

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
微生物碳酸盐岩的显微结构基本特征*
[吴亚生1, 2, 3  , 姜红霞
, 姜红霞4 , 李莹1, 2, 3 , 虞功亮5 ]
, 姜红霞, 李莹|
|

通讯作者简介 姜红霞,女,1976年生,博士,河北地质大学助理研究员,主要从事微生物岩、珊瑚和层孔虫古生态学研究。E-mail: 2928984509@qq.com。
第一作者简介 吴亚生,男,1963年生,博士,中国科学院地质与地球物理研究所副研究员,中国科学院大学岗位教授,主要从事碳酸盐岩古生物学、生物碳酸盐岩与微生物沉积学研究。E-mail: wys@mail.igcas.ac.cn。
以前的微生物岩识别和分类只根据裸眼可见的中观结构,而对显微镜下的微观结构未予使用。微生物的显微结构有什么基本特征,是否可以用于微生物岩的识别和分类是本文关注的焦点。作者从微生物岩形成机制和实际材料的观察来探讨这个问题。在诱导钙化实验中观察到,蓝藻鞘丝藻 Lyngbya的胶鞘表面有碳酸盐矿物形成: 先是在胶鞘表面形成点状碳酸盐颗粒,后来碳酸盐几乎包裹整个丝体,形成一个矿物壳。微生物岩形成的模式推演表明,微生物岩一般都有微生物留下的孔(模孔)和包围模孔的矿物壳2种基本单元。对3个地点的现代微生物碳酸盐岩和1个地点的古代(寒武纪)微生物岩的观察,发现在显微尺度上都具有模孔,此外还可能有矿物壳或胶结物。模孔的形状、大小、排列方式是微生物碳酸盐岩显微结构研究的核心内容,在今后的研究中可以用于微生物岩的识别和分类,以及作为分析形成机制和形成环境的基础。
About the corresponding author Jiang Hong-Xia,born in 1976,is an assistant professor at Hebei GEO University. Her studies concentrate on microbialites,paleontology of stromatoporoids and corals.
About the first author Wu Ya-Sheng,born in 1963,is an associate Professor at Chinese Academy of Sciences and a professor at University of Chinese Academy of Sciences. His studies focus on biosedimentology,microbial carbonate,and paleontology of carbonates. E-mail: wys@mail.igcas.ac.cn.
Previously microbialites were identified and classified based only on mesofabrics, i.e.,the fabrics visible to naked eyes,but not on the basis of microfabrics, i.e., the fabrics visible under the microscope. This paper focuses on the basic characteristics of microfabrics of microbial carbonates and microbialites,and whether they can be used for their identification and classification. An experiment on microbe-induced calcification that we conducted shows that among a variety of microbes,only the filaments of the cyanobacterium Lyngbya form carbonate minerals on the surface of their sheaths,firstly as scattered carbonate particles,and finally as crusts almost enveloping the whole filaments. It is thus proposed that the microbial carbonates that are formed by induced precipitation have two basic constituents,namely,the mold holes, i.e.,the holes left by the microbes,and the carbonate crusts. Microscopic observations of the modern microbial carbonates from the Shark Bay of Australia,Highborne Cay of Bahamas,and Atoll of Kiritimati and the Cambrian dolomitized microbialites in Xinjiang,China revealed that mold holes and the crusts or cements between the pores are a basic characteristic. The shape,size and arrangement of the mold holes and proclots are the core contents of the study on the microfabrics,and these features can be used for the identification and classification of microbialites,as well as the basis for the analysis of their formation mechanism and environments.
微生物岩的原始定义(Burne and Moore, 1987)指底栖微生物群落与碎屑或化学沉积物相互作用形成的沉积, 是通过微生物群落(1)对碎屑沉积的捕集和粘结作用、(2)无机钙化作用、(3)生物影响的钙化作用形成的沉积; 这3种作用形成的岩石分别叫微生物粘结岩、微生物泉华、微生物骨架岩; 生物影响钙化作用可能是光合作用引起的化学变化或者是晶体在微生物胶鞘表面成核的结果(Burne and Moore, 1987); 微生物岩的内部结构包括叠层石型的、凝块石型的、核形石型的、球形的、隐形的(Burne and Moore, 1987); 叠层石主要是微生物捕集和绑结作用形成的, 凝块石主要是生物影响的钙化作用形成的。叠层石是生物成因的具有细纹层构造的碳酸盐岩块体(Kalkowsky, 1908; Awramik and Margulis, 1974; Krumbein, 1979; Burne and Moore, 1987)。凝块石的原始定义(Aitken, 1967)指未钙化的蓝藻和绿藻形成的、具有宏观的凝块结构或海绵结构的岩石; 凝块由微晶(8~20 μ m)灰岩组成。虽然后来的学者提出了一些新的微生物岩类型, 如树枝石和均一石(Riding, 1991, 2011a), 但是迄今未见可靠的实例报道。
由于微生物岩的定义来自于宏观特征, 所以微生物岩的识别都是根据宏观特征进行的, 即根据对露头、手标本、光面、薄片的肉眼直接观察确定(Aitken, 1967, 图 19; Burne and Moore, 1987, 图 10; kennard and James, 1986, 图 2; Kah, 1992, 图 2, 图 3; Grotzinger, 2000, 图 6; Shapiro, 2000, 图 3, 图 4; Riding, 2011b, 图 8, 图 10)。一些学者的文章中虽然有微观照片, 但是微观特征并没有用于微生物岩的识别和分类。中观特征固然是微生物岩的重要特征, 但某些中观特征是成岩作用形成的, 并不是微生物的作用形成的, 例如二叠纪— 三叠纪界线地层的微生物岩在露头上表现为凝块状, 但是, 深入研究后发现是成岩作用形成的结构(Wu et al., 2014; 吴亚生等, 2018)。根据中观特征识别微生物岩的另一个缺陷是不能对小的标本、特别是小于1 cm的标本进行识别。而在石油工业中大量的钻井是不取心的, 岩性的确定是根据岩屑; 岩屑一般小于1 cm。微生物岩能不能根据微观特征来识别, 微生物岩的显微结构有什么基本特征、是否可以用于微生物岩的识别和分类, 是作者关注的主要问题。作者从微生物碳酸盐的形成机制上讨论其显微结构特征, 并且用现代和古代微生物岩的实例来验证理论推测。
微生物作用主导形成的矿物叫微生物矿物; 微生物作用主导形成的岩石叫微生物岩; 微生物作用主导形成的碳酸盐矿物组成的泥晶或颗粒形成的沉积物叫微生物碳酸盐沉积; 微生物作用主导形成的碳酸盐矿物的建造或岩石, 叫微生物碳酸盐岩或简称微生物岩。建造的含义是岩石的骨架是由原位的微生物形成的。微生物作用形成的碳酸盐矿物、微生物作用形成的碳酸盐沉积物、微生物作用形成的碳酸盐建造或岩石, 统称为微生物碳酸盐岩。
微生物矿化指微生物体的某个部分、某些部分或全部变成矿物的现象。微生物诱导结晶指微生物作用引起的矿物结晶。国外文献中将微生物诱导结晶作用笼统叫做微生物诱导矿化。微生物诱导结晶作用形成的矿物覆盖微生物体的表面形成的壳状结构叫矿物壳。
作者通过一个微生物碳酸盐的培养实验来增加对微生物碳酸盐形成过程和机制的认识, 然后根据对形成机制的认识, 推断微生物碳酸盐岩应具有的显微结构特征。再通过对现代和古代微生物岩实例的观察, 确定它们是否有预测的显微结构特征。
微生物碳酸盐培养实验于2018年7月至10月间在中国科学院地质与地球物理研究所进行。实验容器为一个30× 50× 30 cm大小的无盖塑料箱, 装10 cm厚自然土壤, 加入自来水, 使水深达到10 cm, 温度22~35 ℃, 正常日光照射。
根据文献调研, 巴哈马、鲨鱼湾、基里巴斯都有典型的现代微生物碳酸盐岩在形成。由于不方便采样, 作者根据前人文献中微生物碳酸盐岩的显微照片分析其显微结构特征。
为了观察古代微生物碳酸盐岩的显微结构特征, 此次研究对新疆塔里木盆地阿克苏城西南苏盖特布拉克村附近寒武系第三阶剖面的微生物岩进行剖面测量, 并采集标本, 制作薄片, 在偏光显微镜下观察其显微结构特征。
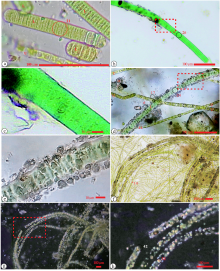
实验2018年7月份开始。1周后, 土壤表面生长出深蓝色的丝状微生物, 镜下确定微生物包括蓝藻的鞘丝藻属Lyngbya和丝状绿藻丝藻属(Ulothrix); 14 d后, 显微镜下见到少数Lyngbya的胶鞘表面出现一些颗粒状的碳酸盐(图 1-b, 1-c, 1-d, 1-e, 1-g, 1-h; 图 2-a, 2-b, 2-e, 2-f); 23 d后, 很多Lyngbya丝体表面有碳酸盐颗粒, 颗粒呈点状, 直径1.8~6.4 μ m; 35 d后, 取样做扫描电镜观察, 见到一部分Lyngbya丝体胶鞘表面全部被碳酸盐颗粒覆盖(图 1-f右下角; 图 2-c, 2-d), 形成一个包裹丝体的矿物壳。但是, 丝体的前端表面没有碳酸盐矿物; 很短的Lyngbya丝体、新形成的丝体表面没有碳酸盐矿物形成。始终未见其他蓝藻属种或其他藻类表面有碳酸盐矿物形成。10月份气温下降到10 ℃以下, 箱中的微生物死亡。
 | 图 1 蓝藻诱导结晶培养实验的照片 a— 4个粗的是蓝藻丝体(L: Lyngbya), 表面尚无碳酸盐颗粒形成; 1个细的是绿藻丝体(U: Ulothrix); b— 单偏光照片, 1个蓝藻丝体(Lyngbya), 一半有碳酸盐颗粒; c— 是B的红色方框的放大; d— 1个蓝藻丝体(L: Lyngbya)和4个绿藻丝体(U: Ulothrix), 蓝藻丝体有胶鞘, 胶鞘表面覆盖有1层碳酸盐颗粒; e— 是d的红色方框的放大; f— 7个有矿物壳的蓝藻丝体(Lyngbya), 2个未钙化的蓝藻丝体(Lyngbya), 许多黄绿色的绿藻丝体(Ulothrix), 右下角的1个蓝藻丝体全部被碳酸盐覆盖, 其余的未被全部覆盖; g— 正交偏光, 有至少10个表面有碳酸盐颗粒的蓝藻丝体; h— 是g的红色方框的放大, 干涉色为高级白, 是方解石或文石的特征Fig.1 Photomicrographs of microbes in an experiment on induced carbonate precipitation by cyanobacteria |
 | 图 2 蓝藻丝体的扫描电镜照片及能谱图(与图 1同源的样品) a— 4个丝体表面有稀疏分布的颗粒状碳酸盐; b— 是a的红色方框的放大, 标出能谱测点位置; c— 3个蓝藻丝体表面全部被碳酸盐颗粒覆盖; d— 是c的红色方框的放大; e— 1个蓝藻丝体表面的碳酸盐颗粒; f— 是e的红色方框的放大, 显示碳酸盐颗粒由0.1× 1 μ m的棒状晶体组成; g和h— 是b中位置14和15处的能谱图Fig.2 Scanning electron microscope(SEM)graphs and energy spectrogram(EDS)(the same cyanobacterial samples as in Fig.1) |
该实验没有人工添加微生物菌种, 所以, 微生物全部来源于土壤中的微生物菌种。只有Lyngbya的表面有碳酸盐矿物形成, 说明一定的条件下只有某些微生物能够引起碳酸盐矿物形成, 即微生物碳酸盐的形成与微生物的种类有关: 有的属种能够形成, 有的属种不能形成, 说明诱导结晶作用确实存在。不是所有微生物的表面都有碳酸盐矿物形成, 说明矿物是诱导结晶作用形成的, 不是无机作用形成的; 如果所有微生物的表面都有碳酸盐矿物形成, 那么, 反而说明可能是无机结晶作用形成的。
在本实验条件下, 蓝藻Lyngbya丝体只在胶鞘表面形成碳酸盐, 而不在胶鞘内部形成。Riding(1977)观察到碳酸盐矿物在蓝藻Plectonema的胶鞘内部形成, 使之可以保存为化石。诱导结晶是在胶鞘表面形成碳酸盐矿物, 形成的矿物壳的厚度不均匀, 不能形成可以进行属种鉴定的化石。
诱导结晶是一个逐渐进行的过程, 最初只在微生物体表面形成少数点状分布的碳酸盐矿物颗粒, 然后数量逐渐增多, 再后覆盖整个微生物丝体的表面, 形成一个包围丝体的矿物壳。
微生物的生长是微生物碳酸盐形成的先决条件。微生物的生长需要一定的温度。扬州位于北纬32.39° , 北京市区位于北纬39.9° 。吴亚生曾对扬州和北京的河流及湖泊的水华发生情况进行了多年的观察, 发现无论是扬州还是北京, 冬季的河水表面是清澈的, 没有微藻水华, 说明蓝藻等微生物较少。但是, 在春夏秋季, 凡是富营养化的水体, 表层水都会因为水华的存在而呈绿色。微藻水华主要由蓝藻以及部分微小的绿藻形成。这个现象说明, 控制微生物生长的第一因素是温度, 第二因素是水体的营养元素含量。当然还有其他次要因素。
微生物碳酸盐必须是微生物作用形成的。Burne和Moore(1987)认为, 形成微生物碳酸盐的作用主要有2种: 捕集作用和生物影响的钙化作用。前者指丝状以及非丝状微生物的黏性胶鞘粘结和包裹沉积物的作用(注: 本文作者认为, 微生物的胶鞘未必是黏性的), 后者指微生物的作用使碳酸盐在生物体表面或者在胶鞘内部形成的作用。另外一些学者将生物有关的碳酸盐形成作用分为3种: 生物诱导、生物影响(Dupraz et al., 2009)和生物控制(McConnaughey, 1989; Lowenstam and Weiner, 1989; Grotzinger and Knoll, 1999; Reid, et al., 2000; Franke and Bazylinski, 2003; Reid et al., 2003; Weiner and Dove, 2003; Dupraz et al., 2004; Riding, 2011a; Kazmierczak et al., 2015)。生物诱导指生物的生理活动影响了环境, 例如, 微生物的光合作用导致微环境的pH上升, 从而导致碳酸盐矿物形成的作用。生物影响指生物的特性影响晶体的形状和成分的碳酸盐形成作用。根据作者对微生物诱导结晶实验的观察结果, 对前人提出的微生物碳酸盐形成模式做了细化和补充, 认为形成微生物碳酸盐的作用主要有3种(图 3): 捕集作用, 微生物诱导结晶作用, 生物控制钙化作用。
捕集作用指丝状或非丝状微生物的聚集体, 对沉积物的阻挡作用和保护作用。阻挡作用使水流中携带的泥晶或者颗粒沉积下来, 保护作用的含义是使已经沉积在微生物体之间的沉积物不被水流冲刷走。分2种情况:
1)微生物体之间的沉积物未能在微生物体腐烂降解之前半固结或固结, 这样, 在微生物体腐烂之后, 很可能不能在沉积物中留下孔洞(图 3-a)。如此形成的沉积物, 与水流中携带的沉积物由于重力作用而沉积形成的沉积物没有显著的区别, 因而难以识别。这种成因的岩石是否属于微生物碳酸盐有待商榷。
 | 图 3 以丝状蓝藻为例的微生物碳酸盐形成模式 a1~a3、b1~b3— 捕集作用trapping; c1~c3、d1~d3、e1~e3— 生物诱导结晶; f1~f3— 生物控制钙化。c1~c3— 微生物碳酸盐呈点状附着在微生物体表面; d1~d3— 微生物碳酸盐在微生物体表面形成壳, 壳之间的空间未被充填满; e1~e3— 微生物碳酸盐在微生物体表面形成壳, 且因为有充分的时间生长, 壳之间的空间被微生物碳酸盐充填满; f1~f3— 微生物碳酸盐只分布在胶鞘内部, 形成厚度均匀的壳, 即化石。a3如果能够识别, 也属于微生物碳酸盐, 但是难以识别; b3、d3、e3属于狭义的微生物碳酸盐; f3既属于微生物碳酸盐, 又属于微生物礁的范畴Fig.3 Formation models of microbial carbonates by filamentous cyanobacteria |
2)由于胶结作用, 或者由于露出水面而发生的脱水作用, 沉积物在微生物体腐烂消失之前固结或者半固结, 因此, 在微生物体腐烂降解之后, 微生物体在沉积物中留下孔洞, 称为模孔。模孔使沉积物的结构不均匀(图 3-b): 如果沉积物由于模孔定向排列或其他原因形成纹层结构, 就叫叠层石; 如果没有纹层结构, 就叫凝块石。
指微生物的生理活动, 例如光合作用, 导致其周围水体微环境的物理化学性质发生变化, 如pH值上升, 从而引起碳酸盐矿物结晶的作用(在英文中叫induced calcification, 但这是不恰当的叫法, 因为实际上不是微生物本身变成钙质)。实验表明, 这种作用形成的矿物一般是附着在微生物体的表面(如果微生物体有胶鞘, 那么指胶鞘的表面), 并且开始是呈孤立的点状, 然后逐渐覆盖微生物体的整个表面。可以分为4种情况:
1)碳酸盐呈点状分布在微生物体的表面(图3-c), 如果微生物体死亡分解, 那么, 碳酸盐矿物就以分散的颗粒存在, 形成松散的沉积物, 这种点状碳酸盐颗粒的大小在1.8~6.4 μ m, 亦可更大或更小。这种成因的岩石是否属于微生物碳酸盐岩亦有待商榷。
2)如果碳酸盐矿物形成包围微生物体的壳, 但是, 相邻微生物体的碳酸盐壳不相互联接, 那么, 微生物体死亡腐烂之后, 碳酸盐壳将形成松散的沉积物, 壳的内部有微生物体留下的孔, 叫做模孔。这种成因的岩石是否属于微生物碳酸盐岩同样有待商榷。
3)如果碳酸盐矿物形成包围微生物体的壳, 且相邻微生物体的碳酸盐壳之间相互联接, 碳酸盐矿物没有完全充填满壳之间的空间, 那么碳酸盐壳将形成多孔的微生物碳酸盐建造(图3-d)。这种建造是典型的微生物岩。在碳酸盐沉积学中, 建造指处于原位的生物骨骼构成的碳酸盐体。在微生物碳酸盐建造中, 壳处于生长时的位置; 壳的内部有微生物体留下的模孔。壳之间有未被充填的孔隙, 称为剩余孔洞。
4)如果碳酸盐矿物形成包围微生物体的壳, 而且壳之间的空间被碳酸盐矿物充填满, 那么, 将形成比较致密的微生物碳酸盐建造(图 3-e)。
生物控制钙化作用指生物的生理活动引起矿物在生物体内部或者表面的特定部位聚集, 形成具有特定形状、结构的骨骼的作用。例如, 蓝藻的生理活动促成碳酸盐矿物在蓝藻胶鞘内形成, 最终形成厚度均匀的壁。这种厚度均匀的壁保持了蓝藻的形状和大小(图 3-f); 它们如果被保存在地层中, 就叫化石。现代自然环境中, 如印度洋的亚达伯拉环礁(Aldabra Atoll)上的一个小水坑中就有生物控制钙化的织线藻Plectonema(Riding, 1977)。再例如, 双壳类外套膜的表面有碳酸盐晶体的形成, 形成有特定形状、结构和成分的壳, 构成外骨骼。珊瑚的骨骼具有同样的性质。
以上模式图是以丝状微生物为例的。但无论是丝状微生物, 还是非丝状微生物都可以发生捕集作用和诱导结晶作用, 例如, 太平洋的基里巴斯岛上的现代潟湖中正在形成的微生物碳酸盐建造主要由非丝状的微生物形成(Arp et al., 2012)。这说明, 典型的微生物碳酸盐建造或微生物岩都有微生物留下的模孔, 并且往往有包围模孔的矿物壳。
微生物碳酸盐在微生物席死亡之后就进入成岩阶段。目前所知最老的微生物碳酸盐岩是澳大利亚皮巴拉37亿年前的叠层石(Nutman et al., 2016)。古老地层中的微生物碳酸盐岩可能遭受了白云石化、溶蚀作用、胶结作用、去白云石化作用等各种成岩作用的影响, 尤其是白云石化作用、溶蚀作用和胶结作用。这些成岩作用会不同程度地改造或破坏微生物碳酸盐岩的原始结构, 但现代自然环境的微生物碳酸盐岩基本未受到成岩作用改造。
世界很多地方的现代自然环境中都有微生物碳酸盐岩在形成(Reid et al., 2003; Jahnert and Collins, 2011; Suosaari et al., 2016)。要确定这些微生物碳酸盐岩的显微结构是否都有模孔和矿物壳, 一种办法是去这些地方采样进行研究, 另一种办法是检查前人文章里的显微照片。作者采用第2种办法, 检索并研究了所有涉及现代微生物碳酸盐岩的文献, 发现只要有显微照片, 并且显微照片的分辨率足够高, 就能看到这些微生物碳酸盐岩有模孔和矿物壳。这里仅举鲨鱼湾、巴哈马、基里巴斯3个例子作为代表。
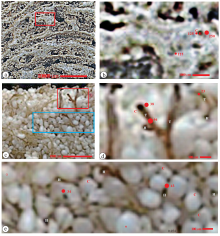
澳大利亚鲨鱼湾的叠层石举世闻名, 凝块石也同样重要。这2种微生物碳酸盐岩分布在鲨鱼湾的哈梅林潟湖。前人的研究发现, 凝块石分布在潮上带、潮间带上部和潮下带下部, 而叠层石只分布在潮间带下部和潮下带上部(Reid et al., 2003; Jahnert and Collins, 2011; Suosaari et al., 2016)。叠层石由钙质纹层与排列成纹层的孔洞交互组成(图 4-a; Jahnert and Collins, 2011, 图 3B)。放大后(图 4-b), 可见钙质纹层由模孔和其间的钙质组成。围绕模孔的钙质颜色浅, 应当为矿物壳, 厚度为125 μ m。模孔圆形或蠕虫状, 直径有125 μ m和250 μ m 2种。
 | 图 4 澳大利亚鲨鱼湾和巴哈马海布礁现代微生物碳酸盐岩的显微结构特征 a— 修改自Jahnert 和 Collins(2011)的图 3B, 被称为胶状叠层石Colloform structure。纹层状, 由钙质纹层和其间的孔洞组成的纹层组成; b— 是a的红色方框的放大, 见到钙质纹层由模孔(H)和其间的钙质组成; 模孔圆形或蠕虫状, 直径125、250 μ m; 模孔周围有白色的矿物壳(C), 其厚度125 μ m; c— 修改自Patterson(2014)的图 49A, 手标本, 鲕粒凝块石, 见到白色的鲕粒和其间的黄色和褐色藻丝体; d— 是c的红色方框的放大, 见到2种直径的藻丝体(T), 直径分别为23、46 μ m。粗藻丝体外面的乳白色矿物壳(C)厚度为38 μ m; e— 是c的蓝色长方形框的放大, 可见白色鲕粒(o)之间有很多模孔(H)和乳白色的矿物壳(C), 模孔的直径为34~43 μ mFig.4 Microfabrics of the modern microbial carbonates in Shark Bay of Australia and Highborne Cay, Bahama |
巴哈马是现代微生物碳酸盐岩的著名产地之一, 因被认为是现代正常浅海环境微生物碳酸盐岩的唯一代表而地位特殊。巴哈马的微生物碳酸盐岩包括叠层石和凝块石2种, 都产在礁后潟湖的潮间带和很浅的潮下带(Reid et al., 1995; Planavsky and Ginsburg, 2009)。巴哈马海博内礁(Highborne Cay)的凝块石形成小型平坦的碳酸盐岩台地(Patterson, 2014)。虽然Patterson(2014)把这种微生物碳酸盐岩叫凝块石, 但在手标本上, 看不到典型的凝块结构。之所以被叫做凝块石, 可能是因为2个原因: (1)缺乏纹层结构, (2)有一些0.5~3 cm大小的洞。
采集自台地表面的标本保留了丝状藻及其外的矿物壳(图 4-c, 4-d)叫做矿物壳, 仅仅因为它包裹在丝状藻的周围; 其实它是丝状藻类诱导碳酸盐矿物结晶形成的。有很多丝状的孔洞(图 4-c, 4-d), 其周围有浅色的矿物壳, 所以它们是丝状藻类腐烂之后留下的, 属于模孔。模孔的直径34~43 μ m(图4-e)。形成这种模孔的丝状微生物可能是Dichothrix(Patterson, 2014)。这是一个以锐角行假分枝的丝状蓝藻属, 包括40多个种。照片中该丝状藻的确是以锐角分枝的, 应当属于Dichothrix。
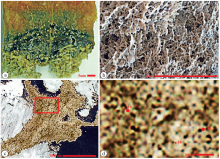
基里巴斯是太平洋的一个岛国, 岛上有很多高盐度湖泊。高盐湖的水很浅, 只有几米。其底部生长着一层橙黄色的微生物席, 微生物席的下面是微生物作用形成的碳酸盐岩(Arp et al., 2012; 本文图 5-a)。在手标本上就可以看到这种微生物碳酸盐岩呈白色不规则的网状结构(Arp et al., 2012; 本文图 5-b)。这种网状结构似乎归入凝块石比较合适。在显微照片上可以看到, 微生物碳酸盐岩由密集的模孔和其间的碳酸盐组成(图 5-c, 5-d)。模孔圆形和蠕虫形, 直径10 μ m和19 μ m。模孔之间的碳酸盐的厚度与模孔相似, 可达19 μ m; 它们是否为矿物壳, 难以确定。
 | 图 5 基里巴斯现代微生物碳酸盐岩的显微特征(据Arp等, 2012; 有修改) a— 修改自Arp等(2012)的图 6E, 采集自基里巴斯盐湖的手标本, 上层是橙色的微生物席, 中层是死的微生物, 下层白色的是网状的微生物碳酸盐岩; b— 修改自Arp等(2012)的图 13B, 采集自与a相同的地点, 为白色网状的微生物碳酸盐岩; c— 修改自Arp等(2012)的图 13F, 薄片显微照片, 标本采集自与a相同的地点, 单偏光; d— 是c的红色方框的放大, 显示微生物碳酸盐岩由模孔和其间的碳酸盐组成, 模孔圆形或蠕虫形, 直径有10 μ m和19 μ m 共2种Fig.5 Microfabrics of modern microbial carbonates in Kiribati(modified from Arp et al., 2012) |
以上3个例子说明, 现代自然环境的微生物碳酸盐岩的基本特征就是具有模孔。模孔的外面可能有矿物壳。矿物壳之间可能是被胶结的鲕粒或者砂屑。如果没有砂屑, 壳之间可能有原始孔隙, 叫剩余孔隙。剩余孔隙可以在后来的成岩作用中被较晚形成的矿物充填。
地质历史时期的成岩作用是否会把微生物碳酸盐岩的原始显微结构特征抹去?需要通过对地质历史时期的微生物碳酸盐岩的研究来确定。虽然前人对地质历史时期的微生物碳酸盐岩做了很多研究, 但是一般都不是以显微结构为主要内容的。作者虽然已经对大量的古代微生物碳酸盐岩进行了研究, 但限于篇幅在此只对中国新疆寒武系第三阶的一个剖面进行介绍。
寒武纪是显生宙的第1个纪, 是地球演化的一个重要时期, 寒武纪第三阶(521-> 517 Ma)是生物大爆发的主要时期(Zhang et al., 2014)。在世界范围内发现了很多寒武纪微生物岩。中国寒武纪微生物碳酸盐岩在华南、华北、塔里木三大板块都有发现。华南板块寒武纪微生物碳酸盐岩目前发现较少(王剑等, 1990; Chen et al., 2018)。华北板块分布较多, 保存好, 研究多(Chen et al., 2014; Ezaki et al., 2017; Wu et al., 2017; Yan et al., 2017)。塔里木板块寒武纪地层普遍白云石化, 微生物岩是重要的油气储集层, 得到较多的研究(You et al., 2014)。
以塔里木盆地阿克苏城西南80 km处的苏盖特布拉克剖面为例, 介绍寒武纪微生物岩的宏观、中观、微观特征。该剖面有出露很好的寒武系第三阶肖尔布拉克组。很多学者对该剖面肖尔布拉克组的微生物岩进行了研究(宋金民等, 2012; Bai et al., 2017; 邓世彪等, 2018; 郑剑锋等, 2019)。作者实测确定该组厚度187 m, 几乎全部由白云岩组成。根据宏观特征将该组分为5段。第Ⅰ 段, 黑色, 薄层状; 第Ⅱ 段, 黑色, 似薄层状或块状, 有蜂窝状溶洞; 第Ⅲ 段, 灰色, 纹层状; 第Ⅳ 段, 灰黄色, 丘状体和其间的薄层状深灰色白云岩(图6-a); 第V段, 灰黄色— 黑色, 中层状。在第Ⅲ 段、第V段露头上可见纹层构造, 其余地层露头上看不到任何微生物岩特征。在显微镜下可见, 第Ⅰ 段以纹层结构为主, 也有凝块结构。第Ⅱ 段, 以凝块结构为主。第Ⅲ 段, 是纹层结构与凝块结构的复合结构。第Ⅳ 段以凝块结构为主(图6-a, 6-b, 6-c)。第V段下部以纹层结构为主, 上段以砂屑结构和纹层结构为主。
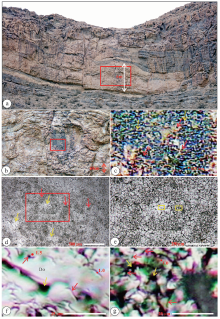
第Ⅳ 段厚度约24 m, 露头上见到由丘状体和其间的薄层状岩石构成。丘状体高度6~20 m。丘状体内部在露头上呈块状。在显微镜下, 可见丘状体内部的块状白云岩、丘状体之间的薄层状白云岩都具有凝块结构, 都属于凝块石。丘状体内部的凝块石由暗色不规则网状、斑点状、蠕虫状的“ 凝块” , 以及其间的浅色区域(称为亮斑)组成(图 6-d, 6-e)。斑点状“ 凝块” 的直径、蠕虫状“ 凝块” 的宽度、组成网状结构的“ 凝块” 的宽度为44~140 μ m; 亮斑的直径最大可达400 μ m。
 | 图 6 中国新疆塔里木盆地阿克苏市乌什县苏盖特布拉克村附近寒武系第三阶微生物岩的显微结构 a— 1个微生物丘的野外露头, 丘形, 内部块状, 由白云岩组成; 丘周围是层状白云岩地层; b— 是a的方框的放大; c— 是b的方框的放大, 见到岩石由模孔(红色箭头所指)和网状的“ 凝块” (黄色箭头所指)组成; d— 采集自b的标本的薄片, 由暗色的蠕虫状和网状的“ 凝块” (红色箭头所指)和其间的亮斑(黄色箭头所指)组成; e— 是d的方框的放大; f— 是e的左边黄色方框的放大, 为白云石晶体的一部分(Do), 其上有少数丝状的模孔(红色箭头所指); 模孔直径1~1.5 μ m; g— 是e的右边黄色方框的放大, 由较小的不规则形状的白云石晶体组成, 有较多的丝状模孔(红色箭头所指), 以及黑色的孔洞(黄色箭头所指)Fig.6 Photomicrographs of microfabrics of microbiailte of the Cambrian Stage 3 (Xiaoerbulake Formation) at Sugaitebulake, Aksu, Xinjiang, Northwest China |
对亮斑和暗色“ 凝块” 进行局部放大后(图 6-f, 6-g)可见, 亮斑和暗色“ 凝块” 都有丝状的小孔。因为形状呈规则的丝状, 认为它们是微生物的模孔。亮斑中模孔的密度比“ 凝块” 中小很多。亮斑内部和暗色“ 凝块” 内部的模孔特征相同, 多数呈稍微弯曲的管状, 长度可达15 μ m, 少数是圆形的, 直径为1 μ m。亮斑由宽度达到50 μ m的半自形白云石晶体组成。可见白云石晶体的边界。亮斑内模孔的间距较大, 所见最大间距可以达到15 μ m; 模孔之间是浅色的十分均匀的白云石。浅色的白云石胶结物使得亮斑呈浅色。暗色“ 凝块” 内部的模孔很密集, 间距一般只有1 μ m。
暗色凝块石内部模孔密集, 亮斑内部模孔稀疏, 代表当时微生物的分布不均匀。亮斑内的微生物十分稀疏。环境条件的改变, 使得微生物稀疏处的剩余孔洞没有被诱导结晶矿物充填, 后来被白云石晶体充填胶结。微生物密集处, 模孔密集。由于某种原因, 使得模孔之间的碳酸盐呈黑色。黑色的碳酸盐与黑色的模孔2个因素使得暗色凝块石区域是暗色的。
模孔的直径和长度与现代海洋的丝状蓝藻群动盖丝藻(Geitlerinema ionicum)(王梦梦等, 2017)相似。G. ionicum的丝体直径1.2~1.4 μ m。所以, 模孔可能是Geitlerinema ionicum留下的。
塔里木盆地寒武系第三阶微生物岩的例子说明即使是寒武纪的经过强烈白云石化改造的微生物岩也保留了微生物的模孔。无论前寒武纪还是显生宙, 除了白云石化的微生物岩地层外, 还有许多未遭受白云石化的地层, 它们的微生物岩原始结构应当有更好的保存。既然严重白云石化的微生物岩中模孔可以保存, 那么, 成岩作用影响较小的微生物岩地层中的模孔更应当能够被保存。
几个典型的现代微生物碳酸盐岩的实例以及塔里木盆地寒武系第三阶微生物岩的例子都说明, 现代和古代微生物碳酸盐岩的研究不能只在宏观、中观尺度上, 还要在显微和亚显微尺度上进行。
在宏观尺度上主要观察微生物岩的产状。宏观产状包括几个厘米厚的薄层状, 也可以是几米到几十米高的丘状。在中观尺度上主要观察微生物岩是纹层状的还是凝块状的, 观察其薄层的特征、其几厘米大小溶洞的特征、其窗格构造的特征。超过4 cm大的孔洞只能在手标本上观察, 因为在薄片中不能看到其全貌。在微观尺度上, 主要观察几十到几百微米大小的结构特征, 以及毫米和厘米级结构的显微特征。对凝块石而言, 在几百微米的尺度上, 俨然由几百微米厚的蠕虫状、网状的黑色“ 凝块” 和其间的亮斑组成。“ 凝块” 由较细的晶体组成, 一般由泥晶或粉晶组成; 亮斑是由较粗的晶体组成, 颜色较浅。在亚显微或几微米的尺度上, 几百微米尺度的“ 凝块” 是由模孔和其间的白云质组成, 亮斑其实是模孔稀疏的地方; “ 凝块” 其实是模孔较为密集的地方。
不同尺度的特征有不同的意义。几百微米尺度的结构, 反映的是微生物生长的不均匀, 是由底质状况、适宜微生物发育的环境条件的持续时间长短等因素决定的。而几微米尺度的结构, 反映的是微生物席的组成、水体沉积物的多少等特征。微生物席的组成受环境的物理化学条件控制。所以, 不同尺度的研究都有意义, 都是不可缺少的。
前人定义的凝块指的是孔洞之间的泥晶骨架。作者的研究说明, 以前的所谓凝块其实由更小的基本单元— — 模孔和矿物壳或者模孔和其间的矿物组成, 因此建议, 当模孔之间的碳酸盐无法区分是矿物壳或其他类型时, 笼统叫做基本凝块(proclot)。模孔是微生物留下的, 其大小和形状是规则的, 且与微生物的大小和形状一样。在成岩阶段, 模孔可能被胶结物充填, 也可能遭受溶蚀作用而扩大和形状变得不规则。虽然成岩作用可能形成溶蚀孔隙, 但溶蚀孔隙的形状和大小是不规则的。通过对形状和大小的分析, 可以区分模孔和溶蚀孔。
1)实验表明, 诱导结晶只在某些微生物的表面发生, 开始是形成点状碳酸盐颗粒, 后来形成碳酸盐矿物壳。一般而言, 微生物碳酸盐岩的基本组成单元是模孔和矿物壳, 还可以有孔隙和胶结物。
2)无论是现代还是古代的微生物碳酸盐岩, 模孔的存在是其基本特征。以3个典型的现代微生物碳酸盐岩的实例, 说明现代自然环境的微生物碳酸盐岩显微结构都有模孔, 可能大多数有矿物壳。塔里木盆地寒武系第三阶的微生物岩在显微尺度上由暗色“ 凝块” 和亮斑组成; 在亚显微尺度上, 暗色“ 凝块” 由模孔和其间的胶结物组成。
3)建议。微生物碳酸盐岩需要从4个尺度上进行研究: 宏观(> 10 cm)、中观(10~1 mm)、显微(50~1000 μ m)、亚显微(< 50 μ m)。以前的研究侧重于宏观和中观尺度, 对显微和亚显微尺度的研究开展较少。今后的微生物岩研究要对显微和亚显微结构的特征给予同样的重视。只有开展显微和亚显微尺度的研究, 才能全面认识微生物碳酸盐岩的特征, 进而分析其形成机制和形成环境。
(责任编辑 郑秀娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|