



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
光合作用微生物席主导的寒武系苗岭统崮山组均一石:以山东省泗水县圣水峪剖面为例*
[梅冥相1, 2  ]
]
]
|
|

作者简介 梅冥相,男,1965年生,教授,博士生导师,主要从事沉积学和地层学研究工作;E-mail: meimingxiang@263.net。
作为一种缺乏沉积构造的隐晶质块状泥晶所表征的非纹层状微生物碳酸盐岩类型,均一石自从命名之后就很少被描述或自古老的地层记录之中被识别出来,再加上缺乏现代实例,造成了对这种微生物碳酸盐岩类型研究的困难。来自于山东省泗水县圣水峪剖面寒武系苗岭统崮山组中的均一石,以厚层块状、缺乏沉积构造的泥晶灰岩为主,表现为生物层状的微生物礁,与相对较深水的中厚层中至深缓坡相的条带状泥晶灰岩和泥灰岩一起,构成较为典型的潮下型碳酸盐岩米级旋回,集中发育在淹没不整合型三级沉积层序顶部的强迫型海退体系域之中,代表着相对海平面下降阶段的沉积记录。在这些主导着生物层状微生物礁灰岩的均一石中,不均匀地高密度保存着致密缠绕以及较薄的泥晶壁所成型的丝状葛万菌物质体是其最为基本的特征,表明均一石形成于光合作用微生物席复杂的钙化作用。另外,在均一石的致密泥晶组构中,局部发育毫米级至厘米级大小而且形态极端不规则的亮晶显微管状组构。基于其特别的组构特征,将这样的组构解释为具有公共鞘的丝状蓝细菌即念珠菌菌落的钙化作用残余物,其致密泥晶组构可能代表着蓝细菌公共鞘的钙化作用残余物,其中的亮晶显微管则可能为蓝细菌丝状体的钙化作用残余物,而且在这样的组构之中还发育和共生着丝状葛万菌菌落,从而延伸了对蓝细菌菌落钙化作用残余物的认识和解释,并对具这种组构的海绵木乃伊解释提供了另外一种重要的研究镜像。因此,山东省泗水县圣水峪剖面寒武系苗岭统崮山组上部生物层状微生物礁体中的均一石,不仅提供了一个较为典型的寒武纪苗岭世不借助于二氧化碳浓缩作用机制(CCM机制)的蓝细菌钙化作用的实例,而且提供了一个在丝状蓝细菌主导的微生物席之中发育着念珠菌菌落的多重状微生物席的典型实例,从而拓宽了对显生宙最高的大气圈CO2含量以及较高的大气圈O2含量条件下的较为典型的方解石海中与寒武纪后生动物大爆发相平行的蓝细菌繁荣和钙化作用的了解。
About the author Mei Ming-Xiang,born in 1965, graduated from China University of Geosciences(Beijing) in 1993 with his Ph.D. degree. Now he is a professor at School of Earth Sciences and Resource,China University of Geosciences(Beijing),and is engaged in sedimentology and stratigraphy. E-mail: meimingxiang@263.net.
Microbialites have been defined as the organosedimentary deposits that have accreted as a result of a benthic microbial community trapping and binding sediment and/or forming the locus of mineral precipitation or teemed as the benthic microbial deposits. Microbial carbonates are understood as the calcified microbialite as well as the geological record of calcified bacterial-algal mats and biofilms or the most spectacular example of the MISSs(microbially-induced sedimentary structures). On the basis of the meso-structures,microbialites can be subdivided into four types,inlcuding stromatolites,thrombolites,leiolites and dendrolites. Leiolites were named relatively late and characterized by relatively structureless,aphanitic,mesostructures,lacking lamination or clotted structures. No examples of modern leiolite have been published and only few examples have been described or discerned in stratigraphic records since it was firstly named,which led to difficulties in the study of leiolites. Leiolites from the Miaolingian Gushan Formation at the Shengshuiyu section in Sishui,Shandong Province,are predominated by massive micrite and expressed as a biostromal microbial reef. They,together with the ribbon-like marls and micrite,make up the subtidal carbonate cycle belonging to the relatively deep-water deposits. Thus,a sedimentary succession of this type of subitidal carbonate cycles constitutes a forced regressive system tract of a third-order sequence of the Miaolingian Formation at the Shengshuiyu section,which represents a typical record of the falling stage of a third-order relative sea-level change. Within dense and massive micrite of the leiolites,a common and uneven high-density preservation of Girvanella is marked by densely tangled masses of fairly thin-walled,unbranched filaments or tubes,which indicates that these leiolites possibly resulted from the sophisticated calcification of cyanobacterial mats. In addition,a particular micro-fabric is marked by the microspar tubes of filaments within dense and dark micrites and makes up an irregular-shaped and mm-to cm-long clump. This micro-fabric is similar to the Lithocodium,described as the Lithocodium-like fabric,which can be regarded as colonies of calcified cyanobacteria. Importantly and interestingly,this micro-fabric was interpreted as the sponges including spicula sponges or non-spicula sponges in studies of microbial carbonates in recent years,which resulted in a highly different cognition. According to its fundamental feature, i.e. the microspar filaments or tubes within the dense and dark micrite,I deduced that it could be the residue of calcified Nostoc colonies with common sheathes. The microspar filaments or tubes might represent the calcified residues of filaments or tubes;the dark and dense micrite might have been formed by calcification of sheathes. This interpretation is further supported by its intergrowth with Girvanella. Leiolites predominating the biostromal microbial reefs of the Miaolingian Gushan Formation at the Shengshuiyu section not only provide an example of the Cambrian leiolites but also provide insights into the cyanobacterial calcification without the help of CCM mechanism and the sophisticated calcification of multiple cyanobacterial mats predominated by both the Girvanella and the residue of the Nostoc colonies. This study broadens further understanding of a particular calcification of cyanobacteria and a cyanobacterial blooms that are paralleled with the metazoan exploration in typical calcite sea during the Cambrian under a high atmosphere CO2 and O2.
微生物岩是指“ 底栖微生物群落捕获与黏结沉积物和矿物沉淀作用而形成的有机沉积物的沉积” (Burne and Moore, 1987), 被归为“ 底栖微生物沉积” (Riding, 1991a)。基于中等尺度的构造特征, 微生物岩可划分为4类: 纹层状的叠层石(Kalkowsky, 1908), 凝块状的凝块石(Aitken, 1967), 树枝状的树形石(Riding, 1991a)和贫乏构造的均一石(Braga et al., 1995)。此外, 纹理石和核形石也应该被纳入微生物岩的范畴(梅冥相, 2007), 就像鲕粒可以看作是“ 纹层状的有机沉积构造” 并遵循着微生物岩体系的一些形成特征(Diaz and Eberli, 2019)那样, 近年来的许多研究成果均支持这一观点(Tang et al., 2014; Han et al., 2015; 梅冥相等, 2019a, 2019b, 2020a)。然而, 微生物岩所具有的特征性微球粒聚合致密泥晶微组构, 常掩盖了它们确切而真实的形成机理, 对解释该机理造成了巨大的挑战(Burne and Moore, 1987; Riding, 1991a, 2000, 2011a; 梅冥相, 2007; Dupraz et al., 2009, 2011; Suosaari et al., 2019); 加之微生物岩具有37× 108年的发育历史(Nutman et al., 2016)、多样化的产出环境(Flannery et al., 2019)和多种矿物构成, 造成微生物岩是地质学领域一个长久的研究主题。
微生物碳酸盐岩是钙化的微生物岩, 为微生物席的建造物或微生物诱发的沉积构造(MISS; 梅冥相, 2011a, 2011b, 2014; Noffke and Awramik, 2013; Maisano et al., 2019)的钙化实例(Dupraz et al., 2011), 但缺乏可以类比的现代实例(Dupraz et al., 2011), 而且均一石自从命名以来就很少在地层记录之中被发现和识别。虽然在寒武系已经发现了一些实例(梅冥相等, 2019c, 2019e; 肖恩照等, 2020), 但由于均一石的块状泥晶构成与生物礁岩石— — 粘结岩(Embry and Klovan, 1971; Riding, 2002)最为相似, 而且与那些定义与成因存在着较大争议的碳酸盐泥丘(Pratt, 1995; 彭阳等, 1998; Wood, 2001; Riding, 2002; Schlager, 2003; 章雨旭等, 2005; Pomar and Hallock, 2008; Reijmer, 2016)也难以区分, 所以, 还存在着一些概念上的不确定性, 这进一步增加了对均一石研究的困难。
对均一石确切的驱动机制和响应机理研究目前还存在着许多不确定性, 表现在: (1)对应着寒武纪大气圈的CO2含量高于当前25倍(Berner and Kothavala, 2001), 当时大气圈的O2含量还存在着相对较低(Berner et al., 2007; Campbell and Allen, 2008; Lenton et al., 2018; Riding et al., 2019)与相对较高的旋回性变化(Large et al., 2019)等不同认识; (2)Large等(2019)认为苗岭世与芙蓉世之交(大致500 Ma)大气圈O2含量下降, 尽管该结论与大洋范围缺氧的解释(Gill et al., 2011)相一致, 但是又与Saltzman等(2011)所认为的伴随着SPICE碳同位素正异常相关联的O2含量上升(也几乎可以达到25%)这一结论存在着矛盾; (3)许多学者认为, 寒武纪相对较低的大气圈O2含量水平有利于激发厌氧新陈代谢作用, 从而有利于微生物碳酸盐岩的发育(Edwards et al., 2017; Lee and Riding, 2018; Riding et al., 2019), 这与Large 等(2019)的寒武纪相对较高的氧气含量、以及Saltzman 等(2011)所认为的伴随着SPICE碳同位素正异常相关联的氧气上升(也几乎可以达到25%)的结果和认识不一致。但是, 寒武纪时期与后生动物的寒武纪大爆发(Dove, 2010; Peters and Gaines, 2012)相平行的微生物碳酸盐岩的复苏(Riding, 1991a, 2000, 2006a, 2011a)、蓝细菌繁荣以及不需要借助于二氧化碳浓缩作用机制(CCM; Riding, 2006b; Kah and Riding, 2007)的蓝细菌钙化作用(Riding, 1991b, 1991c, 2000, 2006b, 2011b), 在近年来的研究中得到了进一步的证实, 因为在寒武纪核形石(Liu and Zhang, 2012; Han et al., 2015; Wilmeth et al., 2015; 梅冥相等, 2019a, 2019b, 2020a)、鲕粒(Liu and Zhang, 2012; 代明月等, 2014; 梅冥相等, 2020b)与微生物碳酸盐岩(齐永安等, 2017; Latif et al., 2019; 梅冥相等, 2017, 2019c, 2019e, 2020b; Mei et al., 2020; 肖恩照等, 2020)中, 均较为普遍地发育蓝细菌鞘的钙化化石。
山东省泗水县圣水峪剖面寒武系苗岭统崮山组为一个淹没不整合型三级沉积层序(Gó mez and Fernández-Ló pez, 1994; 梅冥相, 1996, 2010, 2011a; 马永生等, 2017)。在该三级沉积层序上部的强迫型海退体系域或晚期高水位体系域之中, 由厚层块状、贫乏构造的泥晶灰岩所主导的均一石(表现为生物层状的微生物礁)与相对较深水的中厚层中至深缓坡相的条带状泥晶灰岩和泥灰岩一起, 组成较为典型的潮下型碳酸盐岩米级旋回(Osleger, 1991; Mei et al., 2000)。在这些均一石的块状致密泥晶之中, 普遍发育致密缠绕、较薄的泥晶壁所成型的丝状葛万菌(Girvanella; Nicholson and Etheridge, 1878; Riding, 1977, 1991b, 1991c, 2000, 2006a, 2011b), 以及较为特别的致密泥晶中的亮晶显微管所表征的而且认识上存在巨大争议的石松藻(Lithocodium; Elliott, 1956; Riding, 1991b, 1991c; Cherchi and Schroeder, 2006)和石松藻状组构(Lithocodium-like fabric; Rameil et al., 2010; Schlagintweit and Bover-Arnal, 2013; Schmitt et al., 2019), 这为了解均一石形成时的微生物活动属性、石松藻状组构在寒武纪的确立及其合理的生物亲和性提供了较为罕见的实际材料; 同时, 也为拓宽与后生动物的寒武纪大爆发相平行的方解石海中的蓝细菌繁荣提供重要的思考途径和研究线索。笔者的目的在于: (1)展现一个较为典型的寒武系苗岭统均一石的宏观特征和显微构成; (2)阐释寒武系均一石可能的微生物席的原始生产者; (3)确立石松藻状组构在寒武纪的确立及其可能的念珠菌生物亲和性。
华北地台寒武系包括第二统上部、苗岭统和芙蓉统(彭善池, 2009; Peng et al., 2012; 彭善池和赵元龙, 2018), 与前寒武系之间为类似于北美地台(Peters and Gaines, 2012)的“ 巨型不整合面” (冯增昭等, 1990, 2004; Meng et al., 1997; 章宇旭, 2001; Lee and Chough, 2011; 马永生等, 2017; 梅冥相等, 2017, 2019a, 2019b, 2019c, 2019d, 2019e, 2020a, 2020b; Woo et al., 2019; Mei et al., 2020)。毛庄组、徐庄组、张夏组和崮山组大致构成了华北地台的苗岭统, 根据三叶虫化石带的年代地层学意义(彭善池, 2009; Peng et al., 2012), 崮山组大致对应苗岭统(彭善池和赵元龙, 2018)古丈阶。
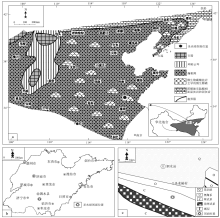
山东省泗水县圣水峪剖面(剖面位置见图 1)的崮山组为一个三级沉积层序, 发育在较为典型的淹没不整合面组成的层序界面(图 2; Schlager, 1989, 1998, 1999; Gó mez and Fernández-Ló pez, 1994; 梅冥相, 1996, 2010, 2011c; 马永生等, 2017; Latif et al., 2018; 王龙等, 2018; Riaz et al., 2019a, 2019b; Mei et al., 2020)之上, 以总体向上变浅的沉积序列为特征, 即从陆棚相泥页岩地层构成的凝缩段向上变浅为高水位体系域的鲕粒滩灰岩与厚层块状微生物礁所组成的序列(即CS+HST序列)。表征着生物层状微生物礁的厚层块状致密泥晶灰岩所代表的均一石灰岩(图 2-b, 2-c), 与深缓坡至中缓坡相的泥质条带泥灰岩和泥晶灰岩一起, 组成若干个潮下型碳酸盐岩米级旋回(Osleger, 1991; Mei et al., 2000)。这套厚度接近30 m的旋回序列构成了三级沉积层序的晚期高水位体系(LHST)或强迫型海退体系域(FRST; Hunt and Tucker, 1992; Helland-Hansen and Gjelberg, 1994; 梅冥相和杨欣德, 2000; Schlager and Warrlichw, 2009; 梅冥相, 2010; Desjardins et al., 2012; Samanta et al., 2016; Mei and Liu, 2017), 为文中研究的关键层位。该旋回序列代表着Schlager和Warrlichw(2009)所倡导的相对海平面下降阶段的沉积记录, 从早期高水位体系域上部的鲕粒滩向上演变为微生物礁的序列, 表现为与较慢的侵蚀作用和基准面下降速率相对应的较高的沉积物生产作用, 而且这样的向上得到增强而与海平面下降阶段相对应的碳酸盐岩序列, 其本身就可能表明了碳酸盐生产和沉积作用指向了光合作用新陈代谢的特征(Rameil et al., 2010; Schmitt et al., 2019)。更加显著的是, 在崮山组构成的三级沉积层序的晚期高水位体系域(或强迫型海退体系域)的形成时期, 华北地台发育着一个面积大于100× 104 km2的鲕粒滩, 其代表着寒武纪类型多样的碳酸盐台地类型中(Pratt et al., 2012)一种新的碳酸盐台地类型(图 1-a), 即鲕粒滩主导的浅海碳酸盐岩陆架, 向海一侧为缓坡, 向陆一侧为潮坪(马永生等, 2017; 梅冥相等, 2019a, 2019b, 2019c, 2019d, 2019e, 2020a, 2020b), 而且在大面积展布的鲕粒滩局部发育微生物碳酸盐主导的微生物礁(Riding, 2002; Pratt et al., 2012), 圣水峪剖面崮山组顶部的微生物礁就是一个较为典型的实例。
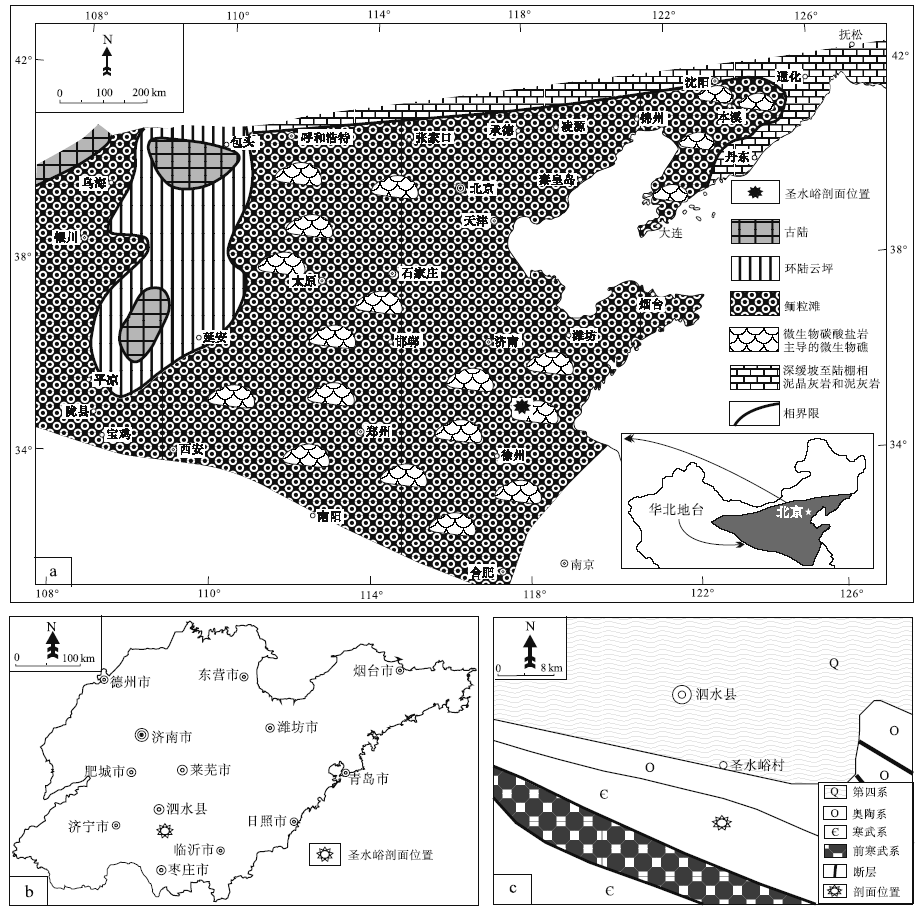 | 图 1 山东省泗水县寒武系崮山组圣水峪剖面位置 a— 为崮山组构成的三级层序的强迫型海退体系域沉积期的沉积相, 修改自冯增昭等(1990, 2004); b— 山东省的轮廓; c— 泗水县附近的地质略图Fig.1 Diagrams showing location of Shengshuiyu section for the Cambrian Gushan Formation in Sishui County of Shandong Province |
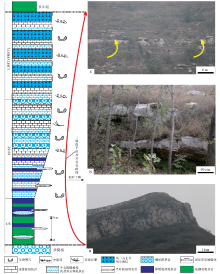
 | 图 2 山东省泗水县圣水峪剖面寒武系崮山组沉积序列 a— 崮山组顶部的块状鲕粒滩相灰岩; b— 崮山组上部的厚层块状均一石灰岩与深缓坡相条带状泥灰岩互层组成的潮下型米级旋回; c— 表示崮山组与上覆的长山组之间的淹没不整合面组成的三级层序界面, 以陆棚相至深缓坡相条带状泥灰岩和泥晶灰岩直接覆盖在浅缓坡相的块状均一石灰岩之上为特征。CS代表三级沉积层序的凝缩段, EHST代表早期高水位体系域, LHST代表晚期高水位体系域并等同于强迫型海退体系域(FRST)Fig.2 Diagram showing depositional succession for the Gushan Formation of Cambrian at Shengshuiyu section in Sishui County of Shandong Province |
如图 2-b、2-c以及图 3-a所示的厚层块状隐晶质泥晶灰岩, 沉积构造不发育, 从而符合均一石(Braga et al., 1995)的定义。这些均一石常与较深水的条带状中厚层泥灰岩一起组成潮下型碳酸盐岩米级旋回(Osleger, 1991; Mei et al., 2000; 梅冥相, 2011d), 而且在下伏的鲕粒滩相灰岩中与均一石灰岩相对应的位置发育厚层块状鲕粒灰岩层, 间接地表明厚层块状隐晶质泥晶灰岩主导的均一石灰岩应为浅水高能的潮下带沉积。这些厚层块状隐晶质泥晶灰岩主导的均一石灰岩构成了生物层状(Kershaw, 1994)的微生物礁体(Riding, 2002; Pratt et al., 2012; Lee and Riding, 2018), 从而区别于一般的泥晶灰岩。在均一石灰岩中, 不均匀分布着成因不明的泥质条带, 其以缝合线状的形式发育在块状泥晶灰岩主导的均一石灰岩之中(图 3-b, 3-c)。
 | 图 3 山东省泗水县圣水峪剖面寒武系崮山组均一石的宏观特征 a— 浅缓坡相厚层块状均一石灰岩与较深水的中至深缓坡相条带状泥灰岩和钙质泥页岩组成的潮下型米级旋回; b, c— 均一石灰岩的近照, 表现为含泥质条带的块状泥晶灰岩Fig.3 Images showing macroscopic feature of leiolite for the Cambrian Gushan Formation at Shengshuiyu section in Sishui County of Shandong Province |
需要强调的是, 均一石曾因成为“ 隐秘微生物(cryptomicrobial)” 的同义语而被归为凝块石(Kennard and James, 1986)。均一石与其他类型的非纹层状微生物碳酸盐岩(如凝块石和树形石)的主要区别是其具有隐晶质、贫乏构造的泥晶灰岩构成的块状构造, 与生物礁岩石— — 粘结岩(Embry and Klovan, 1971; Riding, 2002)最为相似, 而且与那些定义和成因均存在着较大争议的碳酸盐泥丘(Pratt, 1995; 彭阳等, 1998; Wood, 2001; Schlager, 2003; 章雨旭等, 2005; Pomar and Hallock, 2008; Reijmer, 2016)一样是由块状泥晶构成。因此, 上述这些不同术语和概念之间的严格界定, 将是一个尖锐而且有待于进一步澄清的沉积学问题。
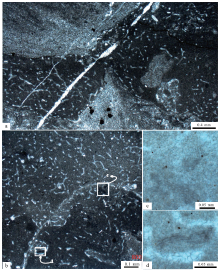
在显微镜下, 均一石的块状泥晶由突出的暗色致密泥晶和少量微亮晶构成(图 4-a, 4-d), 可见到少量的三叶虫化石, 与其共生的是较为普遍的微亮晶, 不发育叠层石的纹层状组构(Riding, 2000, 2011a; Dupraz et al., 2011)以及凝块石的凝块状组构(Aitken, 1967; Riding, 1991a, 2000, 2011a; Dupraz et al., 2011; 梅冥相, 2019d; Mei et al., 2020)。与暗色致密泥晶共生的碳酸盐微亮晶(图 4-a, 4-d)具有明显的非均质性, 总体上表现为被极薄的(小于1 μ m)而且不规则弯曲的暗色泥晶薄膜所分隔的方解石微亮晶组构, 与多被描述在中生代的孢网菌(Bacinella; Radoič ić , 1959; Riding, 1991a, 1991b)或孢网菌状组构(Bacinella-like fabric或“ Bacinelloid fabric” ; Rameil et al., 2010; Schlagintweit and Bover-Arnal, 2013; Schmitt et al., 2019)较为相似。在图 4-a中部几乎贯穿整个图像的宽度为20~30 μ m的杂色线状体, 也表现在高倍显微镜照片图 4-b和4-c之中, 可能就是图 3-b和3-c之中的那些缝合线状的泥质条带的显微组构表现, 应该为压实作用的产物。
 | 图 4 山东省泗水县圣水峪剖面寒武系崮山组均一石的微观特征 a, d— 低倍显微镜照片, 均一石由暗色致密泥晶和少量微亮晶构成, 可见少量三叶虫化石; b, c, e— 高倍显微镜照片, 分别放大于相应的低倍显微镜照片中的长方形框所表示的部分, 表现出分布不均匀而且高密度保存的丝状葛万菌, 黑色小点可能代表着黄铁矿晶体的残余物Fig.4 Images showing generally microscopic feature of leiolite for the Cambrian Gushan Formation at Shengshuiyu section in Sishui County of Shandong Province |
在高倍显微镜下, 在均一石的暗色致密泥晶之中发现了较为普遍的葛万菌(Girvanella; Nicholson and Etheridge, 1878)化石(图 4-b, 4-c, 4-e), 总体产出为由松散或致密缠绕的较薄的壁所成型的不分叉管状物的物质体, 也包括一些非缠绕、不分叉丝状体的物质体(图 4-b, 4-c), 以及一些电缆线状的不分叉丝状体的物质体(图 4-e)。这些丝状或管状微生物化石, 为较薄(1~2 μ m)而且均匀的暗色泥晶壁所成型, 直径10 μ m左右, 长度一般为数十至数百微米, 最长可以达到毫米级别(图 4-e), 它们构成了一个特征的不分叉丝状体的葛万菌(Girvanella)化石组合(Riding, 1991a, 1991b)。这些丝状化石的物质体多为长条状和纹层状, 宽度多为30~50 μ m, 长度为数百微米乃至毫米级别, 高密度发育, 占据着接近一半的体积, 表明它们是均一石的主要建造物; 而少数与其共生的三叶虫化石的出现(图 4-a, 4-d, 4-e), 表明这些丝状微生物在生长期间并不排斥三叶虫。
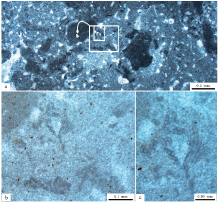
进一步观察表明, 在发育着较多而且可以类比于孢网菌(Bacinella; Radoič ić , 1959; Riding, 1991a, 1991b)或孢网菌状组构(Bacinella-like fabric或“ Bacinelloid fabric” ; Rameil et al., 2010; Schlagintweit and Bover-Arnal, 2013; Schmitt et al., 2019)的微亮晶构成的暗色致密泥晶之中(图 5-a), 残余着更多葛万菌(Girvanella; Nicholson and Etheridge, 1878)化石(图 5-b, 5-c), 即松散或致密缠绕的较薄的壁成型的不分叉管状物的物质体。这些物质体多为形态极端不规则的拉长状物质体, 与其生长基质(致密泥晶和微亮晶基质)之间的边界较为清晰, 说明这些不分叉的丝状体为丝状微生物化石组成的菌落。但是, 棱角状的总体形态以及锯齿状的过渡性边界, 表明这些物质体不是外来的内碎屑物质, 应该是原地堆积和生长的产物。构成这些不分叉的丝状物质体的丝状化石, 包括高度缠绕到不缠绕的形态, 几乎占据着均一石的泥晶质50%左右的面积或体积。虽然也曾经在华北地台寒武纪芙蓉世的均一石中报道过类似的现象(梅冥相等, 2019c, 2019e; 肖恩照等, 2020), 但在寒武纪均一石中发现如此高密度保存的丝状微生物化石菌落还是第1次。
 | 图 5 山东省泗水县圣水峪剖面寒武系崮山组均一石的致密泥晶和可能的孢网菌状组构 a— 低倍显微镜照片, 均一石由暗色致密泥晶和少量微亮晶构成, 微亮晶构成代表着可能的孢网菌状组构; b, c— 高倍显微镜照片, 分别放大于照片a中的长方形框所表示的部分, 表现出不均匀分布而且高密度保存的丝状葛万菌, 被暗色泥晶薄膜分隔的微亮晶可能代表着孢网菌状组构, 黑色小点可能代表着黄铁矿晶体的残余物Fig.5 Images showing dense and dark micrites as well as few possible Bacinella-like fabric within leiolite for the Cambrian Gushan Formation at Shengshuiyu section in Sishui County of Shandong Province |
在圣水峪剖面崮山组上部主导着生物层状微生物礁的均一石的泥晶中, 发现了亮晶显微管状组构(图 6), 其构成厘米级别的、形态极端不规则的物质体(图 6-a, 6-b), 与普通发育着丝状微生物化石的致密泥晶和微亮晶组构共生, 二者之间具有1个较为清晰的锯齿状边界。暗色致密泥晶以颜色更加偏向黑色而区别于周围的致密泥晶和微亮晶组构, 并组成厘米级别、形状不规则的暗色致密泥晶团块; 而亮晶显微管由微弯曲的亮晶小管所构成, 直径为5~10 μ m, 长度为数十微米至数百微米, 多为独立产出且很少相互连接, 组成一种不规则的网状结壳, 在局部也见到丝状葛万菌化石的小型物质体(图 6-b, 6-c, 6-d)。这种沉积组构类似于石松藻(Lithocodium; Elliott, 1956; Riding, 1991b, 1991c; Cherchi and Schroeder, 2006; Vé drine et al., 2007)或石松藻状组构(Lithocodium-like fabric; Rameil et al., 2010; Schlagintweit and Bover-Arnal, 2013; Schmitt et al., 2019), 常与上文所提及到的孢网菌(Bacinella; Radoič ić , 1959; Riding, 1991b, 1991c)或孢网菌状组构(Bacinella-like fabric或“ Bacinelloid fabric” ; Rameil et al., 2010; Schlagintweit and Bover-Arnal, 2013; Schmitt et al., 2019)的微亮晶组构共生, 发育在中生代的微生物碳酸盐岩以及核形石的皮层(Vé drine et al., 2007)之中, 尤其是在晚侏罗纪和白垩纪的浅海环境中较为普遍(Cherchi and Schroeder, 2006; Vé drine et al., 2007; Rameil et al., 2010; Schlagintweit and Bover-Arnal, 2013; Schmitt et al., 2019)。它们被描述为一个存在着巨大疑问的钙化蓝细菌化石组合, 即石松藻(Lithocodium)-孢网菌(Bacinella)组合(Riding, 1991b, 1991c), 在华北地台寒武纪芙蓉世(梅冥相等, 2017)和苗岭世(梅冥相等, 2019e)的叠层石中也曾经描述过这样的组构。
 | 图 6 山东省泗水县圣水峪剖面寒武系崮山组均一石的致密泥晶中较为典型的石松藻状组构 a, b— 低倍显微镜照片, 均一石的暗色致密泥晶中较为典型的不规则形态、厘米级别大小的石松藻状组构, 即暗色致密泥晶中的亮晶显微管组构; c, d— 高倍显微镜照片, 分别放大于照片b中的长方形框所表示的部分, 表现出不均匀分布而且高密度保存的丝状葛万菌, 以及亮晶显微管的亮晶方解石构成, 黑色小点可能代表着黄铁矿晶体的残余物Fig.6 Images showing typically possible Lithocodium-like fabric within leiolite for the Cambrian Gushan Formation at Shengshuiyu section in Sishui County of Shandong Province |
进一步观察表明, 暗色致密泥晶中的亮晶显微管状组构, 表征着可能的石松藻或石松藻状组构之中高密度保存的丝状葛万菌化石的小型凝块或团块(图 7)。这些葛万菌凝块或团块形态极不规则, 大小不一, 多为数十微米到数百微米, 锯齿状的边界、棱角状的形态以及漂浮状的产出特征表明它们不是外来的碎屑物, 而应该是原地堆积的产物; 也表明这样的可以归为石松藻或石松藻状组构的特别的沉积组构, 与丝状葛万菌代表的微生物菌落之间具有良好的共生关系。构成丝状蓝细菌物质体的丝状葛万菌组合的化石, 包括高度缠绕以及不缠绕的丝状体, 其中丝状体为较薄(1~2 μ m)而且较为均匀的暗色泥晶壁所成型, 直径为10 μ m左右, 长度在数十至数百微米之间, 而且在丝状葛万菌物质体之间还穿插着亮晶显微管。
 | 图 7 山东省泗水县圣水峪剖面寒武系崮山组均一石中较为典型的石松藻状组构中的丝状葛万菌物质体 a— 低倍显微镜照片, 均一石的暗色致密泥晶构成之中较为典型的不规则形态、厘米级别大小的石松藻状组构, 即暗色致密泥晶中的亮晶显微管组构; b, c— 高倍显微镜照片, 分别放大于照片a中的长方形框所表示的部分, 表现出不均匀分布而且高密度保存的丝状葛万菌以及亮晶显微管的亮晶方解石构成; d, e— 分别放大于照片b与c中的箭头所示的葛万菌物质体或菌落, 亮晶显微管由非晶质方解石亮晶构成。高倍显微镜照片中的黑色小点可能代表着黄铁矿晶体的残余物Fig.7 Images showing typical masses of filamental Girvanella within possible Lithocodium-like fabric in leiolites for the Cambrian Gushan Formation at Shengshuiyu section in Sishui County of Shandong Province |
更加特别的是, 由亮晶显微管状组构所表征的石松藻或石松藻状组构, 与孢网菌或孢网菌状组构的微亮晶组构相互共生, 组成复杂的致密泥晶与微亮晶组构(图 8-a), 说明这2种类型的沉积组构具有密切的共生关系, 形成一种不规则状开放式的泥晶网状体的结壳状物质体(Riding, 1991b, 1991c)。在其中也残留着小型的丝状葛万菌物质体(图 8-b, 8-c), 即不分叉的丝状或管状微生物化石的物质体, 其模糊的边界和大小不一的漂浮状产出特征, 表现出原地生长或堆积的特点。
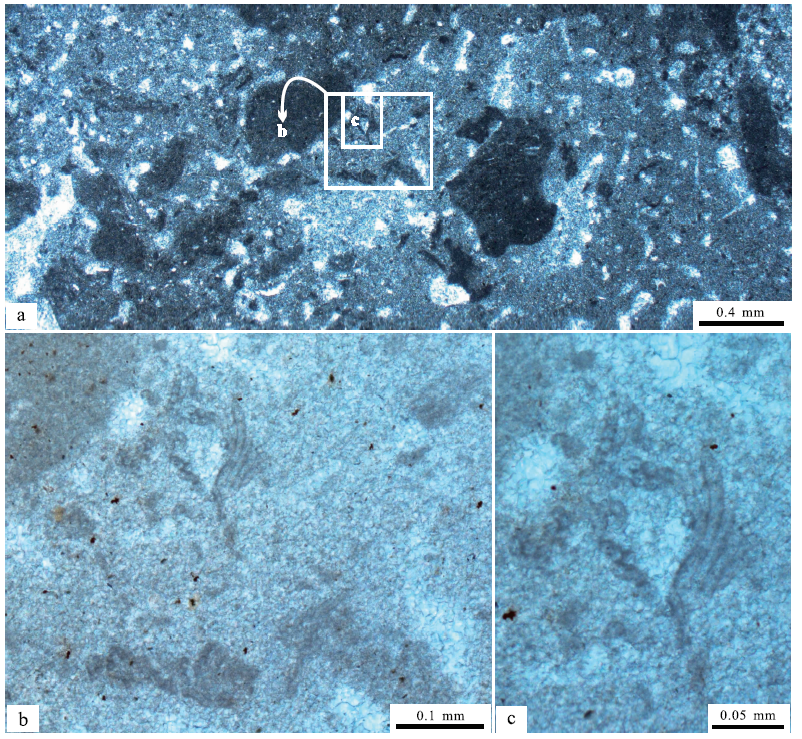 | 图 8 山东省泗水县圣水峪剖面寒武系崮山组均一石的致密泥晶和可能的石松藻和孢网菌状组构 a— 低倍显微镜照片, 均一石由暗色致密泥晶和少量微亮晶构成, 被暗色泥晶薄膜所分隔的微亮晶代表着可能的孢网菌状组构, 孢网菌状组构以及致密泥晶中的亮晶显微管代表着可能的石松藻状组构, 黑色泥晶团块代表着特别的凝块; b, c— 高倍显微镜照片, 分别放大于照片a中的长方形框所表示的部分, 表现出不均匀分布而且高密度保存的丝状葛万菌, 可见被暗色泥晶薄膜分隔的微亮晶代表着可能的孢网菌状组构, 黑色小点可能代表着黄铁矿晶体的残余物Fig.8 Images showing possible Lithocodium-like and Bacinella-like fabrics within leiolite for the Cambrian Gushan Formation at Shengshuiyu section in Sishui County of Shandong Province |
类似于致密泥晶中的由亮晶显微管状组构(图 6至图 8)所代表的可能的石松藻或石松藻状组构, 与北美地台的寒武纪芙蓉世叠层石(Coulson and Brand, 2016)和海绵生物礁(Lee et al., 2019)、华北地台的寒武纪芙蓉世的迷宫状凝块石组成的微生物礁(Chen et al., 2014; Lee et al., 2014, 2015, 2016; Lee and Riding, 2018)中曾经描述过的沉积组构较为相似, 只是这些学者将这样的沉积组构解释为海绵木乃伊, 而且还存在骨针海绵(Chen et al., 2014; Lee et al., 2014, 2015)与非骨针角质海绵(Lee et al., 2016, 2019; Lee and Riding, 2018)的不同认识。因此, 以下问题将成为关键的沉积学问题并成为文中讨论的核心内容: (1)与孢网菌状组构一样, 石松藻状组构能否发育在寒武纪; (2)石松藻状组构究竟是钙化蓝细菌菌落的残余物还是海绵木乃伊; (3)石松藻状组构如果解释为蓝细菌菌落的钙化作用残余物, 那么可能的蓝细菌亲和性的相对合理的答案又是什么。
山东省泗水县圣水峪剖面苗岭统崮山组为一个较为典型的淹没不整合型三级沉积层序(图 2; Schlager, 1989, 1998, 1999; Gó mez et al., 1994; 梅冥相, 1996, 2010, 2011b; 马永生等, 2017), 单层厚度为50~100 cm的浅缓坡相厚层块状泥晶灰岩主导的均一石灰岩, 与中厚层较深水的中至深缓坡相泥质条带泥灰岩和泥晶灰岩一起组成潮下型碳酸盐岩米级旋回(Osleger, 1991; Mei et al., 2000)。这些潮下型碳酸盐岩米级旋回表现出下部深水单元变薄、上部浅水单元变厚的基本特征, 组成一个总体向上变浅的旋回序列并构成三级层序强迫型海退体系域(Hunt and Tucker, 1992; Helland-Hansen and Gjelberg, 1994; 梅冥相和杨欣德, 2000; Schlager and Warrlichw, 2009; 梅冥相, 2010; Desjardins et al., 2012; Samanta et al., 2016; Mei and Liu, 2017), 进一步表明由致密泥晶主导的均一石构成的生物层状(Kershaw, 1994)微生物礁(Riding, 2002; Pratt et al., 2012; Lee and Riding, 2018)的沉积样式, 代表着海平面下降阶段的沉积记录(Schlager and Warrlichw, 2009)。其将碳酸盐生产和沉积作用指向光合作用新陈代谢的基本特征(Rameil et al., 2010; Schmitt et al., 2019), 表明这些均一石所主导的生物层状的微生物礁体占据着浅水热带工厂(Schlager, 2003; Reijmer, 2016; Michel et al., 2019; 颜佳新等, 2019)位置(因为局部发育在大片分布的鲕粒滩之中, 见图 1-a)的碳酸盐泥丘工厂或M-工厂。
在这些构成生物层状微生物礁(图 2-b, 2-c; 图 3-a)的均一石的致密泥晶之中, 至少存在着2种典型的、富有争议的钙化蓝细菌构成(图 4至图 8): (1)最为普遍的是葛万菌(Girvanella; Nicholson and Etheridge, 1878)化石代表的丝状蓝细菌, 这是较为肯定地可以类比近代织线菌(Plectonema)的丝状蓝细菌化石(Riding, 1977, 1991b, 1991c, 2000); (2)可能的孢网菌以及可能的石松藻代表的石松藻(Lithocodium; Elliott, 1956)-孢网菌(Bacinella; Radoič ić , 1959)组合, 代表存在着有较大疑问的钙化蓝细菌(Riding, 1991b, 1991c)。
以丝状不分叉的丝状体或管状物为特征的葛万菌(Girvanella), 在其命名的时候先被当作有孔虫(Nicholson and Etheridge, 1878)。Bornemann(1886)首先认识到它的蓝细菌属性, Pollock(1918)第1次将其识别为钙化的鞘, 后来又曾被认为是钙质海绵、绿藻和红藻, Riding(1977)则将其类比于现代或近代织线菌(Plectonema)的钙化鞘化石, 该解释相对较为肯定而且一直沿用至今(Riding, 1991a, 1991b, 2000, 2006b, 2011b; Liu and Zhang, 2012; Liu et al., 2016; 梅冥相, 2017, 2019a, 2019b, 2019c, 2019d, 2020a, 2020b; Mei et al., 2020); 另外, 类似于葛万菌(Girvanella)的缠绕状丝状蓝细菌鞘化石, 还被类比于现代的伪枝菌(Scytonema; 齐永安等, 2017)。就像图 4至图 8所示的葛万菌组合的丝状化石物质体那样, 除了高度缠绕的不分叉丝状体或管状物组成的较为典型的葛万菌(Girvanella), 还包含着不缠绕而且不分叉的可能代表着一种新类型而且可以类比于现代第谷菌(Tychonema; De los Rí os et al., 2015)的化石(图 4-b; 图 5-b, 5-c; 图 6-d; 图 7-e; 图 8-b, 8-c), 局部还见到直线状的和电缆线状的束线菌(Subtifioria, 图 4-e)之类的化石。丝状蓝细菌化石物质体(或菌落)表现为大小不一而且边缘多为锯齿状、形态为极端不规则的棱角状的致密泥晶体, 并占据着均一石几乎50%的面积和体积, 表明它们是原地堆积与生长的残余物。由于葛万菌组合的化石属于较为肯定地类比于现代蓝细菌的化石, 因此就可以较为肯定地推断出, 构成圣水峪剖面的崮山组均一石是较为典型的丝状蓝细菌主导的微生物席的钙化作用残余物(Stal, 2012)。这些丝状蓝细菌化石的最基本特征是具有较薄(1~2 μ m)而且很均匀的暗色致密泥晶壁, 并被解释为在大气圈CO2含量下降到10PAL(现代大气圈水平)条件下借助于CO2浓缩机制(CCM机制)的蓝细菌活体鞘的钙化作用(一种特别的生态生理学响应机制; Riding, 2006b; Kah and Riding, 2007)残余物。蓝细菌的鞘是含有生物遮光剂(如伪枝藻素)之类的特别的蛋白质而成为结构化的细胞外聚合物质(EPS), 但是在寒武纪尤其是在苗岭世, 大气圈的CO2含量几乎达到25PAL, 所以蓝细菌的活体鞘钙化作用不需要借助于CCM机制, 就像前寒武纪早期(12× 108年之前)的情况那样, 从而成为一个特例(Riding, 2011b)。尽管认为寒武纪较高的大气圈CO2含量有利于激发厌氧新陈代谢作用, 从而有益于微生物碳酸盐岩的发育(Edwards et al., 2017; Lee and Riding, 2018; Riding et al., 2019), 但是, 伴随着微生物碳酸盐岩的寒武纪复苏(Riding, 1991a, 1991b, 1991c, 2000, 2006a, 2011a)所出现的蓝细菌繁荣和普遍的蓝细菌钙化作用(Riding, 1991a, 1991b, 1991c, 2000, 2006b, 2011b), 不仅表现为在寒武纪的核形石(Liu and Zhang, 2012; Han et al., 2015; Wilmeth et al., 2015; 梅冥相等, 2019a, 2019b, 2020a)、鲕粒(Liu and Zhang, 2012; 代明月等, 2014; 梅冥相等, 2020b)与微生物碳酸盐岩(齐永安等, 2017; Latif et al., 2019; 梅冥相等, 2017, 2019c, 2019d, 2019e, 2020b; Mei et al., 2020; 肖恩照等, 2020)中较为普遍地发育着蓝细菌鞘的钙化化石, 而且发生在与基因控制的生物矿化作用(Perry et al., 2007; Dupraz et al., 2009; 梅冥相, 2012)得到进化而代表的寒武纪后生动物大爆发(Dove, 2010; Peters and Gaines, 2012)相平行的方解石海(Sandberg, 1983; Hardie, 1996; Balthasar and Cusack, 2015; Kiessling, 2015; Vulpius and Kiessling, 2018)之中, 这可能得益于寒武纪具有较高的大气圈O2含量(Saltzman et al., 2011; Large et al., 2019), 还可能得益于寒武系与前寒武系之间的巨型不整合面(Peters and Gaines, 2012)所表征的长时间风化作用所激发的海水中的Ca2+浓度突然从新元古代上升到寒武纪期间的显生宙最大值。再者, 蓝细菌CCM进化的时间进程也不清楚, 其可能进化在新元古代晚期或者在石炭纪的低CO2幕之中, 并通过横向基因转移传播到已经得到确立的主要分支中, 也可能是促进蓝细菌钙化作用的CCM机制在寒武纪确实已经出现并得到进化(Raven, 2012)。
将图 6至图 8中的 “ 暗色致密泥晶中的亮晶显微管状组构” 类比于石松藻(Lithocodium aggregatumElliott(1956)), 而且将 “ 暗色泥晶薄膜(宽度小于1 μ m)分隔的微亮晶组构” 类比于孢网菌(Bacinella; Radoič ić , 1959), 主要是基于形态学特征。实际上, 石松藻(Lithocodium)首先被Elliott(1956)描述在伊拉克 “ 下白垩统” 的灰岩中, 主要基于其与松藻Boueina属的气孔状纹层的比较, 所以在命名的时候被当作松藻的一种类型, 后来被归为存在着疑问的钙化蓝细菌(Riding, 1991b, 1991c)。这种毫米级别、形态不规则的填充在暗色致密泥晶基质中的不规则的微亮晶方解石填充的丝状体网状物所表征的石松藻, 曾经被解释为有孔虫(Schmid and Leinfelder, 1996; Schmid, 1996), 后来被解释为生物亲和性还有待于进一步调查的一种钙化蓝细菌菌落(Cherchi and Schroeder, 2006), 而且进一步描述为石松藻状组构(Lithocodium-like fabric; Rameil et al., 2010), 最终被认为是“ 生长态型确是表现出与特定的叠层石生长样式类型特别相似的结壳状微生物” 。那些被暗色泥晶薄膜(宽度小于1 μ m)分隔的微亮晶组构, 类比于孢网菌(Bacinella; Radoič ić , 1959), 如图 4至图 8所示(尤其是较好地表现在图 4-a至4-d、5-a、6-a、7-a、8-a所示的低倍显微照片之中, 也较好的表现在图 8-b、8-c的高倍显微照片之中), 代表着一种不规则的多孔状微亮晶结壳, 曾经被归属为一种绿藻(Schlagintweit et al., 2010), 后来又被几乎相同的作者解释为表现出与现代蓝细菌微生物席具有一些相似性的沉积组构, 并进一步定义和描述为孢网菌状组构(Bacinella-like fabric; Schlagintweit and Bover-Arnal, 2013; Schmitt et al., 2019)。所以说, 自从它们命名之后, 从一个存在着疑问的石松藻(Lithocodium)-孢网菌(Bacinella)钙化蓝细菌组合(Riding, 1991b, 1991c), 最终演变为相应的结壳状微生物组构而被分别称为石松藻状组构(Lithocodium-like fabric; Rameil et al., 2010)和孢网菌状组构(Bacinella-like fabric; Schlagintweit and Bover-Arnal, 2013; Schmitt et al., 2019), 本身就代表着重要的认识与概念进步。这样的概念进步也得到了进一步支持: (1)这2种组构在瑞士牛津期灰岩形成一种多孔网状的低能核形石皮层(Vé drine et al., 2007), 表现出明显的微生物结壳的特征; (2)在石松藻状组构中确实包含着有孔虫, 而且包围着真正的海绵骨针网状物所代表的海绵木乃伊(Cherchi and Schroeder, 2006), 从而否决了将其解释为海绵和有孔虫的认识(Schmid and Leinfelder, 1996; Schmid, 1996)。但是, 这样的沉积组构是否能够运用到寒武纪的微生物碳酸盐岩研究之中, 确实存在着巨大的挑战。
而在针对古生代叠层石等相关的微生物碳酸盐岩的研究中, 许多学者将类似石松藻状组构(Lithocodium-like fabric; Rameil et al., 2010)的沉积组构解释为海绵动物的钙化残余物(海绵木乃伊)。这样的解释首次出现在中国安徽下奥陶统红花园组微生物到后生动物生物礁转变的研究之中(Adachi et al., 2009), 后来被运用到寒武纪尤其是芙蓉统的微生物礁的研究之中, 而且产生了2种不同的认识: (1)海绵骨针网状物(甚至还是硅质海绵骨针网状物)(Hong et al., 2012, 2014, 2016; Chen et al., 2014; Lee et al., 2014, 2015; Coulson and Brand, 2016); (2)基于Luo和Reitner(2014, 2015)的概念而解释为非骨针角质海绵的钙化作用残余物(Lee et al., 2016, 2019; Lee and Riding, 2018)。梅冥相等(2017)可能率先对“ 石松藻(Lithocodium)状组构” 的海绵动物亲和性的解释提出了质疑, 将其解释和推测为“ 石松藻(Lithocodium)状的钙化蓝细菌” , 类似于现代湖泊中能够分泌大量细胞外聚合物质的颤菌(Oscillatoria)和蓆菌(Phormidium)等丝状蓝细菌所主导的微生物席的钙化作用产物(Voorhies et al., 2012), 或者更有可能类比于现代湖泊微生物席中的薄鞘丝菌(Leptolyngbya)或假鱼腥菌(Pseudanabaena)之类的丝状蓝细菌(Voorhies et al., 2012; De los Rí os et al., 2015; Kaź mierczak et al., 2015)的铸模或残余物。
尽管许多细节问题还需要进一步研究, 暗色致密泥晶中的显微管状组构所表征的石松藻(Lithocodium; Elliott, 1956; Riding, 1991b, 1991c)或石松藻状组构(Lithocodium-like fabric; Cherchi and Schroeder, 2006; Vé drine et al., 2007; Rameil et al., 2010)(图 6至图 8), 确实应该解释为具有公共鞘、而且是不分叉的丝状蓝细菌的钙化作用产物。念珠菌(Nostoc)(Potts, 1997; Wright et al., 2005; Soule, 2009; Mohr et al., 2011; Helm and Potts, 2012)正好符合这一基本特征, 这个推断和认识主要基于以下几个方面的考虑。
1)暗色致密泥晶基质的形成, 应该解释为丝状蓝细菌菌落的公共鞘的钙化作用产物, 因为蓝细菌的鞘是结构化的细胞外聚合物质(EPS), 而且是含有遮光剂之类蛋白质的EPS(Dupraz et al., 2009, 2011; Decho, 2010, 2011; Stal, 2012; Tourney and Ngwenya, 2014; Decho and Gutierrez, 2017)。这一点与成型丝状葛万菌较薄(1~2 μ m)、均匀的暗色泥晶壁(图 4-b, 4-c, 4-e; 图 5-b, 5-c; 图 6-c, 6-d; 图 7-b, 7-c, 7-d, 7-e; 图 8-b, 8-c)可能的活体鞘的钙化作用机制(Riding, 1991b, 1991c, 2006b, 2011b; Kah and Riding, 2007)相似, 念珠菌(Nostoc)丝状体的公共鞘同样分泌伪枝藻素(Scytonemin)之类的生物碱遮光剂(Wright et al., 2005; Soule, 2009; Helm and Potts, 2012), 二者均由暗色致密泥晶构成, 明显区别于那些形成均一石的较厚的微生物席中非结构化的EPS化作用产物(图 4-a, 4-b, 4-c, 4-d; 图 5-a; 图 6-a; 图 7-a; 即不均匀和不规则的致密泥晶与微亮晶组构; Arp et al., 2012), 所以较好地解释了图 6-a、6-b和图 7-a所示的石松藻状组构— — 特别的致密暗色泥晶组构的公共鞘的属性及其可能的形成机理。就像成型丝状葛万菌的暗色致密泥晶壁(图 4-b, 4-c, 4-e; 图 5-b, 5-c; 图 6-c, 6-d; 图 7-b, 7-c, 7-d, 7-e; 图 8-b, 8-c)所代表的可能的活体鞘钙化作用那样, 念珠菌的繁盛可能得益于寒武纪较高的大气圈CO2含量(Berner and Kothavala, 2001), 而且这种念珠菌的公共鞘的钙化作用可能更多得益于寒武纪较高的大气圈O2含量(Saltzman et al., 2011; Large et al., 2019), 还可能得益于寒武系与前寒武系之间的巨型不整合面(Peters and Gaines, 2012)所表征的长时间风化作用所激发的海水中的Ca2+浓度突然从新元古代上升到寒武纪期间的显生宙最大值。
2)暗色致密泥晶中的亮晶显微管(图 6至图 8), 更有可能是蓝细菌细胞列(香毛簇(trichomes); Riding, 1991b, 1991c, 2000, 2011b; Mohr et al., 2011)有机体的降解作用和方解石化作用残余物。就像葛万菌之类的丝状蓝细菌可能的活体鞘的钙化作用形成均匀的暗色泥晶壁所包围并成型了葛万菌丝状体本身的亮晶方解石构成那样, 图 6至图 8所示的致密泥晶组构中的亮晶方解石填充的丝状体, 解释为弯曲的念珠菌丝状体的钙化作用(方解石化作用)产物更加合适, 虽然组成念珠菌的球形细胞的链状丝状体的形态特征没有保存下来。
3)念珠菌的形态学特征和微生物学行为(Potts, 1997; Wright et al., 2005; Soule, 2009; Mohr et al., 2011; Helm and Potts, 2012), 支持将图 6至图 8所示的暗色致密泥晶组构中的亮晶显微管解释为念珠菌菌落的钙化作用残余物。目前, 念珠菌(Nostoc)作为蓝细菌门中在植物命名法下确立的类群Ⅳ 的念珠菌目(Nostocaceae)中的5个属中的最具有代表性的属, 以发育着固氮的异型细胞的许多球形细胞而组成弯弯曲曲的丝状体为特征, 其最突出的特征是分泌含有生物遮光剂伪枝藻素的细胞外多聚糖构成的公共鞘, 从而形成凝胶状菌落并表现出多细胞行为(Shapiro, 1998), 实际上也表现出较为典型的生物模块性(Suosaari et al., 2018)。念珠菌(Nostoc)的命名充满着传奇性, 念珠菌属的描述可以追溯到1600多年前的东晋王朝时代(公元317— 420年), 其在那个时代被名医葛洪作为主要的食物描述为葛仙米(详见李时珍(1596)的《本草纲目》, 参见Potts, 1997), 而且大型的丝状蓝细菌念珠菌在中国还被用来进行头发培育(Mohr et al., 2011); 对念珠菌(Nostoc)的正式命名发生在500年前的欧洲, 该词为15世纪的科学家、哲学家和炼金师Paracelsus所创, 基于那个时代在沙漠中下雨之后膨胀而形成凝胶状可使用物质体所传说的寓言如“ 合唱团的黄油” 以及“ 巫婆的果冻” , 将其提升为“ 哲学家的石头” 而创名, 用来特指“ 一些多血症的和荒唐的形体的污染物, 或者是从一些风湿病状的行星的鼻孔中吹出来的排泄物” ; 但是, 这个奇怪的名称一直被运用到今天, 就像葛万菌在创名的时候当作有孔虫那样, 作为一种能够分泌公共鞘的丝状蓝细菌凝胶状菌落的代表, 念珠菌(Nostoc)分泌公共鞘的基因过程已经取得了一些突破性的认识(Wright et al., 2005; Soule, 2009; Mohr et al., 2011; Helm and Potts, 2012), 并已经认识到其可能起源于大约20× 108年之前(Schopf, 2012)。所以说, 将图 6至图 8所示的暗色致密泥晶组构中的亮晶显微管组构解释为念珠菌菌落的钙化作用残余物, 拓宽了石松藻(Lithocodium; Elliott, 1956; Riding, 1991b, 1991c)或石松藻状组构(Lithocodium-like fabric; Cherchi and Schroeder, 2006; Vé drine et al., 2007; Rameil et al., 2010)的认识, 而且为暗色致密泥晶中的亮晶显微管组构的“ 海绵木乃伊” 解释(Adachi et al., 2009; Kwon et al., 2012; Chen et al., 2014; Li et al., 2015; Park et al., 2015; Coulson and Brand, 2016; Luo and Reitner, 2014, 2015; Hong et al., 2012, 2014, 2016; Lee and Riding, 2018; Shen and Neuweiler, 2018; Lee et al., 2014, 2015, 2016, 2019; Chen et al., 2019)提供了一个重要的研究镜像。
4)念珠菌(Nostoc)的现代类比物确实发生在多重状蓝细菌微生物席之中(De los Rí os et al., 2015), 从另一个侧面支持了将图 6至图 8所示的暗色致密泥晶组构中的亮晶显微管组构解释为念珠菌菌落的钙化作用残余物的推断和解释。生长在极地地区的地表融水水池中的微生物席群落主要由丝状体的颤蓝细菌目所主导, 包括被较细的丝状蓝细菌纤发鞘丝菌(Leptolyngbya)包围的较粗的双须菌(Dichothrix)丝状体, 还包括缠绕的织线菌(Plectonema)和非缠绕的第谷菌(Tychonem), 在其中就发育着数十微米至数百微米的念珠菌(Nostoc)菌落(De los Rí os et al., 2015)。这样的现象与图 6至图 8所示的念珠菌菌落的钙化残余物和丝状葛万菌物质体共生的现象很相似, 即念珠菌菌落生长和发育在由丝状蓝细菌(包括球形蓝细菌)主导的多重状蓝细菌微生物席之中。
5)与海绵组织降解物产生的组构的差异性。基于解剖学和分子数据, 海绵确实是进化最早的动物, 在埃迪卡拉纪早期的沉积物中还发现了海绵骨针(Du and Wang, 2012), 所以地质学家们一直关注着依据海绵及其海绵组织的降解物产生的组构去探索后生动物的起源, 但是得到的研究结果却明显不同于图 6至图 8所示的“ 致密泥晶中的亮晶显微管” 组构。较有代表性的研究包括: (1)多泥组构(polymud fabric), 是指那些含有不规则次生的、而且被间沉积物填充的空洞的自生钙碳酸盐组构, 其是基于现代实例的研究而被Neuweiler等(2007)所定义, 并进一步认为这种结构为硅质海绵正在降解的细胞外凝胶(collagenous)基质(ECM)的钙化作用所产生, 而且类似的组构已经追溯到新元古代(1083— 779 Ma)的小达尔群之中(Neuweiler et al., 2007); 2)一种毫米至厘米级别的、总体上被较薄的泥晶壁所分开的碳酸盐胶结物构成的房室状构造, 被Wallace等(2014)发现于成冰纪微生物礁复合体中, 而且被解释为原始海绵, 包括海绵级别的生物或复杂的微生物前体, 在形态学特征上有点像大型房室状孢网菌状组构; (3)海绵组织的细菌降解作用形成的球粒结构和致密泥晶组构(Reitner, 1993; Reitner and Neuweiler, 1995), 即如果出现很多细菌的话正在腐烂的海绵组织将表现为球粒结构, 而如果出现少量细菌的话海绵组织将崩溃而且形成致密泥晶组构, 但是图 6至图 8所示的 “ 暗色致密泥晶中的亮晶显微管组构” 还包含着亮晶显微管, 而且包含着较多的丝状葛万菌物质体, 所以成为拒绝将这样的组构解释为海绵木乃伊的证据。
因此, 把图 4至图 8之中的暗色泥晶薄膜分隔的微亮晶基质类比于孢网菌(Bacinella; Radoič ić , 1959; Riding, 1991b, 1991c), 并进一步解释为代表着现代蓝细菌微生物席的一些特征的孢网菌状组构(Bacinella-like fabric; Schlagintweit et al., 2013; Schmitt et al., 2019)。把图 6至图 8中的暗色泥晶基质中的亮晶显微管组构类比于石松藻(Lithocodium; Elliott, 1956; Riding, 1991b, 1991c), 并解释为代表着蓝细菌菌落钙化作用残余物的石松藻状组构(Lithocodium-like fabric; Cherchi and Schroeder, 2006; Vé drine et al., 2007; Rameil et al., 2010), 进一步解释为念珠菌菌落的钙化作用残余物。这2种特别的沉积组构明显地发育和生长在由丝状葛万菌表征的蓝细菌微生物席之中, 从微观组构上印证了圣水峪剖面崮山组上部均一石主导的生物层状微生物礁在宏观特征上所指向的光合作用新陈代谢活动的基本特征。
但是, 还是不能将这些构成生物层状微生物礁(图 2-b, 2-c; 图 3-a)的均一石理解为丝状蓝细菌直接的建造物, 因为在高倍显微镜下, 在均一石的致密泥晶组构局部发育的大大小小的丝状蓝细菌物质体之中总是见到稀疏分布的黑色小点(大小多小于1 μ m)(图 4-b, 4-c、4-e; 图 5-b, 5-c; 图 6-c, 6-d; 图 7-b, 7-c, 7-d, 7-e; 图 8-b, 8-c), 其代表着可能的黄铁矿晶体残余物(Berner et al., 1984; Schieber, 2002; Rickard et al., 2017; Siahi et al., 2017; Flannery et al., 2019), 说明形成均一石的微生物沉淀作用(Riding, 2000)和早期石化作用(Dupraz et al., 2009)与硫酸盐还原反应(Rickard et al., 2017; Siahi et al., 2017; Flannery et al., 2019)存在着成因联系, 进一步表明微生物席内碱度发动机(Gallagher et al., 2012)和细胞外聚合物质(Decho, 2010; Tourney and Ngwenya, 2014; Flemming et al., 2016; Decho and Gutierrez, 2017)构成的细胞外有机基质(Helm and Potts, 2012)这2个关键要素的紧密耦合, 并涉及到碳酸盐矿物的原地沉淀作用。由于蓝细菌是最为成功的造席生物(Stal, 2012), 蓝细菌作为原始生产者的微生物活动最终造成了由细胞外聚合物质(EPS)主导的细胞外基质(ECM)的生产作用, 并形成特别的生物膜和微生物席, 而且构成生物膜和微生物席ECM基质在硫酸盐还原细菌之类的异养细菌的降解作用过程中又造成了矿物沉淀作用所产生的钙化作用(Dupraz et al., 2009, 2011), 这一个复杂的作用过程最终形成了寒武系苗岭统非纹层状光合作用微生物席(或生物膜)所主导的均一石, 以及这些均一石主导的生物层状微生物礁。但是, 在苗岭世乃至整个寒武纪较高的大气圈CO2含量以及较高的O2含量之下的方解石海中, 与后生动物的寒武纪大爆发相平行的蓝细菌繁荣、以及不需要借助于CCM机制的蓝细菌鞘的钙化作用, 它们确切的形成机理还有待于今后的深入研究, 因为蓝细菌钙化作用尤其是蓝细菌活体鞘的钙化作用并不是必需的(Riding, 2011b), 而且钙化作用的开启可能要求海水处于很高的碳酸盐饱和状态, 所以说蓝细菌的钙化作用取决于环境条件而且发生得很早, 明显要早于由蓝细菌主导的微生物席所建造的均一石(图 2-b, 2-c; 图 3-a)成型并变成灰岩体的整体钙化作用。但是, 形成均一石主导的生物层状的微生物礁体所要求的复杂的多重钙化作用确切的形成机制和环境响应机理, 明显表现在复杂的地球动力学过程控制的地球生物学过程之中, 所以许多细节问题确实还有待于更加深入全面的研究才能得到更加合理的阐释。
山东省泗水县圣水峪剖面苗岭统崮山组上部均一石主导的生物层状的微生物礁, 构成潮下型碳酸盐岩米级旋回的上部浅水单元, 以一个总体向上变浅的旋回序列组成三级沉积层序的强迫型海退体系域, 与丝状蓝细菌化石(包括葛万菌和可能的念珠菌菌落)和非均一的沉积组构一起, 不仅代表着较为典型的海平面下降阶段的沉积记录, 而且指向了光合作用、新陈代谢作用所主导的沉积作用样式。最为特征的是, 与丝状蓝细菌物质体共生的可以类比于石松藻、并可以进一步解释为石松藻状组构的“ 暗色致密泥晶中的亮晶显微管状组构” , 可以进一步解释和设想为具有公共鞘的丝状蓝细菌菌落即念珠菌菌落的钙化作用残余物, 为这样的组构所表征的许多学者所倡导的“ 海绵木乃伊” 解释提供了另外一个研究镜像, 而且拓宽了对生物亲和性存在巨大争议的石松藻、以及可能代表着特定的钙化蓝细菌菌落的石松藻状组构的进一步了解。将石松藻以及所延伸出的石松藻状组构解释为念珠菌菌落的钙化作用残余物, 为今后对寒武纪蓝细菌繁荣乃至蓝细菌进化研究提供了一个重要的实例。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
| [147] |
|
| [148] |
|
| [149] |
|
| [150] |
|
| [151] |
|
| [152] |
|
| [153] |
|
| [154] |
|
| [155] |
|
| [156] |
|
| [157] |
|
| [158] |
|
| [159] |
|
| [160] |
|
| [161] |
|
| [162] |
|
| [163] |
|
| [164] |
|
| [165] |
|
| [166] |
|
| [167] |
|