外碎屑陆棚、滨外浅水碳酸盐陆棚及深水碳酸盐陆棚沉积环境(Shao et al., 2011; 邵龙义和刘钦甫, 1994)。
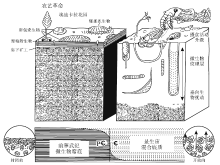
华北晋东南地区上石炭统— 下二叠统太原组是一套由陆源碎屑岩、煤和海相碳酸盐岩组成的海陆交互相沉积, 其中碳酸盐岩中遗迹化石丰富, 已识别出10个遗迹属14个遗迹种(胡斌等, 2013), 建立了4种遗迹组合(图6)。 ① Zoophycos-Thalassinoides 遗迹组合, 以 Zoophycos, Chondrites, Thalassinoides 和Ophiomorpha 为特征, 形成于受陆源碎屑影响的海湾— 潟湖环境; ②Planolites-Helminthopsis 遗迹组合, 主要由Zoophycos, Palaeophycus, Helminthopsis, Planolites 和Rhizocorallium 等组成, 产生于平均海平面之下至正常天气浪基面附近的浅海上部沉积环境; ③Zoophycos-Teichichnus 遗迹组合, 以 Zoophycos 和Teichichnus 共生为特征, 出现于正常天气浪基面之下至风暴浪基面之上的浅海中部沉积环境; ④ Nereites-Chondrites 遗迹组合, 主要由 Zoophycos, Nereites 和 Chondrites 组成, 发育于风暴浪基面之下、 水体较深且贫氧的浅海下部沉积环境。
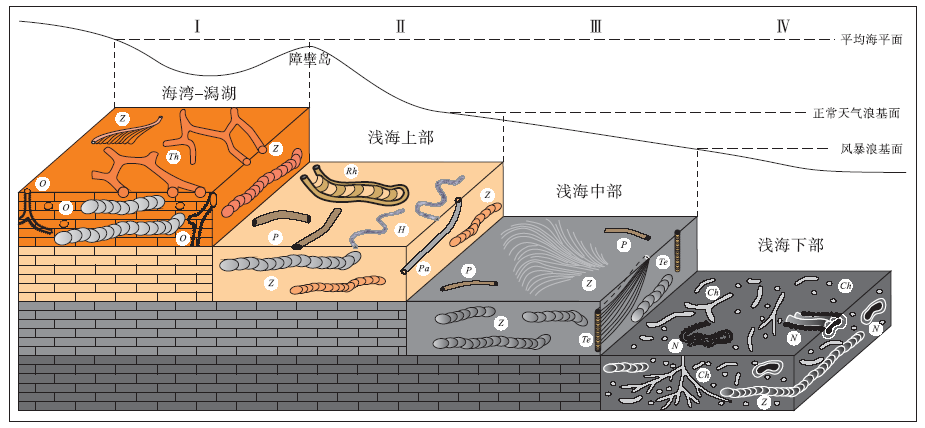 | 图 6 晋东南地区太原组遗迹化石组合的沉积环境模式(据胡斌等, 2013) Ⅰ -Zoophycos-Thalassinoides遗迹组合; Ⅱ -Planolites-Helminthopsis遗迹组合; Ⅲ -Zoophycos-Teichichnus遗迹组合; Ⅳ -Nereites-Chondrites遗迹组合; Ch-Chondrites; H-Helminthopsis; N-Nereites; O-Ophiomorpha; Pa-Palaeophycus; P-Planolites; Rh-Rhizocorallium; Te-Teichichnus; Th-Thalassinoides; Z-ZoophycosFig.6 Sedimentary environmental model of ichnoassemblages in the Taiyuan Formation in southeastern Shanxi Province(after Hu et al., 2013) |
河南境内下二叠统太原组遗迹化石及沉积环境的研究程度相对较高。胡斌等(2010)对豫西北焦作地区太原组的遗迹化石研究中, 识别出10个遗迹属11个遗迹种, 建立了3种遗迹组合: ①Zoophycos-Nereites遗迹组合, 主要由Zoophycos, Nereites, Chondrites, Teichichnus和较强生物扰动层组成, 遗迹化石丰度较高, 出现在水体相对较深、浪基面之下且沉积底层内氧气贫乏的远岸浅海沉积环境; ②Zoophycos-Taenidium遗迹组合, 以大量Zoophycos成层出现为特色, 伴生Planolites, Taenidium和Rhizocorallium等遗迹化石, 产生于水体相对较浅的浪基面之上的近岸浅海沉积环境; ③Ophiomorpha-Thalassinoides遗迹组合, 由Zoophycos, Rhizocorallium, Thalassinoides, Ophiomorpha 和Gordia组成, 发育于海岸带潮坪或海湾、潟湖沉积环境。
宋慧波等(2011)通过对河南省内46个代表性钻孔以及露头剖面的岩性、沉积特征、实体及遗迹化石的分析, 将太原组沉积划分出5种沉积相(潮坪相、潟湖相、障壁岛相、局限台地和开阔台地相)、3种沉积组合(局限台地— 潮坪相组合、开阔台地— 潮坪— 潟湖相组合、开阔台地— 障壁岛— 潮坪相组合)及3种岩相区(局限台地— 潮坪泥晶灰岩、开阔台地— 潮坪— 潟湖灰岩、开阔台地— 障壁岛— 潮坪灰岩)。在利用遗迹化石进行地层和沉积相对比方面, 探索性地将Zoophycos潜穴充填物颜色及含氧量变化与古水深结合起来分析, 提出了深水型Zoophycos遗迹化石的识别特征, 可作为太原组区域地层及沉积相的对比标志(平瑞等, 2013), 并基于沉积环境的时空对比分析, 提出了豫西地区太原组沉积期海水进退变化规律(胡斌等, 2016)。华北北部内蒙古西乌旗地区下二叠统寿山沟组中识别出7个遗迹属, 划分出 Zoophycos 和Helminthoida-Helminthopsis遗迹化石组合, 均属 Zoophycos 遗迹相, 反映了半深海环境的大陆斜坡沉积(黄欣等, 2013)。
中国华东和华南地区下— 中二叠统栖霞组和茅口组灰岩中Zoophycos潜穴的研究较深入。王冰等(2012)在安徽巢湖中二叠统栖霞组中识别出Zoophycos遗迹相, 表明巢湖地区中二叠统栖霞组臭灰岩段和顶部灰岩段属于贫氧的碳酸盐岩台缘斜坡沉积。张立军等(2015)专门研究了广西来宾地区栖霞组中部和茅口组下部泥粒状灰岩及粒泥状灰岩中大量Zoophycos(动藻迹)遗迹化石的生态习性特征, 基于对大量Zoophycos边缘管、蹼层、大蹼纹和小蹼纹的产状及行为方式的详细分析, 重建了Zoophycos 3-D形态。
中国晚二叠世遗迹化石研究仅在重庆巫溪和四川北部广元一带开展了工作。重庆巫溪红池坝乐平统长兴阶大隆组中, 已识别出遗迹化石8属13种, 表明该地大隆组沉积期(长兴中— 晚期)沉积环境由深水盆地向深水陆棚逐渐过渡, 基于时间序列的遗迹化石记录显示出深水陆棚水体含氧量频繁加速波动的过程, 大隆组下部为潮下深水陆棚环境, 中部硅质页岩缺乏生物扰动, 为深水缺氧盆地环境, 上部潜穴类型多样, 指示水循环良好的深水陆棚环境(杨巍等, 2018)。四川北部广元上寺乐平统大隆组中上部海相硅质岩、硅质灰岩中赋存较为丰富的遗迹化石, 已识别出觅食迹和居住迹5属7种, 基于遗迹化石组成及其分布特征、生物扰动指数、实体化石和地球化学分析, 可以认为该地区大隆组中上部硅质岩中发育小型Chondrites潜穴(直径1.0~1.5 mm), 整体处于水动力较弱、滞流缺氧或贫氧环境, 但不同时期含氧水平有所波动, 到本组顶部变为缺氧富硫或贫氧富氧交替出现的沉积环境(张鑫和张立军, 2019)。
关于石炭纪— 二叠纪地层中遗迹组构的研究, Gong等(2010)详细分析了华南广西来宾地区栖霞组和茅口组浅海灰岩中的Zoophycos 复合遗迹组构, 相似的复合遗迹组构也发育在澳大利亚悉尼盆地的中二叠统Broughton组Westly Park 砂岩段的滨海相杂砂岩中。基于对Zoophycos潜穴的形态和构造特征及多期次相互交切关系的研究, 论述了造迹生物营造的多阶层(浅阶层、中阶层和深阶层)特征, 以及从软底(softground)到僵底(stiffground)再到固底(firmground)的变化规律。
宋慧波等(2012)研究了豫西地区下二叠统太原组灰岩层中发育的生物扰动构造, 将太原组碳酸盐岩中的遗迹化石划分为5种遗迹组构, 包括类型A— Gordia-Planolites遗迹组构、类型B— Rhizocorallium-Thalassinoides遗迹组构、类型C— Zoophycos-Taenidium遗迹组构、类型D— Zoophycos-Speckle burrow(斑状潜穴)遗迹组构和类型E— Chondrites-Nereites遗迹组构。这些遗迹组构主要分布于潟湖潮坪和台内浅滩(类型A)、正常天气浪基面之上的浅海上部或局限台地(类型B)、正常天气浪基面之下的开阔台地(类型D)和风暴浪基面之下的浅海下部或水动力条件类似的较闭塞的沉积环境(类型E)。
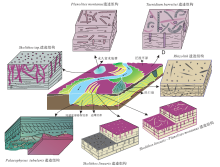
宋慧波等(2015)根据对晋中南地区太原组碳酸盐岩中遗迹化石及其宿主岩特征的综合分析, 识别出10个遗迹属14个遗迹种, 建立了4种遗迹组构及其沉积环境分布模式。 类型A— 具褐红色、灰白色充填物的大型Zoophycos-Thalassinoides遗迹组构, 出现于海湾(潟湖)及潮坪沉积环境; 类型B— 具红色、灰白色充填物或带衬壁的大型Zoophycos-Planolites遗迹组构, 发育在海平面至正常天气浪基面附近区域的浅海上部沉积环境或水体浅且较平静的局限碳酸盐岩台地环境; 类型C— 具灰色、深灰色充填物的中型Zoophycos-Teichichnus遗迹组构, 主要产生于正常天气浪基面之下、风暴浪基面之上的浅海中部沉积环境; 类型D— 具深灰色— 黑色充填物的小型Zoophycos-Chondrites遗迹组构, 形成于风暴浪基面之下、水体较深且贫氧的浅海下部沉积环境。按照生物扰动构造的阶层分布特征, 可将太原组灰岩中的遗迹组构划分为4种类型(图 7): ① 表层阶层遗迹组构(Epigenic tiering ichnofabric), 以大量出现在层表面的Gordia和二分叉形遗迹(Dichasialichnus jiaozuoensis)为特征, 有时会变为Helminthopsis和 Planolites, 具有高丰度、低分异度的特点, 生物扰动指数BI为 2~3, 产生于低能局限海湾或潟湖环境; ②浅层阶层遗迹组构(Shallow tiering ichnofabric), 由出现在底层上部的大型Zoophycos并常常伴生Rhizocorallium, Taenidium 或 Planolites 和 Palaeophycus等遗迹化石构成, 有时出现Skolithos, 中— 高丰度和分异度, 生物扰动深度一般位于层面以下15 cm内, 生物扰动指数BI为3~5, 形成于平均低潮线与正常天气浪基面之间的浅海上部环境; ③中阶层遗迹组构(Medium tiering ichnofabric), 以中型Zoophycos并伴生Teichichnus 和Taenidium遗迹化石为特征, 有时变为居住迹Skolithos 和 Thalassinoides, 中— 高丰度和分异度, 生物扰动指数BI为3~5, 生物扰动深度一般渗入层面以下15~25 cm内, 发育于正常天气浪基面与风暴浪基面之间的浅海中部环境; ④深阶层遗迹组构(Deep tiering ichnofabric), 主要由小— 中型具有深灰色— 黑色潜穴充填物的Zoophycos并伴随Chondrites 和 Nereites遗迹化石组成, 具有高丰度、低分异度的特点, 生物扰动指数BI为2~5, 生物扰动深度可渗入到层面以下25~35 cm内, 产生于风暴浪基面以下至氧化还原界面附近较深水的浅海下部环境(Song et al., 2015)。
 | 图 7 华北地区太原组中遗迹组构的沉积环境模式(据Song et al., 2015) Ch-Chondrites; D-Dichasialichnus; G-Gordia; H-Helminthopsis; N-Nereites; Pa-Palaeophycus; Pl-Planolites; Rh-Rhizocorallium; Sk-Skolithos; Ta-Taenidium; Te-Teichichnus; Th-Thalassinoides; Z1-Zoophycos with red filling; Z2-Zoophycos with greywhite filling; Z3-Zoophycos with grey filling; Z4-Zoophycos with dark grey filling; Z5-Zoophycos with grey-black filling; Z6-Zoophycos with black fillingFig.7 Sedimentary environment model of ichnofabrics in the Taiyuan Formation of North China(after Song et al., 2015) |
沉积地层中古氧相的判识有多种途径, 采用岩石学、生物学和地球化学方法分析富/常氧(oxic)、贫氧(dysoxic)、极贫氧(suboxic)、缺氧或厌氧(anoxic)环境都各有一定的特征指标或标志。同样, 遗迹学方法也可用于古氧相分析, 因为生物对其殖居地所处生态环境具有高度敏感性, 所以生物在原地营造的遗迹也必然是当时古环境及古氧相条件下的产物。宋慧波等(2017)基于对豫西禹州和焦作地区下二叠统太原组中的遗迹化石组成与产状特征的研究, 以及对不同颜色 Zoophycos 潜穴充填物中碳氧同位素和微量元素含量的测试分析, 提出了 3种古氧相(富/常氧相、贫氧相和厌氧相)的遗迹化石响应关系及其形成的沉积背景(图 8)。(1)富/常氧相。遗迹化石主要以层内潜穴Zoophycos, Teichichnus, Planolites, Palaeophycus, Thalassinoides 和 Skolithos为主, 其次为层面觅食迹 Dichasialichnus, Gordia和Helminthopsis 以及少量生物扰动构造。Zoophycos 潜穴充填物颜色常见为灰白色和灰色, δ 13C值较低, 主要介于-0.8‰ ~-4‰ 之间, V/(V+Ni)值小于0.46, Ce/La值小于1.5。所有上述这些特征均反映水体较浅的氧化环境, 发育于浅海上部至滨岸带的碳酸盐岩台地沉积背景。(2)贫氧相。遗迹化石主要由层内潜穴 Zoophycos, Planolites 和 Taenidium 组成, 丰度较高, 但分异度较低。在产状上, Zoophycos 在垂向上分布有多层近水平的板状蹼层, 蹼层宽度一般为1.5~2.5 mm, 表明遗迹化石为中到小型个体。Zoophycos 潜穴充填物颜色常为深灰色或黑色和红褐色, δ 13C值大多介于-0.2‰ ~-1‰ 之间, 比灰色充填物的δ 13C值高, V/(V+Ni)值大于 0.46。所有上述这些特征均反映水体相对较深或者较闭塞的弱还原环境, 形成于浅海中下部或有障壁海岸较闭塞的海湾潟湖或局限碳酸盐岩台地沉积背景。(3)厌氧相。遗迹化石主要以层内潜穴Zoophycos, Chondrites 和Nereites 组合为特征。在产状上, Zoophycos 在垂向上分布有多层近水平的板状蹼层, 蹼层宽度一般都在1 mm 左右, 表明遗迹化石个体较小。Zoophycos 潜穴充填物颜色均为黑色, δ 13C值较高, 多数在0.2‰ ~1.6‰ 之间, V/(V+Ni)值小于0.46, Ce/La值大于1.8。上述特征均反映水体较深的厌氧环境, 产生于浅海下部(深水陆棚)或较闭塞的碳酸盐岩台地沉积背景。
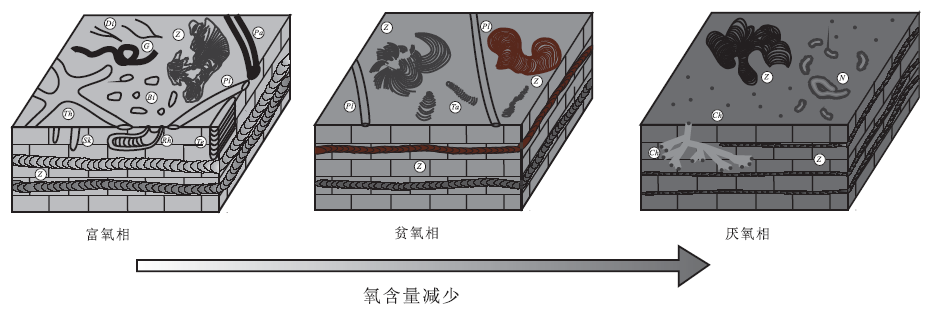 | 图 8 豫西太原组遗迹化石与古氧相响应特征模式(据宋慧波等, 2017) Bi-Bioturbation structure; Ch-Chondrites; Di-Dichasialichnus; G-Gordia; N-Nereites; Pa-Palaeophycus; Pl-Planolites; Rh-Rhizocorallium; Sk-Skolithos; Ta-Taenidium; Te-Teichichnus; Th-Thalassinoides; Z-ZoophycosFig.8 Characteristic model of ichnofossils responding to paleo-oxygen facies of the Taiyuan Formation in western Henan Province(after Song et al., 2017) |
随着研究的深入, 深时地球微生物的研究受到地学界的广泛关注, 逐渐成为新的研究热点。近几年遗迹学领域也开始了对这方面的探索。宋慧波等(2014)从地球微生物学的角度出发, 对不同颜色充填物的Zoophycos潜穴进行研究时, 发现了大量成因与微生物相关的碳酸盐岩微形体, 识别出球状、杆状、簇状、网状、瓶状和似脑球状等 6种形态类型, 其中多数显示出不同的微生物结构, 因此Zoophycos遗迹化石并非为简单的觅食迹或食物储藏所, 其造迹生物与微生物很可能存在互利共生关系。Zhang等(2015b)在研究广西来宾早、中二叠世浅海碳酸盐岩中的Zoophycos时, 在Zoophycos蹼层中也发现丰富的草莓状黄铁矿球粒, 这很可能与硫酸盐还原细菌有关。基于对Zoophycos蹼层中发现的大量黄铁矿莓球粒的分析, 可以认为这些复杂精美的生物成因构造是Zoophycos造迹生物与共生的微生物(如硫酸盐还原细菌)一系列生命行为作用的产物。
中国三叠纪及P/Tr界线附近海相遗迹化石的研究主要集中在扬子地区, 尤其是上扬子地区的四川和贵州以及下扬子地区的皖南、苏西南和浙北地区的早三叠世遗迹化石研究较详, 晚三叠世遗迹化石研究仅限于四川盆地。已报道的三叠纪遗迹化石有66个遗迹属和110个遗迹种, 建立遗迹组合与组构各5种, 其中早三叠世遗迹化石大多分布在碳酸盐台地及台缘下斜坡浊流沉积环境, 而晚三叠世遗迹化石主要形成于潮坪沉积环境。
四川盆地南部赤水地区下三叠统嘉陵江组主要为一套海相碳酸盐岩及蒸发岩, 发现遗迹化石4个遗迹属8个遗迹种, 可划分出2种遗迹组合(刘治成等, 2009): ①Palaeophycus 遗迹组合, 常见Palaeophycus和Helminithopsis遗迹化石, 分异度低, 丰度中— 高, 生物扰动中等, 主要发育于薄— 中厚层状浅灰色、灰色泥晶灰岩夹紫灰色中厚层状亮晶生物碎屑灰岩和棕褐色、褐色泥岩中, 代表水体较浅、受短期海平面变化影响较大的灰岩坪沉积环境; ②Planolites-Helminithopsis 遗迹组合, 由Planolites, Helminihopsis, Phycodes 和 Palaeophycus 等遗迹化石组成, 分异度中等, 丰度中— 高, 生物扰动中— 高, 代表的是与 Palaeophycus 遗迹组合相比水体较深、能量较低的开阔海台地的台内洼地环境。四川峨眉山龙门洞一带下三叠统嘉陵江组一段(嘉一段)碎屑岩中已鉴定出9种遗迹化石, 识别出2种遗迹组构(张国成和王昆, 2010): ① Arenicolites遗迹组构, 主要发育在嘉一段细砂岩中, 组成分子包括Arenicolites, Diplocraterion, Skolithos和Psilonichnus, 丰度较高, 分异度中等, 产生于潮间砂坪环境; ②Palaeophycus tubularis遗迹组构, 主要发育在暗紫红色泥质粉砂岩和粉砂质泥岩中, 组成分子包括Cochlichnus, Helminithopsis, Palaeophycus 和Protovirgularia, 总体上反映了水深较浅、水面变化比较频繁的潮间泥坪环境。此外, 陆廷清等(2012)在龙门洞一带嘉一段中还发现鱼类游泳遗迹— — Undichna unisulca(单槽波形迹), 是中国发现时代最早的脊椎动物遗迹化石之一, 它的形成环境属于河口湾的混积潮坪环境。四川广安谢家槽一带下三叠统夜郎组和嘉陵江组中已识别的遗迹化石有Arenicolites, Chondrites, Didymaulichnus, Monocraterion, Paleophycus, Planolites, Skolithos, Taenidium, Teichichnus, Thalassinoides, Treptichnus, Uchirites等, 嘉陵江组薄层泥晶灰岩、泥粒状灰岩和条带状灰岩中发育Arenicolites, Isopodichnus, Paleophycus, Phycodes, Planolites, Psammichnites, Thalassinoides和Skolithos等, 这些遗迹化石形成于局限台地相和开阔台地相沉积背景(周志澄等, 2014)。
在四川盆地上三叠统须家河组中发育大量遗迹化石, 经金惠等(2013)研究, 已鉴定出3大类7属10种, 可建立3种遗迹组合: ① Cochlichnus-Planolites 遗迹组合, 主要由进食迹和牧食迹组成, 常见有 Cochlichnus anguineus, Planolites beverleyensis 和 Palaeophycus tubularis, 形成于潮间泥坪沉积环境; ② Cylindricum-Planolites 遗迹组合, 以居住潜穴和进食潜穴为主, 常见有 Cylindricum isp., Skolithos linearis, Skolithos verticulis, Ophiomorpha nodosa, Planolites beverleyensis, P. montanus, Palaeophycus tubularis 和 P.striatus等, 丰度和分异度高, 代表了潮间带混合坪沉积环境; ③ Skolithos linearis 遗迹组合, 由长的垂直或高角度倾斜的悬食居住潜穴构成, 常见有Skolithos linearis, Arenicolites isp.和Planolites montanus, 形成于潮间砂坪和水下分流河道沉积环境, 环境水体能量随着相对海平面的变化而发生周期性的变化。值得注意的是, 在四川盆地以西的藏北羌塘盆地东部冬曲地区上三叠统巴贡组下段发现了Paleodictyon(古网迹), 其属于深海、贫氧、高盐度的复理石沉积环境(占王忠等, 2019)。
扬子台地西南缘贵阳花溪地区早三叠世发育一套浅水碳酸盐岩台地— 深水碎屑岩盆地过渡相沉积, 自下而上为大冶组和安顺组。大冶组主要为浅灰色薄层石灰岩与薄板状泥质石灰岩, 富产遗迹化石, 已鉴定出18个遗迹属, 自下而上已识别出3种遗迹组构, 分别为: ① Phycodes遗迹组构, 主要保存于大冶组第一段上部薄层泥质灰岩夹层中, 以Phycodes circinatus的大量出现为主要特征, 伴随有少量的Planolites beverleyensis, 产生于浅海台地相沉积中; ② Phycosiphon-Mammillichnis遗迹组合, 发育于大冶组第二段中部的薄板状灰岩与泥质灰岩中, 以Phycosiphon 的密集分布以及Mammillichnis, Cosmorhaphe, Gordia, Glockerichnus, Megagrapton, Planolites 和Palaeophycus的大量出现为特征, 形成于台缘下斜坡浊流沉积中; ③Thalassinoidess遗迹组构, 发育在大冶组第三段灰色中厚层泥晶灰岩中, 常见Thalassinoides和Phycodes, 产于潮间带滨海环境(时国等, 2009)。安顺组中识别出的遗迹化石有8个遗迹属和9个遗迹种: 一段中发现的遗迹化石主要为Planolites montanus, Phycodes palmatus和 Palaeophycus curvatus 等, 解释为台缘浅滩相沉积; 二段中已识别的遗迹化石有Planolites montanus, Palaeophycus curvatus, Phycodes palmate, Beaconichnus darwinum, Thalassinoides paradoxicus, Chondrites recurvus和Planolites beverleyensis 等, 解释其形成于台缘斜坡沉积环境; 三段中鉴定的遗迹化石有 Planolites beverleyensis, Phycodes palmatus, Rhizocorallium commune, Arenicolites isp.和 Thalassinoides pradoxicus等, 属于靠近台地边缘的底内生物繁盛的潮间— 潮下沉积环境; 四段由紫红色中至厚层状白云岩构成, 时夹盐溶角砾岩, 发育大量次生孔洞和鸟眼构造, 偶见生物扰动构造, 未见形态完整的遗迹化石, 为半局限台地内的潮间— 潮上带沉积环境(余关美和时国, 2016)。
Luo等(2019)在中国西南地区云南省罗平县中三叠统下部关岭组碳酸盐岩中识别出14个遗迹属, 均产于碳酸盐岩台地环境, 其中有9个遗迹属出现在开阔潮下浅海环境; 尤其是各种甲壳动物的遗迹、海生爬行动物的运动迹同丰富的鱼类和十足类动物化石一起出现, 更突显遗迹化石在P-Tr大灭绝后生态系统恢复的价值。
华南中扬子区湖北宜昌三峡一带下三叠统大冶组中— 上部及嘉陵江组中部浅海灰岩中的遗迹化石已识别出14个遗迹属和17个遗迹种, 包括Arenicolites isp., Chondrites filiformis, Circulichnus montanus, Cochlichnus kochi, Didymaulichnus lyelli, Diplocraterion parallelum, Diplocraterion isp., Gordia molassica, Mammillichnis?isp., Oldhamia radiata, Oldhamia isp., Palaeophycus heberti, Palaeophycus tubularis, Paleodictyon isp., Planolites montanus, Skolithos isp., Thalassinoides isp.等。Zhao等(2015)描述和分析了这些遗迹化石的丰度、分异度、生物扰动指数、潜穴大小和阶层, 并依据这些特征评价了华南中扬子区P-Tr大灭绝后生态系统复苏速度和形式, 同时还将最新获得的地球化学数据与理想的复苏模式相结合, 解释了早三叠世遗迹化石组合的时空分布特征。
在下扬子地区, 赵小明和童金南(2010)通过对浙江长兴煤山二叠系— 三叠系界线全球层型剖面附近钻孔岩心的研究, 识别出遗迹化石Chondrites, Palaeophycus, Planolites, Rhizocorallium, Skolithos和Thalassinoides等。研究表明, 遗迹化石的丰度、扰动指数及扰动深度等遗迹学指标在二叠系— 三叠系界线上下呈现两幕式剧烈变化, 这说明二叠纪— 三叠纪之交遗迹化石的演变与实体化石的演变有着相似的幕式变化过程与趋势。
Chen等(2011)研究了下扬子地区的皖南、苏南、赣北和浙西一带早三叠世海相地层中丰富的遗迹化石, 计有16个遗迹属。在下三叠统下部殷坑组绿色或黑色页岩夹泥灰岩或灰岩中仅发现Planolites, 属风暴浪基面以下滨外浅海远端斜波较深水缺氧环境; 在下三叠统中部和龙山组薄层白云质灰岩夹钙质泥岩中识别出Arenicolites, Cochlichnus, Gordia, Gyrochorte, Kouphyichnium, Palaeophycus, Planolites和Treptichnus等遗迹化石, 解释形成于正常天气与风暴浪基面之间的浅海上部环境; 在下三叠统上部南陵湖组中厚层状灰岩夹泥岩中, 遗迹化石最发育, 已识别出12种遗迹属, 被认为产生于临滨到浅海上部或浅海碳酸盐岩台地环境。根据上述遗迹化石的垂向分布特征, 早三叠世早期仅发现1种遗迹属, 表明下扬子区当时较深水浅海环境中后生底栖动物甚少, 到中期出现8种遗迹属, 说明此时近岸浅海环境中后生底栖动物大量复苏, 最后到晚期在临滨至浅海上部环境中发育12种遗迹属, 显然后生底栖动物已繁盛起来。由此可见, 这些在二叠纪末大灭绝之后逐渐增多的遗迹化石是二叠纪末生物危机后生态恢复的有力见证。
张立军等(2015)总结了晚二叠世末大灭绝事件P/Tr界线上下遗迹化石的时空分布特征, 认为主要具有以下3个方面的变化规律: (1)遗迹化石丰度和多样性变化趋势表现为低纬度地区, 遗迹属数量在二叠纪末期呈现出低水平, 早三叠世有所增加, 一直持续到奥伦尼克期, 并在奥伦尼克期晚期达到高峰, 因此中三叠世的遗迹属数量比二叠纪末期丰富; 在高纬度地区, 二叠纪末期遗迹属个体呈现一定的分异度, 且多为深水型分子, 早三叠世早期遗迹属数量急剧减少到最低点, 印度晚期遗迹属数量有所增加, 开始出现了浅水型分子, 至奥伦尼克期和安尼期遗迹属数量达到了顶峰。早三叠世印度期浅海区生物多样性复苏较慢, 印度晚期至奥伦尼克期, 滨海区机会主义遗迹属复苏较快, 深水区遗迹属复苏较慢。(2)遗迹潜穴直径变化趋势与P-Tr大灭绝事件之后海洋底栖生物中普遍的小型化事件相对应, P-Tr大灭绝事件前后遗迹化石潜穴直径也有明显的变化。(3)遗迹化石行为习性变化趋势表现为少数遗迹属(如Planolites和 Arenicolites)能反映造迹生物迅速占据生态坍塌后的裸地, 而一些三维复杂潜穴(如Thalassinoides)和滤食性生物觅食迹(如 Rhizocorallium)则反映了造迹生物垂向分布和生态系统恢复到较高的程度。
丁奕(2018)在浙江煤山剖面乐平统吴家坪阶龙潭组上部至下三叠统印度阶殷坑组下部的连续地层中识别出遗迹化石9个遗迹属和10个遗迹种, 以及其他一些掘穴与逃逸构造, 并把这些遗迹化石归入Skolithos遗迹相。研究认为, 这些遗迹化石不仅能反映古氧相, 而且由于其具有较高的丰度与较低的分异度, 极有可能反映P-Tr事件前后遗迹化石及其造迹生物生态系统的变化。
综上所述, 中国P/Tr界线附近的遗迹化石在扬子区的海相地层中研究程度较高, 可为晚二叠世末生物大灭绝后的生态系统复苏过程提供可靠的遗迹学信息(赵小明和童金南, 2010; Chen et al., 2011; 张立军等, 2015)。
中国陆相遗迹化石大多分布在中生代和新生代陆相沉积盆地中, 近十年来的研究地区主要包括豫西众多中生代盆地、渤海湾新生代盆地、鄂尔多斯盆地、柴达木盆地、四川盆地和苏北盆地等。据不完全统计, 在这些盆地中已发现无脊椎动物遗迹化石共计44个遗迹属和107个遗迹种, 另外还有四足动物足迹和植物根迹。这些无脊椎动物遗迹化石包括居住迹、爬行迹、停息迹、进食迹、觅食迹和耕作迹(Agrichnia)等生态类型, 大多产于冲积扇、辫状河、曲流河及湖泊沉积环境, 可归属于Scoyenia, Mermia和Skolithos等3种遗迹相。基于遗迹化石组成和分布特征, 建立了22种遗迹化石组合和10种遗迹组构, 为陆相沉积环境的精细分析和陆相油气资源勘探开发提供了重要的生物遗迹学信息。
国内有关冲积扇环境中的遗迹化石研究报道较少。杨凯等(2013)、Hu等(2014a)在豫西汝阳盆地白垩系(下河东组、郝岭组和上东沟组)冲积扇沉积(砂质砾岩与棕红色泥质粉砂岩或砂质泥岩互层)中发现遗迹化石7个遗迹属和8个遗迹种, 以及植物根迹和脊椎动物足迹化石等。基于遗迹化石组成、分布及其宿主岩石的沉积特征, 建立了2种遗迹组合: ① Scoyenia-Skolithos 遗迹组合, 主要由Scoyenia, Skolithos, Planolites和Palaeophycus 及根迹组成, 出现在冲积扇沉积体系的辫状河沉积环境; ② Psilonichnus-Palaeophycus 遗迹组合, 以丰富的Psilonichnus, Palaeophycus 和 Arenicolites为特征, 含少量Skolithos和 Palaeophycus, 产于冲积扇沉积体系的中扇前缘小型浅水湖泊环境。
河流环境中的遗迹化石研究以往已有大量报道, 近几年在豫西中生代(三叠纪、白垩纪)盆地、鄂尔多斯(三叠纪)盆地和苏北(古近纪)盆地又有一些新的研究进展。
Wang等(2014)在豫西济源盆地中三叠统油坊庄组河流沉积中识别出7种遗迹组构(图 9): ①Palaeophycus tubularis 遗迹组构, 仅以Palaeophycus tubularis 的大量出现为特征, 生物扰动指数(BI)为2, 保存在河道砂岩的底面, 位于2个河流沉积旋回之间; ② Skolithos linearis遗迹组构, 仅由单一潜穴Skolithos linearis构成, 形成于半干旱气候条件下河道沉积序列的中— 上部沉积中; ③ Skolithos linearis-Planolites montanus遗迹组构, 以层内垂直— 近垂直的Skolithos linearis与水平潜穴Planolites montanus, Planolites beverleyensis, Palaeophycus striatus 和Gordia isp.等遗迹化石为特征, 生物扰动强度中等(BI=3~4), 其宿主岩为棕红色砂岩, 保存于半干旱气候条件下河道沉积序列的上部沉积中; ④ Planolites montanus 遗迹组构, 常见遗迹化石包括Planolites montanus, Palaeophycus tubularis, Skolithos isp.和 Planolites beverleyensis等, 生物扰动强度中— 高(BI=3~5), 大多保存在棕红色中— 厚层状的粉砂质泥岩或粉砂岩中, 形成于河漫滩上浅水湖泊沉积环境; ⑤ Taenidium barrette 遗迹组构, 以具回填纹的Taenidium barretti潜穴为特征, 生物扰动强度中— 高(BI=3~5), 产自具水平和波状纹理的浅棕色或灰绿色粉砂岩或粉砂质泥岩中, 形成于周期性暴露的短期水池或极浅水湖泊或低能远端泛滥平原环境; ⑥ Skolithos isp. 遗迹组构, 仅由单一潜穴Skolithos isp.构成, 出现在浅棕色或浅灰绿色薄层细砂岩或粉砂岩中, 形成于近源泛滥平原或决口扇中— 高能的固底沉积物中; ⑦ Root Trace(根迹)遗迹组合, 根迹轮廓模糊且不连续, 直径0.4~0.5 cm, 长度1~6 cm, 生物扰动程度低(BI=1~2), 常发育在浅灰绿色或浅棕色细砂岩或粉砂岩中, 被解释为决口扇沉积环境。
 | 图 9 豫西济源盆地中三叠统油坊庄组的遗迹组构类型及其沉积环境分布模式(据Wang et al., 2014)Fig.9 Sedimentary environment model of ichnofabric types of the middle Triassic Youfangzhuang Formation in Jiyuan Basin, western Henan Province(after Wang et al., 2014) |
在鄂尔多斯盆地东南部石川河一带的中三叠统河流相沉积中也识别出6个遗迹属和9个遗迹种, 包括Gordia indianaensis, Palaeophycus tubularis, Palaeophycus isp., Planolites beverleyensis, Planolites isp., Scoyenia gracilis, Skolithos linearis, Skolithos isp., Taenidium isp.及根迹等(Guo et al., 2019)。
豫西南西峡盆地上白垩统高沟组、马家村组及寺沟组主要为一套紫红色泥岩、泥质粉砂岩、粉砂岩、浅灰色细砂岩, 间夹浅灰色、灰白色中— 粗粒砂岩或砾岩, 含有丰富的遗迹化石, 已鉴定出10个遗迹属和22个遗迹种(Wang et al., 2016)。基于遗迹化石的组成、分布及其沉积特征, 建立了 4种遗迹组合: ① Skolithos-Gastruichnus 遗迹组合, 产生于辫状河、曲流河的河道沉积环境; ② Beaconites-Taenidium遗迹组合, 出现在河流天然堤和决口扇沉积环境; ③ Beaconites-Palaeophycus 遗迹组合, 发育于泛滥平原上曲流河河道及河漫滩沉积环境; ④Taenidium-Scoyenia 遗迹组合, 常见于周期性暴露的河漫湖泊(凹坑或极浅水湖泊)沉积环境。对寺沟组中的遗迹化石做进一步精细研究后, 在Beaconites-Palaeophycus遗迹组合中识别出10个遗迹属和17个遗迹种(王长征等, 2016)。经过综合分析这些遗迹化石的产状及其沉积特征, 提出了代表曲流河河道及河漫滩沉积环境中遗迹化石的产状及分布模式。
苏北盆地古新世河流沉积中发现遗迹化石5个遗迹属和8个遗迹种, 包括具新月形回填纹潜穴、 简单水平柱状潜穴、简单垂直或近垂直管状潜穴及植物根迹等。其中, 河道沉积中产有Skolithos, Taenidium 和 Beaconites; 决口扇沉积中常见Palaeophycus, Planolites, Taenidium 和 Skolithos; 远端泛滥平原泥质为主的沉积中发育丰富的Taenidium, Beaconites, Palaeophycus, Planolites和Skolithos以及其他甲壳动物和蠕虫类动物的居住潜穴(Zhou et al., 2019)。
湖泊环境中的遗迹化石研究在河南中新生代盆地、鄂尔多斯中生代盆地、新疆柴达木盆地、甘肃宝积山盆地及重庆万州地区都取得了许多新进展。在这些地区的三叠纪、侏罗纪、白垩纪、古近纪和新近纪地层中已识别遗迹化石计有48个遗迹属和85个遗迹种, 建立了16种遗迹组合及3种遗迹组构。
豫西济源地区下三叠统和尚沟组滨浅湖沉积中共识别出9个遗迹属和10个遗迹种, 可划分为3种遗迹组构: ① Scoyenia遗迹组构, 反映了干旱气候条件下的滨湖沉积环境; ② Planolites-Taenidium遗迹组构, 属于浅湖沉积环境; ③ Psilonichnus遗迹组构, 代表了水动力由弱到强的湖泊三角洲沉积环境(胡斌等, 2009)。豫西登封地区下三叠统刘家沟组及和尚沟组的湖相沉积中发现丰富的遗迹化石, 根据遗迹化石的组成、产状和分布特征, 建立了2种湖相遗迹组合: ① Arenicolites遗迹组合, 发育在砂泥交互沉积中, 主要以Arenicolites和Psilonichnus大量发育为特征, 其次为Skolithos和Beaconites等, 分异度中等, 丰度相对较高, 形成于湖泊环境中近岸水流较动荡的滨湖沉积环境; ② Taenidium-Planolites遗迹组合, 主要发育在和尚沟组的粉砂岩和砂质泥岩中, 以Taenidium和Planolites的大量出现为特征, 同时也常伴生出现一些Beaconites, Palaeophycus和Scoyenia等遗迹化石, 分异度中等, 但丰度往往较高, 产生于水体较平静的浅湖沉积环境(胡斌等, 2015)。豫西南留山盆地上三叠统太山庙组中识别出Beaconites, Cochlichnus, Palaeophycus和Planolites等遗迹化石, 主要产自浅湖沉积环境(钟明洋等, 2010)。在豫西南栾川潭头盆地上白垩统秋扒组发现多层丰富的遗迹化石, 按其组成和分布特征, 建立了2种遗迹组合(图 10): ① Palaeophycus-Rhizoliths遗迹组合, 主要组成分子有Palaeophycus, Planolites, Beaconites和根迹, 遗迹分异度和丰度较高, 形成于山间断陷湖盆的扇三角洲平原— 扇三角洲前缘沉积环境; ②Beaconites-Scoyenia遗迹组合, 主要发育Beaconites, Scoyenia, Planolites和Skolithos等, 遗迹分异度较高, 丰度相对较低, 产生于周期性出露地表的滨、浅湖沉积环境(王长征等, 2014)。在西峡盆地上白垩统寺沟组中见有Taenidium, Beaconites, Scoyenia, Palaeophycus和Planolites等遗迹化石, 组成Taenidium-Beaconites遗迹组合, 产生于滨浅湖沉积环境(王长征等, 2016)。
 | 图 10 豫西潭头盆地上白垩统中遗迹化石组合及其沉积环境模式(据王长征等, 2014) A— 扇三角洲平原— 扇三角洲前缘沉积特征及遗迹化石组合; B— 滨浅湖沉积序列及遗迹化石组合Fig.10 Ichnofossil assemblages and their sedimentary environment model of the upper Cretaceous in Tantou Basin of western Henan Province(after Wang et al., 2014) |
近几年在东濮凹陷古近系沙河街组湖相碎屑岩中新发现大量遗迹化石, 已鉴定16个遗迹属和24个遗迹种。根据这些遗迹化石的组成和分布特征, 建立了4种不同湖相环境的遗迹组合(常龙等, 2017; 胡斌等, 2017): ① 出现于浅湖或湖泊三角洲前缘的Palaeophycus-Planolites遗迹组合; ② 发育于较深浅湖— 半深湖环境的Planolites-Helminthoidichnites或Mermoides遗迹组合; ③ 产生于较深湖或前三角洲的Helminthopsis-Paleodictyon遗迹组合; ④ 代表了较深湖浊积扇环境的Helminthopsis-Trichichnus遗迹组合。值得注意的是, 张国成等(2013)专门论述了东濮凹陷沙河街组湖相浊积岩中的Paleodictyon(古网迹)形态与产状特征, 并与海相同类化石进行了对比分析, 认为该遗迹属可作为东濮凹陷乃至渤海湾盆地在古近纪存在海侵的重要证据。
Guo等(2019)最近在鄂尔多斯盆地东南部石川河一带晚二叠世(孙家沟组)至早三叠世湖相地层(刘家沟组、和尚沟组)中发现较多的遗迹化石, 系统地研究了16个遗迹属。研究表明, 二叠纪末生物大灭绝后, 大陆生态系统发生全面复苏应该在早三叠世晚期至中三叠世, 显示高遗迹分异度、强生物扰动和造迹生物行为复杂性等特征。值得关注的是, 在石川河及柳林地区上二叠统孙家沟组上部、早三叠世刘家沟组上部及和尚沟组中— 上部发现44块保存良好的Kouphichnium(一种典型的鲎类足辙迹)标本, 鉴定有6个遗迹种。遗迹化石的组成和分布特征表明晚二叠世至早三叠世生物遗迹的急剧减少及之后逐渐增加的特点(Shu et al., 2018)。
除了上述湖相沉积中的遗迹化石外, 张锋等(2018)在重庆万州区铁峰山下侏罗统自流井组大安寨段湖相沉积中还发现高丰度的Palaeophycus tiefengshanensis(铁峰山古藻迹), 分布于页岩与介壳灰岩界面上, 即介壳灰岩的底面。该种遗迹化石为四川盆地东部地区自流井组沉积期古环境分析提供了重要生物遗迹学信息。刘松等(2017)在甘肃宝积山盆地上三叠统南营儿组发现了石针迹属(Skolithos)中的垂直石针迹(Skolithos verticalis)和线状石针迹(Skolithos linearis), 产自滨湖沉积环境。高正海等(2013)在柴达木盆地东北缘双湖地区中新统灰绿色、灰色砂质岩与棕红色泥质岩互层为主的岩层中发现了Palaeophycus, Skolithos, Scoyenia和Taenidium等遗迹化石。根据这些遗迹化石的分布特点及其沉积环境分析, 建立了 Scoyenia-Taenidium-Skolithos遗迹组合, 代表了潮湿气候条件下的浅湖环境。这一发现填补了柴达木盆地东北缘双湖地区中新世遗迹化石研究的空白。
总的来说, 中国陆相湖泊沉积环境中的遗迹化石是十分丰富的, Hu等(2014b)总结了中国多地陆相湖泊环境中遗迹化石组成特征及分布规律, 划分出6种遗迹组合(图 11): ① Scoyenia-Skolithos遗迹组合, 主要由Scoyenia, Beaconites, Taenidium, Planolites, Skolithos, Thalassinoides, Palaeophycus, Cystichnium, Gordia, Sagittichnus, Favreina 和根迹组成, 产生于干旱或半干旱气候条件下周期性暴露的极浅水湖的滨湖或湖泊三角洲分流间湾沉积环境; ② Palaeophycus-Arenicolites遗迹组合, 主要组成分子有Palaeophycus, Arenicolites, Skolithos, Cylindricum, Polykladichnus, Thalassinoides, Ophiomorpha, Diplocraterion, Trichichnus, Tigillites, Beaconites, Ancorichnus和 Planolites等, 发育于滨湖至浅湖上部沉积环境; ③Planolites-Teichichnus 遗迹组合, 以密集分布的小型Planolites为特征, 伴生有Gordia, Helminthopsis, Brevitubus, Cruziana, Palaeophycus, Arenicolites, Sagittichnus 和Margaritichnus及少量Chondrites, 常见于浅湖下部、局限湖湾和三角洲前缘沉积环境; ④Vagorichnus-Helminthopsis遗迹组合, 代表性遗迹化石包括Cochlichnus, Helminthoidichnites, Helminthopsis, Monomorphichnus, Neonereites, Paracanthorhaphe, Tuberculichnus和 Vagorichnus等, 产于较深水湖浊流沉积环境; ⑤Mermoides-Neonereites 遗迹组合, 由Mermoides, Pilichnus, Neonereites, Helminthoidichnites, Helminthopsis, Vagorichnus, Multilagueichnus, Chondrites, Cochlichnus, Gordia, Planolites 和 Favreina等组成, 形成于平静的深湖沉积环境; ⑥ Semirotundichnus-Chondrites 遗迹组合, 典型的遗迹化石有Semirotundichnus, Patellarisichnus, Neonereites, Circinansichnus, Chondrites和 Planolites等, 出现在平静且含氧量低或贫氧的深湖沉积环境。
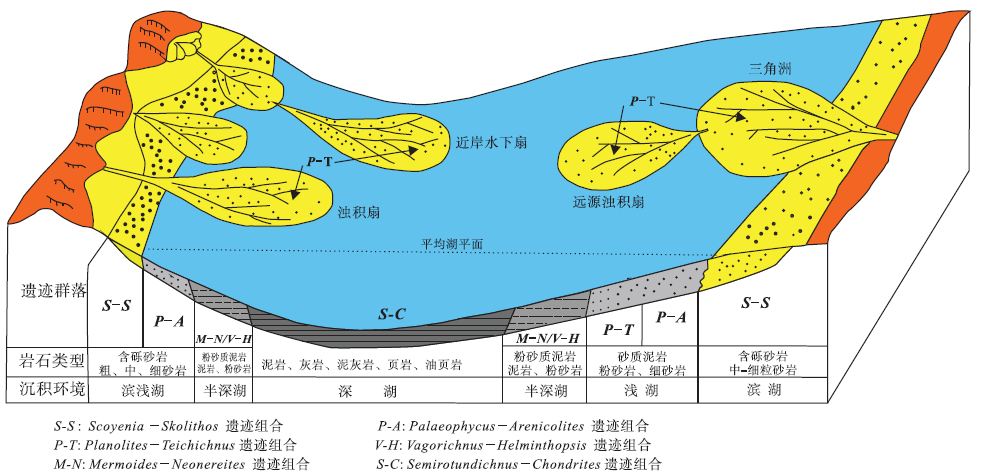 | 图 11 湖泊沉积环境中遗迹化石组合分布模式(据Hu et al., 2014b)Fig.11 Distribution model of ichnoassemblages in lacustrine sedimentary environments(from Hu et al., 2014b) |
中国陆相沉积中的恐龙足迹研究在近十年取得了许多重要成果, 研究区及地层包括:山东胶莱盆地西南部诸城、临沭、郯城及江苏东海的下白垩统; 华北地区张家口、北京延庆、内蒙古乌拉特中旗侏罗系和下白垩统; 西北地区宁夏六盘山固原一带下白垩统, 鄂尔多斯盆地东北部陕西神木地区的白垩系; 四川攀西、峨眉及重庆綦江地区的白垩系。
山东胶莱盆地恐龙足迹化石十分发育, 在下白垩统莱阳群和青山群浅紫色砂岩— 粉砂岩、泥岩层面上保存了大量的兽脚类、蜥脚类和鸟脚类恐龙足迹化石群(柳永清等, 2011)。对足迹特征的分析显示: 蜥脚类恐龙体重大, 四足行走, 前后足迹面积相近, 似椭圆形; 鸟脚类和兽脚类恐龙为两足行走, 足迹为三趾型; 兽脚类恐龙体重最轻、数量最多(李素等, 2012)。何碧竹等(2012)研究了山东诸城早白垩世莱阳期古地震事件与恐龙迁移的关系, 认为区域构造作用使得该时期沉积环境发生变化, 由湖相逐渐过渡到河流相, 在突发古地震事件发生之后, 在滨、浅湖区域留下了丰富的恐龙迁徙足迹。对山东临沭地区早白垩世晚期生活于干旱炎热气候条件下滨、浅湖环境的恐龙所留下的大规模足迹化石群行为学的分析表明, 蜥脚类和鸟脚类恐龙行迹均处于慢行状态, 且鸟脚类恐龙行迹的运动速度极慢(陈军等, 2013)。从恐龙足迹保存特征及其沉积背景来看, 山东沂沭断裂带南部地区(莒南、临沭、郯城和东海)下白垩统中大量的恐龙足迹化石所反映的足迹类型、形态大小、足迹踩踏深度等都存在明显差异(彭楠等, 2013)。山东沭河裂谷带早白垩世晚期恐龙足迹群的恐龙足迹一般为凹形印痕, 造迹者以蜥脚类恐龙为主, 其次为鸟脚类和兽脚类恐龙。足迹和行迹特征显示, 造迹恐龙为群居和在栖息地呈现漫步行走状态。足迹原位保存特征及赋存地层沉积学研究表明, 早白垩世中晚期气候处于由温暖潮湿变得炎热干旱的过渡期, 湖泊、三角洲广布, 恐龙栖息于湖滨环境(旷红伟等, 2013)。恐龙足迹化石多产在砂质滩坝微相细砂岩、粉砂岩层面上, 这说明河湖相细砂岩、粉砂岩有利于恐龙足迹化石赋存, 也暗示蜥脚类恐龙喜好在湖岸边活动, 生活习性与生存环境具有一定的相互制约关系(王宝红等, 2013)。研究还发现蜥脚类与兽脚类恐龙的足迹大小差异较大。在同等条件下, 蜥脚类恐龙足迹通常较兽脚类足迹深, 且蜥脚类恐龙足迹深度随着足迹大小的增加有变深的趋势; 波痕、雨痕和泥裂等沉积构造以及足迹回填和叠覆现象对足迹保存形态和细节有较大影响, 而构造变形对足迹形态和行迹并无明显影响, 但高角度的构造变形会加速差异风化, 使足迹化石更易暴露而被破坏(汪明伟等, 2013)。对山东诸城黄龙沟产于下白垩统中部莱阳群杨家庄组兽脚类与蜥脚类、疑似鸟脚类足迹数量之比远大于正常捕食者和被捕食者之比的研究表明, 早白垩世中期的胶莱盆地生存着以兽脚类为主、蜥脚类和鸟脚类为辅的恐龙动物群。早白垩世晚期, 胶莱盆地以及沂沭断裂带内蜥脚类、鸟脚类恐龙以及鸟类逐渐繁盛, 这与依据华北北部恐龙足迹所恢复的造迹恐龙动物群特征较为一致(许欢等, 2013)。
在华北内蒙古乌拉特中旗一带侏罗纪早期地层中, 李建军等(2010)识别并详细描述了3种恐龙足迹: ①玫瑰实雷龙足迹(Eubrontes glenrosensis), 为大型三趾型足迹, 两足行走, 属于大型兽脚类恐龙足迹; ②海流图卡岩塔足迹(Kayentapus hailiutuensis), 为中型三趾型兽脚类恐龙足迹; ③中型异样龙足迹(Anomoepus intermedius), 为小型四足行走足迹, 前足五指、后足四趾, 3个功能趾, 属于小型鸟脚类恐龙足迹。伴生的其他动物足迹及鳄类足迹的蛙步足迹, 均产自山间河流沉积环境。在冀西北张家口尚义地区晚侏罗世— 早白垩世地层中也发现大量恐龙足迹。恐龙行迹呈近东西向展布, 由西向东行进, 由数十个足迹构成。研究显示, 足迹中包括兽脚类足迹70余个, 组成数个行迹; 蜥脚类足迹15个, 构成1列行迹, 显示其可能属于游泳状的行迹(柳永清等, 2012)。据何情等(2015)报道, 在北京延庆千家店地区上侏罗统土城子组砂岩和凝灰岩中也发现以兽脚类和蜥脚类为主的恐龙足迹, 另外可能还有鸟脚类恐龙足迹。综合分析认为, 当时的沉积环境为含火山碎屑的浅湖, 恐龙足迹的赋存层位为滨湖沉积。
西北地区宁夏六盘山固原市隆德县和泾源县一带下白垩统六盘山群和尚铺组、李洼峡组和马东山组中保存有蜥脚类和鸟脚类恐龙足迹化石, 足迹可识别的有4条行迹, 其中以蜥脚类为主, 根据岩性及沉积构造判断, 造迹恐龙活动环境为离湖岸较近的浅水地带(杨卿等, 2019)。其中, 李洼峡组滨湖相沉积中的鸟脚类恐龙足迹命名为六盘山宁夏足印(Niugxiapus liupanshauensis)(宗立一等, 2013)。鄂尔多斯盆地东北部陕西神木地区下白垩统洛河组中部发现二趾型恐龙足迹3处, 小型四足动物足迹2处, 根据足迹形态对比, 推断为兽脚类实雷龙类、恐爪龙类足迹, 小型四足动物足迹为哺乳动物足迹类, 恐龙足迹分布在水体较浅的滨湖环境(唐永忠等, 2018)。
中国西南地区四川盆地及周边地区的中侏罗世和白垩纪地层中也发现多枚恐龙足迹化石。鄢圣武等(2017)在四川西昌盆地边缘的四川美姑县中侏罗统沙溪庙组底部薄层粉砂岩层面上发现3枚恐龙足迹化石, 均为三趾型兽脚类, 形态特征与经典的小型兽脚类跷脚龙足迹(Grallator)相似。四川峨眉地区上白垩统夹关组中含有多种恐龙足迹, 陆廷清等(2013)在该地白垩系夹关组中部的棕红色细、粉砂岩中新发现斧头状、盘状和碗形等3种类型的足迹, 进一步丰富了该地区上白垩统恐龙足迹形态和保存类型。在重庆綦江区虎山白垩系夹关组中发现了2个大型恐龙天然足模化石, 化石保存在块状砂岩夹薄层泥岩的悬崖底面, 呈现出放射状、同心状或菜花一样的形态, 属于罕见的石花形恐龙幻迹, 同类足迹也发现于韩国和北美白垩系(邢立达等, 2015)。重庆綦江莲花保寨一带夹关组中保存有大量以莲花卡利尔足迹(Caririchnium lotus)为主的恐龙足迹群, 被解释为产生于曲流河边滩或河漫滩环境(代辉等, 2016)。四川攀西地区米市— 江舟盆地白垩系飞天山组、小坝组和雷打树组中发现了9个足迹点及9种不同的非鸟恐龙足迹(6种非鸟兽脚类、1种蜥脚类和2种鸟脚类足迹), 以及翼龙和龟类足迹, 共11种足迹的形态类型由185道行迹(与孤立足迹)组成, 其中Dromaeopodus-Velocaraptorachnus-Manasaurapus组合显示出强烈的东亚土著特色(曹俊等, 2016)。
综上所述, 中国近10年发现的恐龙足迹类型主要包括兽脚类、蜥脚类和鸟脚类足迹等。邢立达等(2018)总结了中国白垩纪的恐爪龙类足迹(两趾型足迹), 这是特征最为鲜明的兽脚类足迹之一。在中国发现的14个足迹点中多数足迹都被归入5个足迹属: Velociraptorichnus, Dromaeopodus, Menglongipus, Dromaeosauripus和Sarmientichnus。其中, Dromaeosauripus包括3个足迹种, 其他足迹属仅包括1个足迹种。李建军(2015)、邢立达等(2018)对新发现的恐龙和鸟类足迹化石进行了详细描述与对比, 并对已报道的足迹属、种进行了重新厘定。据李建军的统计, 中国已发现的中生代爬行动物和鸟类足迹化石共计58属77种, 其中恐龙足迹有43属60种(王原等, 2019)。除了恐龙分类学研究外, 许多学者还运用地层学、岩石学、古生物学和沉积学方法, 综合分析了足迹化石产出地层的沉积环境。研究认为, 大多数足迹都产自河流边滩、河漫滩或冲积平原和滨湖沉积环境(柳永清等, 2011; 旷红伟等, 2013; 何情等, 2015; 代辉等, 2016; 杨卿等, 2019)。此外, 大量足迹的发现及其研究为恐龙类和鸟类古生态学普查提供了动物群的组成信息, 为更大的时空范围内进行横向与纵向的地层对比提供了科学依据。如在四川、宁夏等地区运用恐龙足迹学可有效、快速地判定地层中动物群的构成, 并作为骨骼化石的重要补充进行了类似的生物地层对比(宗立一等, 2013; 曹俊等, 2016)。
遗迹学主要研究生物与沉积物的相互作用, 一般而言, 造迹生物的活动大多在沉积事件的沉积间歇期进行。因此, 遗迹学可揭示事件间歇期的沉积特征、持续时限与过程。此外, 沉积学研究还表明, 沉积作用常有快速的、间歇性的和周期性的突发事件所留下的地层记录, 其比渐进式的沉积作用留下的记录更重要, 因为它更能揭示沉积过程中各种各样的变化, 而遗迹学对研究沉积间断面上的信息具有独特优势。因此, 遗迹学在事件沉积判识或沉积间断面成因分析等方面具有重要的应用价值。
吴贤涛等(2013)对常见事件沉积的环境位置、沉积学和遗迹学标志以及沉积间断时限推测进行了归纳和总结, 指出了事件间歇期的遗迹学标志和代表性的遗迹化石。近十年来, 中国学者对遗迹学在事件沉积方面的应用研究重点集中在2个方面: (1)遗迹学在风暴沉积研究中的应用; (2)遗迹学在浊流沉积中的应用。
6.1.1 风暴沉积中的遗迹化石组成与分布特征
中国近几年在豫西(鲁山、登封、洛阳龙门)和山东中部(淄博、峨庄、莱芜)地区的寒武系、四川龙门山地区的泥盆系、豫西北及晋东南地区的二叠系及四川广安地区的三叠系等地层中开展了风暴沉积与遗迹化石的研究工作, 取得较多新成果。
河南鲁山寒武系第二统辛集组为一套风暴作用影响下的碎屑岩沉积。通过对风暴沉积序列与遗迹化石组成及分布特征的分析, 显示遗迹化石出现在风暴衰减期和风暴间歇期的沉积中, 前者仅出现单一的 Skolithos, 它以短小的垂直居住潜穴及低遗迹分异度和丰度为特征, 后者由多种遗迹化石组成, 包括Palaeophycus, Taenidium, Gordia, Planolites等, 属于Cruziana 遗迹相中的遗迹化石, 以近层面分布的进食、觅食潜穴为主, 遗迹分异度和丰度中等。据遗迹化石的保存特征分析可知, 在早寒武世风暴衰减期, 频繁的风暴事件不利于遗迹化石的保存, 同时风暴间歇期持续时间短, 亦不利于对沉积底质的连续殖居。这种特殊的风暴沉积以及早寒武世造迹生物较差的掘穴能力, 导致这一时期遗迹化石种类及数量远不及寒武纪以后的风暴沉积(白万备等, 2018)。豫西登封地区寒武系第三统馒头组三段发育一套含大量生物成因构造(叠层石和各类垂直潜穴)的碳酸盐岩风暴沉积, 受风暴沉积作用影响的鲕粒灰岩中出现大量滤食动物的垂直或近垂直居住潜穴, 以直管状潜穴 Skolithos为主, 次为U形管状潜穴Arenicolites(齐永安等, 2014b), 这些遗迹化石一般都是较高水动力条件的产物。洛阳龙门地区寒武系第三统张夏组中, 碳酸盐岩风暴沉积及遗迹化石也比较发育, 基于风暴岩的沉积序列分析, 识别出5种风暴沉积类型和6种遗迹化石(Skolithos, Palaeophycus, Planolites, Thalassinoides, Arenicolites和Taenidium), 并自下而上将风暴沉积序列划分出3个层段; 通过对风暴岩沉积序列、遗迹化石及岩性变化等特征分析, 表明洛阳龙门地区寒武系第三统张夏组的沉积环境从水体较浅的潮坪环境逐渐加深为半闭塞、较安静的局限台地环境, 持续海进形成开阔台地正常浅海环境, 然后水体深度逐渐下降, 变为开阔台地高能浅滩环境(郑伟等, 2015)。
山东省中部淄博、峨庄、莱芜一带的寒武系崮山组中保存有完整的风暴沉积序列, 由底至顶划分为6个层段: (1)底部风暴侵蚀层, 滞流沉积物中有砾石和三叶虫化石, 底面凹凸不平, 常保存有Thalassinoides, Palaeophycus, Planolites和Phycodes等遗迹化石; (2)风暴沉积层, 具平行层理, 缺乏生物成因的沉积构造; (3)风暴消减沉积层, 岩性为粒泥状灰岩或泥状灰岩, 含遗迹化石Teichichnus, Palaeophycus和Planolites, 有时具负载构造和粒序层; (4)风暴后侵蚀层, 侵蚀面之下产有丰富的Skolithos, Diplocraterion等, 并具生物钻孔构造; (5)风暴后侵蚀层和正常天气下沉积层之间的过渡层, 为密集堆积的三叶虫碎屑层, 厚1~10 mm, 三叶虫化石有Blackwelderia, Neodrepanura等, 有时含泥砾、具粒序层理; (6)正常天气下沉积层, 主要为薄层状泥岩、页岩和泥灰岩(周志澄等, 2013)。
四川龙门山地区中泥盆统养马坝组上部发育一套砂质近源风暴沉积。这套风暴沉积中遗迹化石丰富, 已识别出5个遗迹属, 包括Chondrites, Changchengia, Rhizocorallium, Skolithos和Zoophycos等。依据遗迹化石的分布位置、相互间的切割关系, 建立了龙门山地区养马坝组风暴岩中各种遗迹化石形成的先后顺序, 依次为Chondrites, Zoophycos, Rhizocorallium, Changchengia和Skolithos。根据遗迹化石的组成与分布特征, 将其划分为Chondrites-Zoophycos遗迹化石组合类型, 产生于正常浪基面以下、风暴浪基面以上的浅海沉积环境(荆锡贵等, 2019)。尤其是对白流坪组和养马坝组中Zoophycos进行详细研究后, 识别出3种风暴产生或诱导的沉积序列: 类型A, 由具有介壳堆积和波状纹理的粉砂岩组成, 顶部发育小型 Zoophycos; 类型B, 以含长波长丘状交错层理的细砂岩和灰黑色泥岩以及顶部大而密集的Zoophycos为特征; 类型C, 为含有短波长丘状交错层理的生物碎屑灰岩、泥质粒泥状灰岩及顶部大型的Zoophycos遗迹化石; 从正常浪基面至风暴浪基面依次为类型A→ 类型B→ 类型C(Li et al., 2018)。通过对龙门山甘溪地区下泥盆统碎屑风暴沉积序列中遗迹化石的产状及分布特征分析, 已建立3种遗迹组构: ① Zoophycos-Chondrites遗迹组构, 常常出现在黑色薄层泥岩和灰色薄层砂岩中; ② Zoophycos-Chondrites-Thalassinoides遗迹组构, 发育在厚层细粒砂岩夹泥岩的地层中; ③Zoophycos-Thalassinoides-Palaeophycus遗迹组构, 主要产生在灰色厚层细粒砂岩的层面上。这3种遗迹组构的化石特征表明, 当时的沉积背景是在风暴过后出现了短暂的缺氧环境, 而且Zoophycos和Chondrites的造迹生物是机会种生物(r-选择), 适存于大的风暴后安静的多营养花园式生态环境(Zhang, 2014)。
豫西北及晋东南地区下二叠统太原组碳酸盐岩中也发育多层风暴沉积并含丰富的遗迹化石, 已识别出7种风暴沉积类型, 与之伴生的遗迹化石有Zoophycos, Palaeophycus, Nereites, Chondrites, Planolites, Palaeophycus, Helminthopsis, Teichichnus, Rhizocorallium, Thalassinoides和Ophiomorpha等。胡斌等(2014)研究表明, 不同类型风暴沉积中的遗迹化石组成与分布有明显差异。
四川广安谢家槽一带下三叠统夜郎组和嘉陵江组中已识别的遗迹化石有17种, 它们可归属于浅海沉积环境中常见的Cruziana遗迹相。风暴沉积主要发育3个沉积序列单元, 自下而上为风暴沉积层、风暴消减沉积层及风暴后侵蚀层。其中, 风暴沉积层以鲕粒层和粗砂岩为特征, 因造迹生物在严酷的生态条件下难以生存而缺少遗迹化石, 仅局部见Skolithos; 风暴消减沉积层沉积物粒度一般较细, 以泥状灰岩或泥粒状灰岩为主, 遗迹化石主要为Palaeophycus和Planolites; 风暴后侵蚀层以起伏不平的侵蚀面为特征, 遗迹化石Skolithos从侵蚀面垂直向下伸入到风暴消减沉积层中。机会种遗迹化石如Treptichnus富集在纹层状灰岩中, 指示一种贫氧的沉积环境(周志澄等, 2014)。
6.1.2 浊流沉积中的遗迹化石组成与分布特征
与风暴沉积中的遗迹学研究相比, 近几年在较深水浊流沉积中的遗迹学研究成果显得较少。海相浊流沉积中较详细的遗迹化石研究主要集中在新疆准噶尔盆地西部地区。准噶尔西北部萨吾尔山至塔尔巴哈台山一带塔尔巴哈台组(D3-C1t)上部主要发育典型的远源浊积岩。纵瑞文等(2014)在该浊积岩中识别出Cosmorhaphe, Neonereites, Phycosiphon, Chondrites, Scalarituba, Zoophycos和Planolites等遗迹化石。这些遗迹化石中含有Nereites遗迹相的典型分子, 绝大多数都是半深海— 深海浊积岩中的常见属种。西准噶尔地区发育浊流沉积的地层自下而上依次划分为上泥盆统— 下石炭统塔尔巴哈台组、下石炭统的包古图组、太勒古拉组和下— 上石炭统哈拉阿拉特组。范若颖等(2015)在该套复理石相地层中识别出的遗迹化石有23属, 这些遗迹化石构成具有典型深水Nereites遗迹相特征的Phycosiphon-Nereites遗迹化石组合和Megagrapton-Glockerichnus-Cochlichnus 遗迹化石组合。前一组合在塔尔巴哈台组、包古图组、太勒古拉组和哈拉阿拉特组均有广泛分布, 以Phycosiphon和Nereites的大量出现为其典型特征, 其他分子还有Chondrites, Zoophycos, Lophoctennium等, 主要由底栖生物的牧食、进食构造构成, 其中Chondrites和 Zoophycos为中— 深阶层遗迹化石, 而Phycosiphon和 Nereites为较浅阶层遗迹化石, 属Nereites遗迹相中的Nereites遗迹亚相, 代表海底扇中— 远源富泥质浊流环境; 后一组合以雕画迹(Megagrapton, Glockerichnus)为特征, 此外还出现有Gordia, Planolites, Chondrites 等遗迹化石, 可归于Nereites遗迹相的Paleodictyon遗迹亚相, 代表海底扇中— 远源富砂质浊流环境。这2个遗迹组合均产自海底扇中下扇扇叶— 扇缘— 盆地平原环境。
遗迹学理论在油气地质勘探中的应用主要包括2个方面: 一方面是通过对遗迹化石、遗迹组合和遗迹相的研究来判识沉积环境, 进而通过沉积相控理论确定烃源岩与储集层的分布规律; 另一方面是通过对生物扰动构造或遗迹组构的研究来精细表征油气储集层的物性特征。近十年来, 中国学者在遗迹组构(生物扰动)影响储集物性方面做了大量的工作, 在碎屑岩储集层和碳酸盐岩储集层方面的研究都取得了许多新成果。
6.2.1 碎屑岩储集层中的生物扰动作用
在碎屑岩储集层方面, 杨伟芳等(2010)通过对塔里木盆地石炭系东河砂岩段中生物扰动作用的研究发现, 该地区东河砂岩段的高能碎屑滨海相临滨亚相沉积中含大量的遗迹化石或生物扰动构造, 可辨认的遗迹化石包括 Thalassinoides, Ophiomorpha, Skolithos, Palaeophycus 和 Planolites 等, 组成了3个遗迹组构。从上临滨到中临滨再到下临滨微相的沉积中, 层理规模由大变小, 泥质表现形式由泥纹到泥质条带, 生物扰动强度由弱变强。多数宏观生物扰动作用破坏原生沉积构造的现象十分明显, 但有些隐性生物扰动作用破坏原生沉积构造不明显, 甚至使沉积物内部变得更加均一。Qi等(2012)在分析塔里木盆地中部地区东河砂岩中生物扰动与储集层物性的关系时发现, 随着生物扰动强度的增加, 岩屑砂岩具有孔隙度增加、渗透率增大的趋势, 生物扰动构造与储集层物性呈正相关; 而在石英砂岩中, 生物扰动与储集层物性呈负相关。林世国等(2012)通过对四川盆地上三叠统生物扰动与储集性能的研究, 划分出4种岩性— 生物扰动组合: (1)厚泥层扰动, 主要形成于厚层黑色泥岩中, 反映滨湖沼泽或浅湖沉积环境; (2)砂泥边界扰动, 发育于厚层深灰色泥岩夹薄层浅灰色粉— 细砂岩中, 扰动指数为2~3, 反映远端砂坝下部环境; (3)砂泥薄互层扰动, 主要发育在砂泥岩薄互层中, 砂层单层厚度为2~8 cm, 扰动指数为4~5, 反映三角洲前缘远端砂坝上部环境; (4)厚砂层扰动, 主要发育于厚层砂岩中, 扰动指数为1~2, 表明其形成于水体能量相对较强的水下分支河道环境。研究认为, 生物扰动构造对储集层物性的影响较大, 生物扰动强度与储层物性呈正相关, 即生物扰动越强, 储层物性越好; 而弱扰动或未扰动部位原生孔隙胶结致密, 孔渗性都非常差, 这主要是因为生物扰动可以提高岩石的均质性和岩石颗粒的分选性。
6.2.2 碳酸盐岩储集层中的生物扰动及遗迹组构
在碳酸盐岩储集层方面, 牛永斌等(2008)在研究柴达木盆地南缘石炭系碳酸盐岩中的遗迹化石时发现, 生物扰动不仅能够改善储集层的孔隙度, 而且可使上、下储集层贯穿, 增加储集层的连通性, 从而可为油气储集层的预测研究提供依据。显微镜下观察显示, 研究样品中Chondrites遗迹化石的潜穴多被方解石充填, 而方解石质的潜穴充填物多被溶蚀, 形成了有效的储集空间; 另外, 潜穴内的充填物与潜穴壁之间的接触关系比围岩内部更容易增加孔隙的连通性, 对储集层物性的改良也具有重要作用。张卿等(2013)通过对四川盆地北缘奥陶系宝塔组灰岩网纹构造特征及油气输导作用的研究, 认为网纹构造是受生物扰动及构造活动因素影响而在同生— 准同生阶段深水沉积环境下的产物, 四川盆地北缘奥陶系宝塔组与上覆志留系的龙马溪组为一个生— 储油系统, 奥陶系宝塔组纵横交错的网纹是志留系龙马溪组生成油气的运移通道, 且在成藏过程中起到关键作用。网纹中充填物的矿物成分表明, 在使用压裂技术下网纹易于改造成为有效的空间通道。因此, 在勘探开发过程中, 可将奥陶系宝塔组致密气和志留系龙马溪组页岩气整体作为一个非常规油气系统一并开发, 勘探效益会更高。董小波和牛永斌(2015a)在豫西北奥陶系马家沟组三段碳酸盐岩中识别出较多的生物扰动构造, 其中已鉴定的遗迹化石有Thalassinoides callianassa, Helminthopsis abeli, Chondrites cf. intricatus, Palaeophycus tubularis和Planolites montanus, 丰度较高(潜穴的体积可占据所采岩样的60%~90%), 且在地层的横向上和纵向上分布均较广泛。通过野外露头实地观测和室内薄片分析, 认为豫西北奥陶系马家沟组三段豹斑灰岩的主要成因为生物扰动潜穴的不均匀白云化作用。生物潜穴充填物经历了白云石化、胶结、溶蚀、压实/压溶、去白云化等成岩作用, 其中溶蚀作用对充填物孔隙发育具有建设性, 压实作用和去白云石化作用具有破坏性, 而白云石化作用、胶结作用和压溶作用则具有建设性和破坏性的双重影响。生物潜穴充填物的孔隙类型主要以白云石晶间孔、晶间次生溶孔和晶内次生溶孔为主。随着成岩环境的变化, 孔隙度经历了降低→ 升高→ 降低→ 升高的反复演化过程。由于不均匀的白云石化作用使岩石呈明显的花斑状、云雾状, 常构成豹斑灰岩(董小波和牛永斌, 2015a, 2015b; 刘梦瑶等, 2020)。这类白云石化作用一般发生在埋藏期。白云石化作用初期, 受扰动的斑块优先被白云石化, 如果白云石化进一步加强, 则表现为斑块内部的白云石晶体较周围未受扰动区域的要粗。在鄂尔多斯盆地马家沟组和三山子组、川西北中二叠统栖霞组等, 无论是野外露头还是钻井岩心, 这种由生物扰动造成的不均匀白云化现象十分普遍和强烈, 值得重视。在对不同生物扰动构成的遗迹组构进行研究时还发现, 不同遗迹组构类型也会出现不同程度的白云石化作用。在豫西北奥陶系马家沟组碳酸盐岩中已识别出3种类型的遗迹组构, 即: Balanoglossites遗迹组构、Planolites遗迹组构和Chondrites遗迹组构。这3类遗迹组构中生物潜穴的充填物普遍发生了不同程度的白云石化作用, Planolites遗迹组构最为强烈, Balanoglossites遗迹组构次之, Chondrites遗迹组构最弱。通过柱塞孔隙度和渗透率测试分析, 表明这3类遗迹组构在孔隙度和渗透率等岩石属性参数上与围岩基质也具有很大不同, 含Balanoglossites遗迹组构的柱塞孔隙度和渗透率值最高, 含Chondrites遗迹组构的柱塞孔隙度和渗透率值次之, 含Planolites遗迹组构的柱塞孔隙度和渗透率值最小。进一步分析还表明, 各类遗迹组构对岩石物性的改造效应主要受遗迹组构类型及形态、生物扰动强度、生物潜穴连通状况及其成岩特征等因素的控制(牛永斌等, 2019)。
6.2.3 生物扰动型储集层的表征与研究方法
关于生物扰动型储集层的表征与研究方法, 近几年也有一些新的探索。牛永斌等(2017)研究生物扰动对塔河油田奥陶系碳酸盐岩“ 基质储集体” 储集性能的改造效应时, 利用Adobe® Photoshop® software CS6软件提供的生物扰动数字图像分析软件包对岩心数字图像中生物扰动区域进行了识别和定量表征。塔河油田奥陶系中已识别出Thalassinoides型扰动潜穴和Helminthopsis型扰动潜穴, 其中前者最为发育, 扰动体积可达90%, 生物潜穴充填物的矿物成分以白云石为主, 可达90%以上, 宿主围岩为泥晶灰岩, 矿物成分为方解石。在综合提取生物扰动型储集层特征信息的基础上, 以沉积学和遗迹学理论为指导, 应用3ds Max软件和X射线显微镜对研究区奥陶系生物扰动型储集层进行了三维重构(牛永斌等, 2018a)。该研究虚拟再现了塔河油田生物扰动型储集层的内部结构与微观特征, 为后续研究这类储集层的宏观非均质性与微观各向异性、定量表征该类储集层微观尺度上的渗流特征奠定了基础; 此外, 还为完善塔河油田奥陶系油藏模型提供了一种新思路。
微生物成因沉积构造(microbially induced sedimentary structures, 缩写简称为MISS)是微生物群与沉积环境相互作用, 并通过微生物生命的新陈代谢、生长、破坏、腐烂等过程在沉积物中留下的各种生物— 沉积构造(Gerdes et al., 2000; Noffke et al., 2001)。在这个意义上, 微生物成因沉积构造也可被视为广义遗迹化石的一种(Seilacher, 2007)。有关微生物成因沉积构造及其生物地质过程的研究近年来发展迅速, 成为深刻认识地球早期生命演化以及生物与环境相互作用过程的重要桥梁。中国近十几年来有许多学者十分关注这一领域的研究, 主要在前寒武纪地层和P/Tr界线附近生物大灭绝与复苏之间的地层中开展了较多的工作。
中国前寒武纪地层中的MISS研究主要集中在华北地台中元古界碎屑岩沉积中, 以近岸浅水陆源碎屑沉积为主, 包括多向波痕、微生物席稳化波痕、微生物席碎片、微生物席平滑波痕、多种微生物席脱水形成的砂裂及不规则网状生长脊等。研究表明, 在缺乏障壁的粗碎屑环潮坪带, MISS主要集中在潮间带上部至潮上带下部, 以微生物席破坏构造为主, 生长构造和腐烂构造少见, 微皱痕构造也比较少见。在有障壁和潟湖发育背景下, MISS类型丰富, 鸟足状、纺锤状、曲线状等“ 不完全型” 微生物席破坏构造占有较大比例, 而撕裂和再沉积席片构造相对较少; 在相对低能的潟湖中, 各种微皱痕与气泡构造较多(史晓颖等, 2008a)。由于障壁沙坝阻隔, 潮上— 海岸带发育的“ 储水池” 通过地下水渗流为潮上带较高部位提供足够的湿度, 故MISS在潮上带较高部位也能大量发育。目前在地层记录中报道的大部分微生物席形成的沉积构造, 特别是微生物席的生长和破坏构造, 主要发育在这2种沉积环境中(汤冬杰等, 2011)。因此, MISS的形态组合和类型特征能够反映沉积微相变化。
中国中元古界中的MISS研究最早开始于华北地区。胡健民和孟庆任(1991)、杨式溥和周洪瑞(1995)、齐永安(2005)在豫西鲁山地区汝阳群云梦山组、白草坪组和北大尖组, 刘洪福和刘池洋(1992)在蓟县长城群串岭沟组, 华洪等(1993)在宁夏贺兰山长城系黄旗口组, 高建华等(1993)在天津蓟县长城系大红峪组都分别发现这类微生物成因沉积构造, 并都把它们作为遗迹化石来研究。发育在天津蓟县至河北兴隆一带中元古界长城群细粒碎屑岩中的纺锤状裂缝, 曾经被解释为后生动物遗迹化石或地震震荡液化脉等, 现在被认为是由于微生物席对沉积面的封闭作用导致沉积物在成岩作用早期发生脱气与脱水等作用的产物, 常与变余波痕、皱饰构造等相伴产出(李林等, 2008)。长城群的常州沟组、串岭沟组、大红峪组碎屑岩也发育变余波痕、皱饰构造和砂质碎片等微生物成因沉积构造(梅冥相等, 2007; 李林等, 2008)。高于庄组中部产出的臼齿状构造, 可能是一种形成于较深水环境中的与藻类生物活动相关的生物沉积构造(梅冥相, 2005)。在对天津蓟县、北京西山和昌平及河北兴隆、宽城等地串岭沟组的观察与研究中, 发现地层中普遍发育一种形态特殊的砂质脉状构造, 在层面上表现为不规则密集分布的细长砂脊或弯曲的砂质条带, 在垂直断面上多呈近直立或 “ 肠状” 弯曲的小型脉状体, 形态与中— 新元古界碳酸盐岩中广泛发育的臼齿构造颇为相似。这种密集发育的砂脉构造被认为是目前地层中识别出的最古老的甲烷排放证据, 是微生物成因的, 而不是后生动物形成的遗迹化石(史晓颖等, 2008b)。梅冥相等(2009)则把在天津蓟县剖面串岭沟组下部环潮坪相粉砂质泥页岩细粒沉积中的这种构造定义为粉砂岩岩墙, 认为是由微生物形成的沉积构造。史晓颖等(2008b)、Lan等(2013)总结了贺兰山中元古界黄旗口组石英砂岩中发育的微生物成因沉积构造, 包括由微生物席生长、破坏和腐烂过程形成的3种类型、9种不同形态的构造, 表明1.6 Ga前以蓝细菌为主的微生物群在环潮坪碎屑环境中也很活跃, 可能代表了微生物由海洋向陆地环境发展的过渡阶段。
邢智峰等(2011, 2020)研究了豫西中元古界云梦山组中发育的大量微生物成因沉积构造, 在地层纵剖面上毫米级的深色沉积物层和浅色的石英颗粒层交替出现, 形成典型的微生物席纹层, 代表着微生物席在沉积表面的多次生长和埋藏, 其中深色层包含有泥质物和细砂— 粉砂颗粒, 被认为是先前微生物席的残留区, 而浅色层则是较纯净的石英颗粒, 由物理沉积作用形成。对纹层区的薄片进行观察, 揭示出5种明显的MISS微结构类型, 分别是波曲层、网状结构、定向颗粒层、细小颗粒层和重矿物层。这些微结构代表了微生物席在沉积物表面殖居、生长、代谢以及与沉积物相互作用的一些特征, 并指示了当时的水动力状况和古环境特征。同时, MISS的大量发育表明了当时微生物群落的繁盛, 揭示出微生物群落对古环境具有的强大改造功能。
梅冥相(2011)论述了陆源碎屑岩中微生物诱发的沉积构造的成因类型及其分类体系, 详细总结了保存在砂岩和页岩中的生物膜或生物席的各种特征, 分析了以往长期有成因争议的碎屑岩中出现的满洲藻迹(Manchuriophycus)、肯尼亚构造(Kinneyia)和星状迹(Astropolithon)这3种不同形态且与微生物作用有关的沉积构造。可以认为, “ 满洲藻迹” 代表了一种特殊的微生物席收缩裂缝(如见于河南卢氏县城北的元古界高山河组的标本), 而且属于“ 微生物席的物理破坏作用类型” 的微生物诱发的沉积构造或微生物稳定作用的产物; 肯尼亚构造(如见于河南汝阳县中元古界云梦山组的标本)可归因于微生物席的埋藏和腐烂过程产生的气体捕获过程的产物, 类似于罕见的“ 砂火山” 式的 “ 星状迹” 构造, 从中心管向外放射的岩墙可能只是形成在一种具有黏性的宿主沉积物之中, 在这种情况下最有可能的是微生物粘结的砂, 所以归到砂岩中的微生物席特征。陈留勤(2013)发现, 河北兴隆一带的长城系大红峪组(1650-1600 Ma)以潮下坪灰白色厚层块状细粒石英砂岩为主, 在砂岩层面上产出丰富的微生物成因构造, 可能反映了华北中元古代浅海和潮坪环境被大面积分布的微生物席所覆盖, 微生物席对砂质沉积物表面起到有效的保护作用, 在一定程度上降低了沉积物的侵蚀和改造作用, 从而影响基本沉积作用过程。郑伟等(2016b)等研究了豫西鲁山地区中— 新元古界汝阳群和洛峪群微生物席及其矿物显微构造特征, 识别出3大类9小类微生物成因构造, 根据MISS石英砂岩的镜下石英颗粒及其他矿物组分方面特征可以得出: (1)宏观形态各异的MISS镜下矿物特征无明显差别; (2)MISS形成过程存在着 2种生物沉积作用, 即生物化学沉积作用和生物物理沉积作用; (3)微生物群落多生长在中等水动力、沉积物为细粒的沉积环境中。此外, 其他学者也相继报道了山西黎城、天津蓟县和辽宁兴城等地中元古界微生物成因构造, 主要是对构造类型组合、微观蓝细菌类型以及古环境意义等进行了研究(郑伟和邢智峰, 2015; Yang et al., 2017; 李晓波等, 2020)。
豫西宜阳地区二叠系— 三叠系界线附近陆相碎屑岩中发育以微生物席生长特征、微生物席破坏特征为主的微生物成因相关沉积构造。该地区上二叠统孙家沟组属湿润— 干旱气候条件下的滨、浅湖碎屑岩沉积, 上覆地层下三叠统刘家沟组属干燥气候条件下的湖泊三角洲相及河流相。微生物成因沉积构造主要产出于孙家沟组上部至刘家沟组的中下部。其中, 砂岩层面上与微生物席生长过程相关的特征与构造主要有变余波痕、斑状波痕、微生物席平滑波痕、皱饰构造、生长脊、瘤状、刺状突起等, 而与微生物席破坏过程相关的特征与构造主要有砂质裂缝、微生物席碎片、卷曲边缘和侵蚀凹坑等。张利伟等(2014)研究认为, 在这些以微生物席生长特征和微生物席破坏特征为主的微生物成因沉积构造发育的地层中, 未发现其他实体化石和遗迹化石, 显然这与二叠纪末生物大灭绝事件或灾变事件造成的特殊水体化学环境和严重退化的生态系统相关。邢智峰等(2018)将宜阳地区刘家沟组微生物成因构造按照形态特征的演化自下往上分为4个反映不同古环境与古气候的组合: 皱饰构造组合、隆脊构造组合、小型脱水裂痕组合和大型脱水裂痕组合, 认为该地区微生物成因沉积构造的发育与演化是对早三叠世陆相古环境细微变化的联动响应。Chu等(2015)研究了河南多地下三叠统刘家沟组滨浅湖碎屑岩中出现的皱饰构造(Wrinkle structures)及其伴生的有稀少的具回填纹构造的遗迹化石, 认为这种陆相皱饰构造反映了一种异常的水化学和物理环境, 推测与陆生生物灭绝有关。虽然在晚二叠世末(P-Tr)生物大灭绝后出现的遗迹化石很稀少, 但它们大多与微生物席一起保存, 这表明微生物席可作为水生底栖动物的食物和氧气来源。在豫西荥阳地区与宜阳地区相当的孙家沟组和刘家沟组中, 也发现了丰富的微生物成因沉积构造(郑伟等, 2016a), 可分为3大类(即席生长构造、席破坏构造和席腐烂构造)和11个小类, 即瘤状突起、变余波痕、隆脊构造、似正弦状脱水裂痕、纺锤状脱水裂痕、树枝状脱水裂痕、多边形脱水裂痕、曲形脱水裂痕、砂火山构造、微生物席砂片和似雨痕构造等。微观分析显示, MISS脊处纵切面显示U型、石英颗粒和黏土矿物条带定向排列、捕捉的云母颗粒不连续和近平行等特征, 均表明其为微生物成因。因此, 这些丰富的微生物成因构造是二叠纪末灾变事件导致陆地生态系统迅速退化、微生物大量繁殖而形成的, 代表着以微生物群落为基础、后生生物少、钙质结核发育等特殊沉积的灾后陆地生态系统, 其蕴藏的微生物演化信息以及与后生生物演化关系的沉积记录, 为海、陆相微生物席的空间分布提供了新的资料, 对全面了解二叠纪末豫西陆相生态系统具有重要的意义(郑伟等, 2016a)。Chu等(2017)详细研究了华北(河南、山西)地区下三叠统刘家沟组碎屑岩沉积中微生物成因构造, 包括“ 老象皮” 构造、皱饰构造、变余波痕、“ 正弦状” 构造、蠕虫状构造、气隆构造和砂裂构造等, 认为产生这些MISS的陆生微生物群落不仅要适应干燥的气候条件, 而且还要适应风暴为主的古环境。在P-Tr生物大灭绝后, MISS变得更常见和广泛, 表明生物大灭绝为陆生生态系统中MISS的发育提供了生物和环境条件。
文中从寒武纪遗迹组合、组构与底质革命, 奥陶纪— 泥盆纪遗迹组合、组构、时空演化及生态习性, 石炭纪— 二叠纪遗迹组合、组构与古氧相, 三叠纪海相地层及P/Tr界线附近遗迹化石组成与分布特征, 陆相遗迹化石组合与组构, 微生物成因沉积构造及遗迹学在事件沉积和油气地质勘探中的应用等多个方面, 总结了近十年来中国遗迹学研究的主要内容和取得的进展。但与国际上发达的国家相比, 中国学者在遗迹分类学、遗迹组构阶层分析、生物扰动与储集层物性的关系、高新技术在精细识别生物遗迹上的应用和三维遗迹形态的表征等方面还存在一定差距, 这也是今后应进一步努力工作的方向, 同时还要重视现代生物遗迹学的研究及其与遗迹化石的比较分析。
因近十年来中国遗迹学领域发展较快, 涉及沉积学和能源地质勘探方面的学术论文较多, 发表的渠道较广, 信息量较大, 难以全面完整地收集, 所以如有疏漏之处, 敬请读者谅解。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
| [147] |
|
| [148] |
|
| [149] |
|
| [150] |
|
| [151] |
|
| [152] |
|
| [153] |
|
| [154] |
|
| [155] |
|
| [156] |
|
| [157] |
|
| [158] |
|
| [159] |
|
| [160] |
|