


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国晚泥盆世至早三叠世楔叶类植物多样性研究*
[邓珍珍1  , 熊聪慧
, 熊聪慧2 , 黄璞3, 4 , 薛进庄1 ]
, 熊聪慧, 黄璞|
|

第一作者简介 邓珍珍,女,1990年生,2016年毕业于北京大学地球与空间科学学院,从事早期楔叶类分类及多样性演化的相关研究。E-mail: dengzz1990@pku.edu.cn。
化石类群的分类单元多样性和形态多样性变化是生物宏演化的2个基本方面,而以往的研究对于后者的关注较少。晚古生代的楔叶类植物易于识别,化石记录丰富,但针对这一类群多样性演化的研究还较为缺乏。基于华北板块、华南板块晚泥盆世至早三叠世楔叶类植物属、种以及叶片形态编码数据库,对楔叶类植物宏演化历程进行详细研究。中国(华北板块和华南板块)楔叶类植物的属、种丰富度在晚古生代呈现出不稳定的持续增长,表现为晚泥盆世的初始兴盛、早石炭世杜内期至早二叠世萨克马尔期的缓慢上升、早二叠世萨克马尔期至晚二叠世吴家坪期的快速上升;晚二叠世长兴期,总体的属种多样性骤减;每百万年属种多样性在二叠纪—三叠纪之交亦有明显降低。华南板块楔叶类植物的属种多样性总体上小于华北板块,达到峰值的时间为卡匹敦期—吴家坪期,而华北楔叶类植物的属级多样性在空谷期—沃德期达到峰值。中国楔叶类植物叶片的形态多样性的剧烈变化与属种丰富度并不同步,表现为法门期至韦宪期较小,在宾夕法尼亚亚纪达到峰值,随后稍微下降并在二叠纪的大部分时段保持平稳。在晚古生代的叶片形态演化过程中,楔叶类植物的叶面积由小变大、叶片由深裂至不裂、叶尖形状由分裂变为圆形或钝圆、叶轮中的叶从大小相等到大小不等并出现叶镶嵌,这可能与当时植物群落中林下层光照强度的变化有关。
About the first author Deng Zhen-Zhen,born in 1990,graduated from Peking University in 2016. She has been mainly working on the taxonomy and diversity of early sphenophytes. E-mail:dengzz1990@pkuedu.cn.
Taxonomic and morphological diversities of fossil groups are two fundamental aspects in studying macroevolution of life,but previous studies have paid little attention to the latter. Late Paleozoic sphenophytes are easily to be recognized and have a rich fossil record,but their diversity pattern remains little known. Based on a compilation of the genera and species record of the Late Devonian-Early Triassic sphenophytes from South China and North China blocks,and a compilation of leaf morphological characters of some selected leafy species,the macroevolution pattern of early sphenophytes is studied with respect to their taxonomic and morphological diversity. The genus/species richness of the sphenophytes from China shows continuous but uneven increase during the Late Paleozoic,including the initial diversification during the Late Devonian,a slow rise from Visean to Sakmarian,and a rapid rise from Sakmarian to Wuchiapingian;and then,the richness sharply declines at the end of the Permian. The diversity per million years also shows significant decease at the Permian-Triassic transition. Sphenophytes from South China are less diverse at both the generic and specific levels than those from North China,and their diversity reaches the peak during the Capitanian-Wuchiapingian;genus diversity of sphenophytes from North China reaches the peak during the Kungurian-Roadian. Leaf morphological diversity of sphenophytes from China,decoupled from the taxonomic richness,is smaller in the Famennian and early Carboniferous,reaches the peak in the Pennsylvanian,and then slightly declines and remains stable in the Permian. Leaf morphologies of the Late Paleozoic sphenophytes show evolution in the following aspects: increase of leaf area;leaf lamina varying from divided to undivided;leaf tip varying from split to round or obtuse round;same-sized leaves to different-sized leaves in the same leaf whorl,and showing leaf mosaic. These changes might be related to variations of light intensity at the understory levels of the Paleozoic plant communities.
地史时期的生物多样性演变是古生物学研究的重要课题(樊隽轩等, 2016)。化石类群分类单元的多样性变化与形态特征的多样化是生物宏演化的2个基本方面(Benton and Newell, 2014)。分类单元多样性(Taxonomic diversity)以统计不同分类级别如科、属、种的数目变化为主要研究手段, 为研究者所熟知且被广泛应用于各地质时代的不同生物类群; 而形态特征的多样性程度则通常以形态多样性(morphological diversity或morphological disparity, 又称形态歧异度)的定量指标为研究依据, 在方法论方面较为复杂且抽象, 因此相关的研究较少。前人研究发现, 形态多样性常与分类单元多样性呈现出不同步的演化特征(Foote, 1993; Bapst et al., 2012; Xue et al., 2015; Halliday and Goswami, 2016)。
楔叶类植物具节与节间、节上轮生叶片, 最早起源于晚泥盆世, 石炭纪、二叠纪其属种多样性达到最大, 构成全球沼泽森林的优势类群, 具丰富的化石记录(杨关秀等, 1994)。Taylor等(2009)将这一类植物归入楔叶植物门(Sphenophyta), 包括3个目: 羽歧叶目(Pseudoborniales)、楔叶目(Sphenophyllales)和木贼目(Equisetales)。笔者遵从这一分类方案, 以“ 楔叶类植物” 指代楔叶植物门中的所有分子。楔叶类植物的形态特征易于识别, 属种丰富, 为开展化石类群分类单元多样性、形态多样性及二者的对比研究提供了难得的素材, 但目前专门针对这一类群开展的相关研究还较为缺乏。
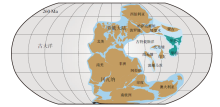
晚古生代, 华南、华北板块及塔里木、柴达木等地块逐渐脱离东冈瓦纳的西北边缘, 并向北半球方向漂移, 石炭纪— 二叠纪位于古特提斯洋的周缘(Huang et al., 2018; 图 1)。中国可靠的楔叶类植物化石记录最早出现于晚泥盆世法门期, 石炭纪和二叠纪, 该类植物达到全盛, 广泛分布于长江流域及华北的很多地区, 三叠纪以后则凋零衰微(杨关秀等, 1994; Li et al., 1995; Wang, 2010; Xue et al., 2015)。现存的楔叶类植物仅有木贼属(Equisetum)约15种, 皆为草本。然而, 在晚古生代, 楔叶类植物中既有高大的乔木, 也有低矮的林下层草本以及攀援的藤本类型(中国古生代植物编写小组, 1974; 杨关秀等, 1994; Taylor et al., 2009; 图 2)。楔叶类的羽歧叶目仅包含1个化石属(Pseudobornia), 发现于北极熊岛与阿拉斯加的晚泥盆世地层中, 被认为是高达20 m的乔木(Schweitzer, 1967)。楔叶目和木贼目首现于晚泥盆世, 在晚古生代广布全球, 其中木贼目又分为芦木科(Calamitaceae)和木贼科(Equisetaceae), 芦木科是具有次生生长的木本植物, 木贼科的化石记录主要出现在中、新生代(Wang et al., 2006; Wang et al., 2006; Taylor et al., 2009; 邓珍珍等, 2016)。
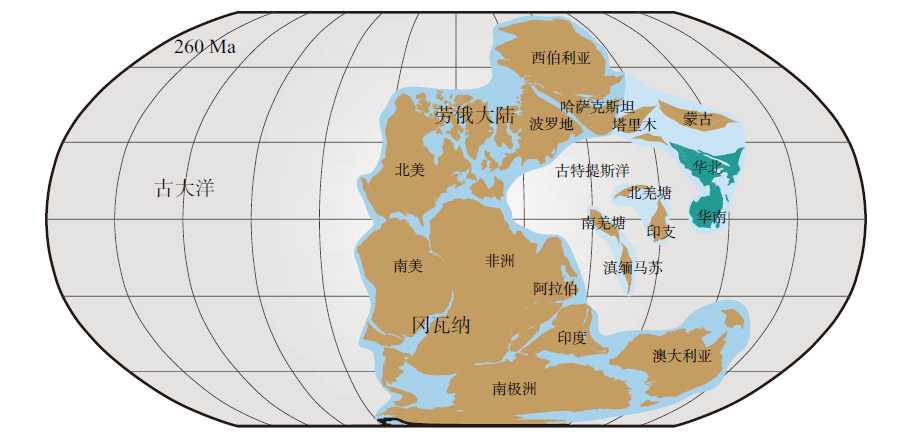 | 图 1 古生代晚期古地理图(约260 Ma), 示华南和华北板块的位置(依据Huang et al., 2018)Fig.1 Late Paleozoic palaeogeographic map(ca. 260 Ma), showing location of South China and North China blocks(based on Huang et al., 2018) |
 | 图 2 晚古生代楔叶类植物景观复原 A— 华南晚泥盆世楔叶目植物 Xihuphyllum景观复原(修改自Huang et al., 2017; 植株高约3 m); B— 华南晚二叠世木贼目植物Calamites景观复原(修改自Feng et al., 2020; 植株高约20 m)Fig.2 Reconstruction of communities of the Late Paleozoic sphenophytes |
楔叶类植物中, 代表营养叶的形态属、种数量最为丰富, 例如楔叶目的楔叶属(Sphenophyllum)、木贼目的轮叶属(Annularia)和瓣轮叶属(Lobatannularia), 这3个属的叶片形态分化是楔叶类植物在晚古生代辐射的主要表现。楔叶类植物为晚古生代陆地植被的重要组成分子, 其多样性演化与华夏植物群的起源、繁盛与衰落息息相关(Li et al., 1995)。对楔叶类植物的属种多样性、叶片形态多样性演化的详细研究有助于深入认识晚古生代的陆地生态系统以及华夏植物群的发展历史。
文中通过统计中国楔叶类植物在晚泥盆世至早三叠世详细的属种记录, 对其多样性变化、灭绝、新生和更替等事件进行深入研究。另外, 以叶片形态学特征为依据, 对晚泥盆世至早三叠世楔叶类植物中的叶片属种进行编码, 采用多元统计分析, 定量研究叶片形态多样性的演化过程。
前人根据构造性质、沉积类型、地层发育历史和古生物特征, 将中国的泥盆系划分为4个地层区: 准噶尔— 兴安地层区、塔里木— 华北地层区、西藏— 滇西地层区与华南地层区(王金星等, 2005a)。石炭系的地层分区与泥盆系相似, 分为4个地层区: 准噶尔— 兴安地层区、华北— 塔里木地层区、华南— 羌塘地层区与喜马拉雅地层区(王金星等, 2005b)。二叠系的地层分区稍有不同, 分为5个地层区: 北部边缘地层区、塔里木地层区、华北地层区、喜马拉雅地层区与华南地层区(谷峰等, 2005; 金玉玕等, 2000)。文中统计的区域为华南地层区与华北地层区(对于泥盆纪和石炭纪, 仅统计华南— 羌塘地层区、华北— 塔里木地层区中的华南和华北区域, 未包含产地为羌塘和塔里木的化石标本)。华南和华北晚古生代植物化石的分布区域, 分别隶属于华南板块与华北板块。其他地层区的化石记录或较为稀少, 或地层时代对比仍有很多不确定性, 因此暂不涉及。因此, 文中所指的中国, 为华南板块与华北板块的总和。
产出楔叶类植物的法门期地层主要有华南板块的五通组、岳麓山组和写经寺组。密西西比亚系主要包括: 华北板块的杨山组、靖远组、臭牛沟组, 华南板块的帽子峰组、测水组和叶家塘组。宾夕法尼亚亚系包括: 华北的红土洼组、本溪组以及太原组下部, 华南的草凉驿组。下二叠统(乌拉尔统)包括: 华北的太原组中上部和山西组。中二叠统(瓜德鲁普统)包括: 华北的下石盒子组与上石盒子组下部, 华南的童子岩组和谷田组等。上二叠统(乐平统)包括: 华北的上石盒子组上部、孙家沟组, 华南的龙潭组与宣威组等。下三叠统包括: 华北的刘家沟组与和尚沟组, 华南的岭文组与九曲江组。各地层单元通过参考原始文献及各地质时期的地层对比方案(吴秀元和赵修祜, 1982; 张善桢等, 1982; Wang, 2010; 全国地层委员会《中国地层表》编委会, 2014), 对应于国际年代地层表中的相应地质时代(见附件1和2)。
楔叶类植物在法门期主要分布于长江中下游的湖南、湖北、江西、安徽、江苏、浙江6省, 另外广东省也有部分楔叶类植物的化石记录。密西西比亚纪楔叶类植物出现在华南地区的广东、江苏、湖南、江西、浙江等省, 华北地区的化石记录较少, 仅在河南、陕西、甘肃有少量产地; 宾夕法尼亚亚纪华南受海侵影响, 植物化石记录稀少, 华北的河南、河北、山西、山东、甘肃、内蒙古等多地广泛分布多种楔叶类植物。早二叠世, 华南楔叶类化石记录极少, 中、晚二叠世楔叶类植物分布于江西、福建、广东等多省; 华北地区在整个二叠纪都有楔叶类植物的广泛分布。
依据《中国地质时期植物群》、《中国古生代植物》、《中国豫西二叠纪华夏植物群— — 禹州植物群》等各类专著及其他文献(如: 中国古生代植物编写小组, 1974; Li et al., 1995; 杨关秀等, 2006; Xiong and Wang, 2011; Xiong et al., 2013; Xue et al., 2015; 2018), 详细统计了华南板块、华北板块晚泥盆世至早三叠世的楔叶目、木贼目植物的属种名录及它们的产地、地层信息等, 包括楔叶目的楔叶属(Sphenophyllum)、木贼目的芦木属(Calamites)、轮叶属(Annularia)、星叶属(Asterophyllites)、瓣轮叶属(Lobatannularia)等(详见附加材料1和2)。
根据植物群的研究程度及区域地层对比的精度, 笔者将中国的晚泥盆世至早三叠世地层划分为11个时段: 法门期(Famennian, t1); 杜内期(Tournaisian, t2); 韦宪期(Visean, t3); 谢尔普霍夫期-莫斯科期(Serpukhovian-Moscovian, t4); 卡西莫夫期-萨克马尔期(Kasimovian-Sakmarian, t5); 亚丁斯克期(Artinskian, t6); 空谷期-沃德期(Kungurian-Roadian, t7); 卡匹敦期-吴家坪期(Capitanian-Wuchiapingian, t8); 长兴期(Changhsingian, t9); 印度期(Induan, t10); 奥伦尼克期(Olenekian, t11)。
根据化石延限与特定研究时段的关系, 可将化石分子划分为4种类型: (1)单延限分子(FL), 其首现层位和末现层位都位于研究层段内; (2)半贯穿分子(bL), 首现层位在研究层段之下, 末现层位位于研究层段内; (3)半贯穿分子(Ft), 首现层位在研究层段内, 末现层位位于研究层段之上; (4)贯穿分子(bt), 首现和末现层位都在研究层段之外。针对这4种类型的分子对多样性的贡献程度, 前人建立了多个生物多样性的计算公式(陈清等, 2012)。对于属级、种级单元的丰富度(richness), 使用以下2种多样性统计公式:
1)总多样性或简单多样性:
Ntot=NFL+NbL+NFt+Nbt
(即: 上述4类分子的总和)
2)无单延限分子多样性:
Ntot-=NbL+NFt+Nbt
总多样性或简单多样性(Ntot)是多样性研究中最常用的统计指标, 但在时间段划分不均一的情况下, Ntot的值容易受到单延限分子数量的影响, 因此, 无单延限分子多样性(Ntot-)可与总多样性曲线进行相互对比。同时, 由于所划分的各时段, 其时间延限长短不一。例如, 二叠纪— 三叠纪之交的几个时段, 其延限较短。因此, 笔者使用了每百万年多样性的指标(多样性除以特定时段的延限), 以降低不同时段的时间延限对多样性的影响。
文中选取的其他多样性指标包括:
3)新生单元数目:
No=NFL+NFt
4)灭绝单元数目:
Ne=NFL+NbL
5)新生比例(新生率):
P=No/Ntot
6)灭绝比例(灭绝率):
Q=Ne/Ntot
7)分类单元(如属、种)更替数目
Nto=No+Ne
8)分类单元(如属、种)净增长数目
Ninc=No-Ne
依据上述指标, 文中分别对中国(华南板块+华北板块)、华南板块、华北板块不同层段的楔叶类植物的分类单元多样性进行了分析。
对楔叶类植物营养叶化石属种进行叶片形态多样性的研究, 包括楔叶目中的楔叶属(Sphenophyllum)、三叉穗属(Eviostachya)、钩蕨属(Hamatophyton)、轮叶蕨属(Rotafolia)和西湖叶属(Xihuphyllum), 木贼目中的星叶属(Asterophyllites)、轮叶属(Annularia)、瓣轮叶属(Lobatannularia)、裂鞘叶属(Schizoneura)和杯叶属(Phyllotheca)等, 共计90个种。这些种属于9个不同的地质时段: 法门期; 杜内期— 韦宪期; 谢尔普霍夫期— 莫斯科期; 卡西莫夫期— 萨克马尔期; 亚丁斯克期; 空谷期— 沃德期; 卡匹敦期— 吴家坪期; 长兴期; 早三叠世(附件3)。与上文中分类单元多样性的划分时段相比, 这里将杜内期、韦宪期合并为一个时段(t2+t3), 将印度期和奥伦尼克期合并为一个时段(t10+t11), 这是由于这4个时期可编码的叶片属种较少。为方便比较, 其他时段仍用t1、t4至t9指代, 与上文保持一致。
参考Xue等(2015)的叶片形态学特征, 文中针对楔叶类植物设定了51个形态学特征(附加材料3), 涵盖整体叶形、叶缘的形态、叶片面积、叶长、叶尖和叶基的形态、叶的分裂与愈合状态、是否存在叶镶嵌与不等叶的现象以及每轮叶的数量。所有性状为二元性状, 即存在该特征赋值为1, 不存在则赋值为0。对90个物种进行编码, 形成多维形态学数据矩阵。
编码后的数据矩阵运用PAST软件(Hammer et al., 2001; http://folk.uio.no/ohammer/past)进行主成分分析(Principal components analysis, PCA)。 之后, 运用主成分轴1、 2、 3来构建二维形态空间, 定性地观察数据点的分布。 不同时段的叶片形态多样性用平均配对不相似度(mean pairwise dissimilarity)来表示(Benson and Druckenmiller, 2014; Xue et al., 2015)。 配对不相似度指的是: 对于任意2个分类单元, 赋值不同的性状数目(即不相似的性状数目)与全部可比较的性状数目之比值。 将某一时段的所有分类单元两两进行比较, 获得配对不相似度, 再将这些数值取均值, 即可得到该时段所有分类单元的平均配对不相似度。 这一指标能够很好地反映不同时段分类单元形态学特征(编码)之间的趋异性, 且受取样强度的影响较小(Benson and Druckenmiller, 2014)。
4.1.1 属种丰富度变化
晚泥盆世法门期至晚二叠世吴家坪期, 中国(华北板块与华南板块)楔叶类植物属种的简单多样性(Ntot)呈现出不稳定的持续增长, 表现在: (1)法门期首次辐射, 而早石炭世杜内期可能由于研究程度不够, 属种多样性不高; (2)早石炭世韦宪期至早二叠世萨克马尔期, 属种多样性缓慢上升; (3)早二叠世萨克马尔期至晚二叠世吴家坪期, 属级多样性趋于平稳, 而种级多样性快速上升, 吴家坪期达到最高峰。晚二叠世长兴期, 多样性急剧下降, 至早三叠世印度期, 降至低谷, 早三叠世奥伦尼克期, 多样性稍有回升。总体上, 无单延限分子多样性(Ntot-)与简单多样性(Ntot)的变化趋势相近, 区别在于, 由于晚二叠世无单延限分子较多, 种级Ntot-在卡匹敦期— 吴家坪期有所降低(表 1; 图 3-A)。
| 表 1 中国晚泥盆世至早三叠世楔叶类植物的总体多样性及每百万年属种多样性数值 Table1 Values of total diversity and genus/species diversity per million of the Late Devonian-Early Triassic sphenophytes from China |
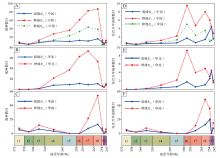
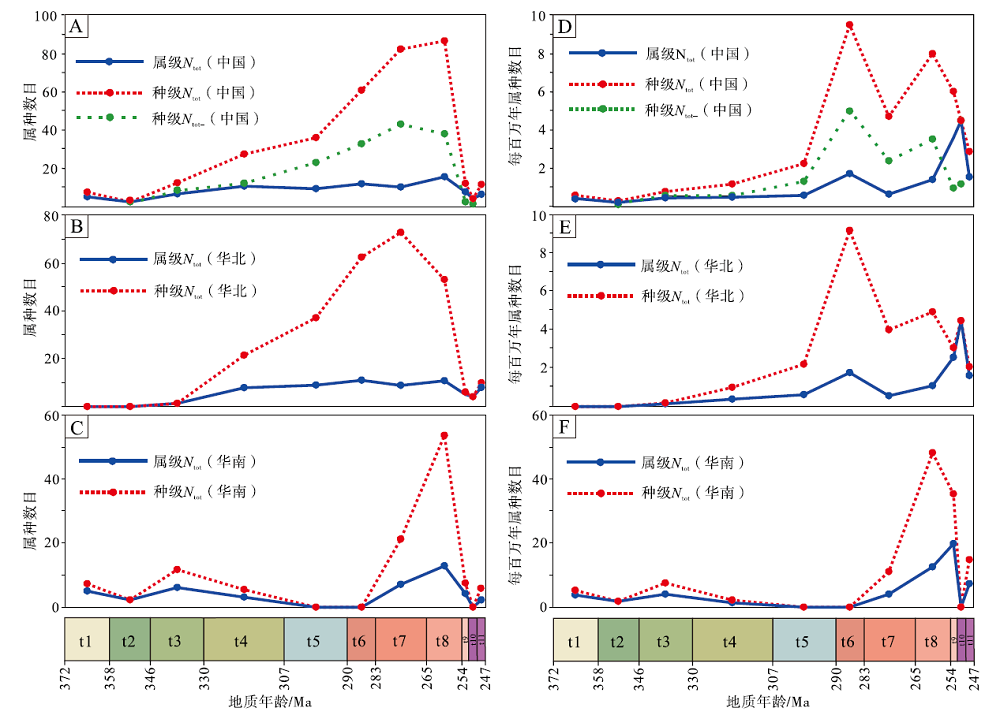 | 图 3 中国晚泥盆世至早三叠世楔叶类植物的总体多样性及每百万年属种多样性变化 A, D— 中国(华南板块+华北板块); B, E— 华北板块; C, F— 华南板块。Ntot— 简单多样性; Ntot-— 无单延限分子多样性; 时段缩写(同图 4, 图 5): t1— 法门期; t2— 杜内期; t3— 韦宪期; t4— 谢尔普霍夫期— 莫斯科期; t5— 卡西莫夫期— 萨克马尔期; t6— 亚丁斯克期; t7— 空谷期— 沃德期; t8— 卡匹敦期— 吴家坪期; t9— 长兴期; t10— 印度期; t11— 奥伦尼克期Fig.3 Total diversity and genus/species diversity per million of the Late Devonian-Early Triassic sphenophytes from China |
华北与华南的属种多样性变化型式有着显著区别: 在华北板块, 楔叶类植物的种级多样性自韦宪期开始持续上升, 至空谷期— 沃德期达到顶峰; 华北的属级多样性峰值出现在卡匹敦期— 吴家坪期(表 1, 图 3-B)。而华南楔叶类植物多样性在法门期和韦宪期出现了辐射; 晚石炭世至早二叠世早期, 华南缺失陆相地层, 因此植物化石记录稀少; 空谷期— 沃德期, 华南楔叶类植物属、种多样性快速上升, 至卡匹敦期— 吴家坪期达到峰值, 长兴期和印度期急剧下降, 奥伦尼克期稍有回升(表 1, 图 3-C)。
每百万年多样性与总多样性变化有着显著区别。中国(华北与华南)楔叶类植物每百万年种级简单多样性和无单延限分子种级多样性变化基本一致, 显示在法门期至萨克马尔期增长极为缓慢, 亚丁斯克期快速增长至峰值, 空谷期— 沃德期有所降低, 卡匹敦期— 吴家坪期再次升高, 随后, 每百万年种级简单多样性在长兴期、印度期和奥伦尼克期持续下降, 但下降幅度不如总体多样性剧烈。每百万年无单延限分子种级多样性则在长兴期下降, 印度期稍微有所回升。楔叶类植物的每百万年属级多样性显示经历了不稳定的持续增长后, 在印度期达到最高值, 奥伦尼克期下降(表 1, 图 3-D)。华北楔叶类植物的每百万年属级多样性变化与中国总体较为一致, 每百万年种级多样性与中国总体的不同在于, 前者显示了印度期有1个回升(表 1, 图 3-E)。华南楔叶类植物每百万年属级、种级多样性变化相似, 均显示了自法门期至吴家坪期的持续增长, 不同之处在于, 属级至长兴期达到最高值, 种级在吴家坪期达到最高值(表 1, 图 3-F)。整体上, 楔叶类植物每百万年多样性在二叠纪— 三叠纪之交的降低不如总体多样性明显, 这是二者的最显著区别。
4.1.2 灭绝、新生和更替
法门期是楔叶类植物的起源时期, 由于这一时期单延限分子的数量较多, 种灭绝率处于较高的程度。杜内期植物基本是法门期植物的延续, 因此新生率为0。韦宪期至早二叠世亚丁斯克期, 中国楔叶类植物的种新生率(0.6~0.9)整体上高于种灭绝率(0.3~0.5), 但谢尔普霍夫期— 莫斯科期是个例外, 推测这一时期较高的灭绝率可能是由于华南缺失陆相地层导致植物化石记录较少造成的。空谷期— 沃德期, 楔叶类植物的新生种比例和灭绝种比例皆为0.5左右, 此后, 新生率持续下降, 至长兴期为低谷(0.17)。与此同时, 则伴随着灭绝率的持续升高, 至长兴期达到高峰(表 2; 图 4-A)。
| 表 2 中国晚泥盆世至早三叠世楔叶类植物新生率、灭绝率和更替率等相关变量数值 Table2 Values of originations, extinctions, turnover numbers and other variables of the Late Devonian-Early Triassic sphenophytes from China |
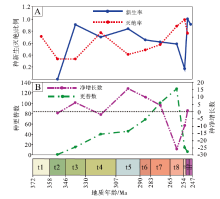
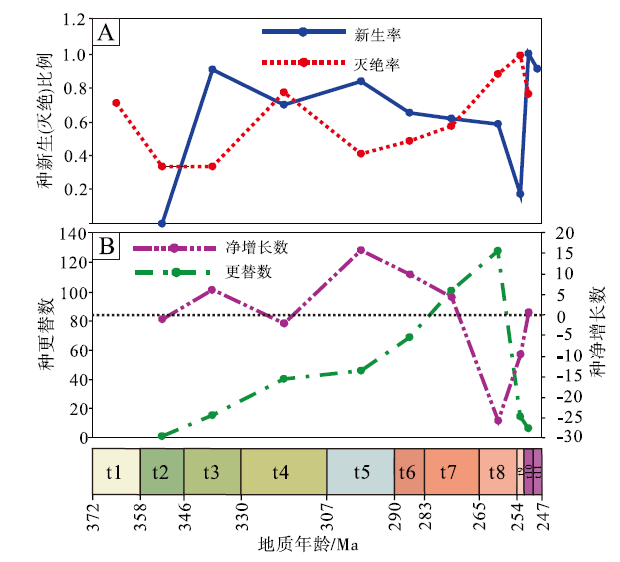 | 图 4 中国晚泥盆世至早三叠世楔叶类植物的种级新生率、灭绝率、更替数及净增长数 A— 种新生率和种灭绝率; B— 种更替数和种净增长数Fig.4 Origination rates, extinction rates, turnover numbers and net growth numbers at species level of the Late Devonian-Early Triassic sphenophytes from China |
中国楔叶类植物的种更替数在法门期和石炭纪缓慢上升, 至二叠纪快速上升辐射, 于卡匹敦期— 吴家坪期达到高峰, 随后急剧下降。物种净增长数目分别在韦宪期和卡西莫夫期— 萨克马尔期出现2次高峰。整体上, 法门期至空谷期— 沃德期, 物种净增长数目为正值, 显示新生物种多于灭绝物种。而自卡匹敦期— 吴家坪期开始, 物种净增长数目为负值, 显示灭绝物种多于新生物种(表 2; 图 4-B)。
4.1.3 楔叶目和木贼目种级多样性变化
中国楔叶类植物最早的多样性辐射是由晚泥盆世楔叶目的起源而贡献的, 随后, 楔叶目的种级多样性持续增加, 杜内期至卡西莫夫期— 萨克马尔期上升较慢, 至亚丁斯克期、空谷期— 沃德期快速上升, 卡匹敦期— 吴家坪期及其后, 呈现下降并趋于灭绝。木贼目始现于韦宪期, 随后物种数目持续增加, 并于空谷期— 沃德期、卡匹敦期— 吴家坪期2个时段快速辐射, 至长兴期下降明显, 早三叠世稍有回升。晚古生代, 木贼目的类群分化和辐射更为快速, 多数时段中木贼目的种级多样性高于楔叶目(表 3; 图 5-A)。
| 表 3 中国晚泥盆世至早三叠世楔叶目、木贼目及代表性属的物种数目 Table3 Species numbers of Sphenophyllales, Equisetales, and some representative genera of the Late Devonian-Early Triassic from China |
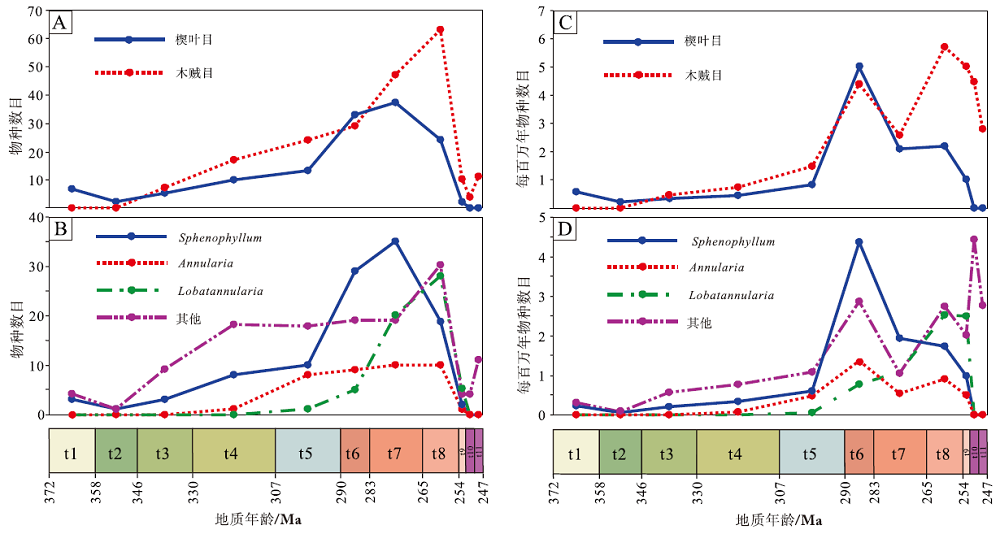 | 图 5 中国晚泥盆世至早三叠世楔叶目、木贼目植物以及常见属的种级多样性 A— 楔叶目和木贼目总体物种数目; B— 代表属的物种数目; C— 楔叶目和木贼目每百万年物种数目; D— 代表属的每百万年物种数目Fig.5 Species diversity dynamics of Sphenophyllales, Equisetales, and some representative genera of the Late Devonian-Early Triassic from China |
楔叶属(Sphenophyllum)是楔叶目中的优势分子, 同时也是晚古生代楔叶类植物中物种最丰富的属, 其种级多样性变化与楔叶目整体较为一致。轮叶属(Annularia)和瓣轮叶属(Lobatannularia)是木贼目中的优势分子。轮叶属的首现较早, 是谢尔普霍夫期— 莫斯科期木贼目中的主要分子, 其物种数目的增长主要发生在卡西莫夫期— 萨克马尔期, 随后直至卡匹敦期— 吴家坪期, 数目基本保持稳定。瓣轮叶属的首现较晚, 在空谷期— 沃德期、卡匹敦期— 吴家坪期这2个时段, 快速辐射, 多样性高于轮叶属(表 3; 图 5-B)。
楔叶目和木贼目每百万年多样性均显示在亚丁斯克期达到高峰, 随后楔叶目分子逐渐减少并灭绝。木贼目在空谷期— 沃德期下降, 卡匹敦期— 吴家坪期又增至最高值, 随后持续下降, 在奥伦尼克期降至最低(表 3, 图 5-C)。楔叶属的每百万年种级多样性变化与楔叶目完全一致, 轮叶属变化与木贼目基本一致, 不同之处在于前者最高值显示在亚丁斯克期。瓣轮叶属每百万年种级多样性显示在卡匹敦期— 吴家坪期和长兴期2个时段最高(表 3, 图 5-D)。整体上, 楔叶目和木贼目及其中几个重要属的每百万年种级多样性变化显示在亚丁斯克期或卡匹敦期— 吴家坪期达到最高值, 在二叠纪— 三叠纪界线附近降低(图 5-D)。
4.2.1 叶片形态空间
将楔叶类植物叶片的多维形态矩阵进行主成分分析, 通过主成分轴的投影, 可定性地观察叶片在形态空间上的分布特征。
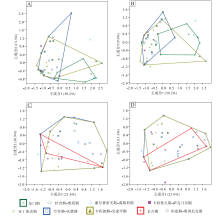
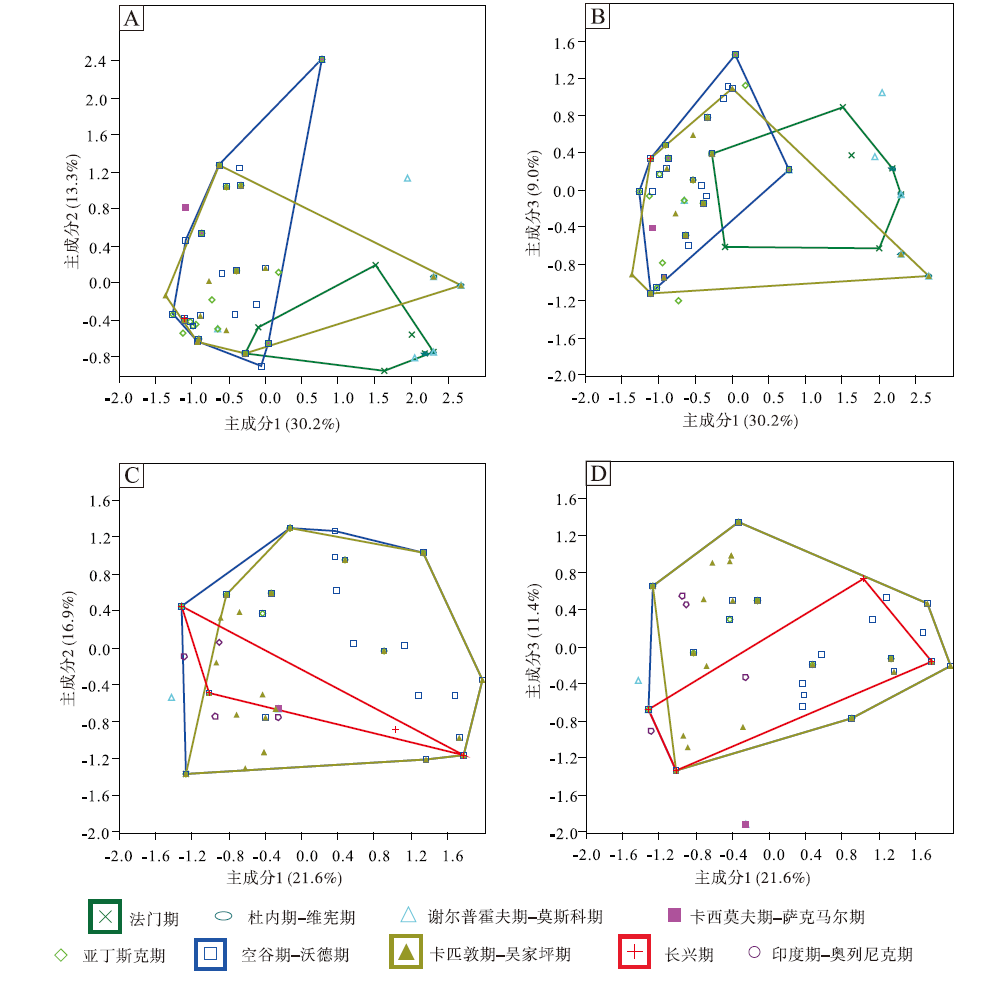
图 6定性地显示了中国楔叶类植物叶片在晚泥盆世法门期至早三叠世奥伦尼克期的形态空间变化。其中, 笔者重点关注了法门期、空谷期— 沃德期、卡匹敦期— 吴家坪期和长兴期共4个叶片形态演化的重要时期。中国楔叶类植物叶片在法门期占据的形态空间最小, 空谷期— 沃德期的形态空间则在石炭纪和早二叠世早期的基础上有所扩大。卡匹敦期— 吴家坪期, 达到最大程度。长兴期, 形态空间大幅度收缩, 至早三叠世收缩到最小程度(图 6-A)。相比全部物种, 中国楔叶类植物首现物种的叶片形态在不同时段的变化更为明显。空谷期— 沃德期, 形态空间较晚泥盆世和早石炭世有了明显扩大, 位置偏移, 同时, 形态空间达到最大。卡匹敦期— 吴家坪期稍有缩小, 位置无明显变化。长兴期, 首现物种的形态空间极小, 仅有几个坐标点。早三叠世, 首现物种的形态空间稍有扩大(图 6-B)。
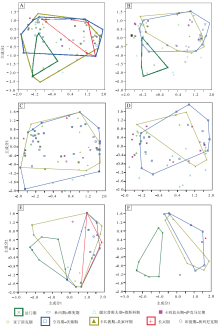
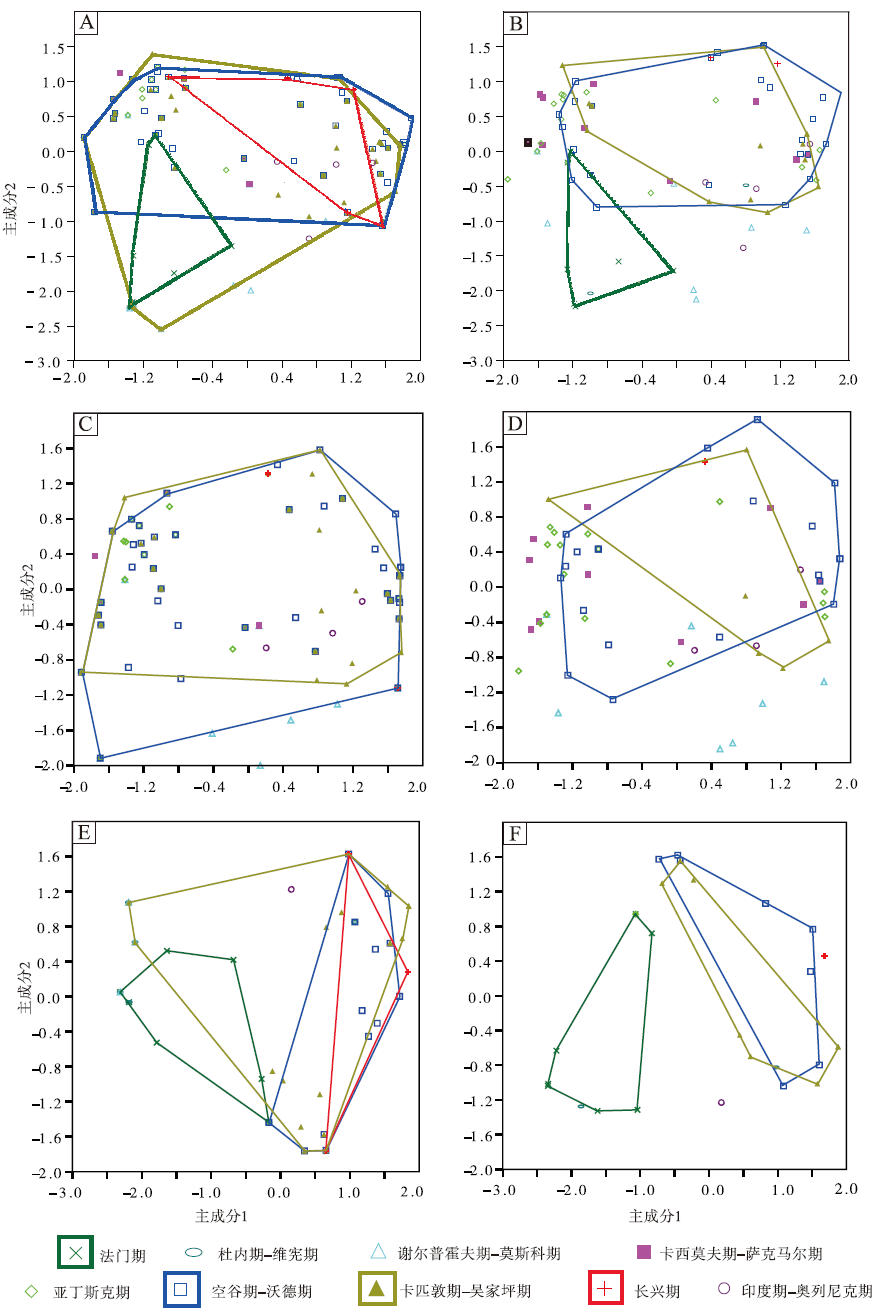 | 图 6 不同时段楔叶类植物叶片的主成分轴1、2的二维形态空间投影(多边形框定了4个时段的物种投点) A— 中国楔叶类的总体物种; B— 中国楔叶类的首现种; C— 华北楔叶类的总体物种; D— 华北楔叶类的首现种; E— 华南楔叶类的总体物种; F— 华南楔叶类的首现种Fig.6 Two-dimensional morphospace plots using PC1-2 axes, showing morphospace occupation of sphenophyte leaves fromdifferent time bins(the convex hulls delimit taxa from four representative time bins) |
华北楔叶类植物叶片的形态空间整体上类似于中国总体的楔叶类植物。不同的是, 华北楔叶类植物在晚泥盆世至早石炭世缺乏记录, 在空谷期— 沃德期达到最大的占据面积, 早于中国总体楔叶类植物(图6-C)。首现物种的形态空间在不同时间段的差别稍大(图 6-D)。华南尽管在晚石炭世至早二叠世早期缺失陆相地层, 但楔叶类植物的叶片形态空间变化仍较为连续。空谷期— 沃德期, 楔叶类植物叶片的形态空间在晚泥盆世至早石炭世的基础上扩大, 但位置发生明显偏移。卡匹敦期— 吴家坪期, 形态空间达到最大。长兴期, 华南的楔叶类植物仍占据小部分的形态空间, 但位置与早期的楔叶类植物不重合。早三叠世, 形态空间极小, 仅为少数几个坐标点(图 6-E)。华南首现物种的形态空间比较突出的特征为: 二叠纪的形态空间与晚泥盆世、早石炭世均不重合(图 6-F)。
4.2.2 形态多样性与物种丰富度变化
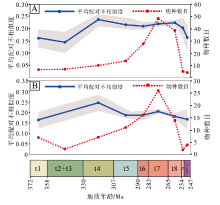
中国楔叶类植物叶片的形态多样性(用平均配对不相似度表示)与编码的物种丰富度的变化并不同步。物种丰富度在晚泥盆世至晚二叠世早期呈现出早期缓慢增长、后期快速辐射模式, 而形态多样性在法门期、杜内期— 韦宪期较小, 谢尔普霍夫期— 莫斯科期达到峰值, 随后稍微下降并保持平稳, 在二叠纪的大部分时间内, 变化不大(表 4; 图 7-A)。首现物种的形态多样性峰值仍在谢尔普霍夫期— 莫斯科期, 二叠纪稍有下降并趋于平稳(表 4; 图 7-B)。晚二叠世长兴期至早三叠世, 形态多样性的降幅不如物种丰富度明显(图 7-A, 7-B), 表明尽管物种发生了大幅灭绝, 但残存物种在形态特征上仍体现了很大的差异性。
| 表 4 中国晚泥盆世至早三叠世楔叶类植物叶片的全部编码物种、首现种的物种数目及其平均配对不相似度 Table4 Numbers of the coded species and first-appeared species, and their mean pairwise dissimilarity among the Late Devonian-Early Triassic sphenophytes from China |
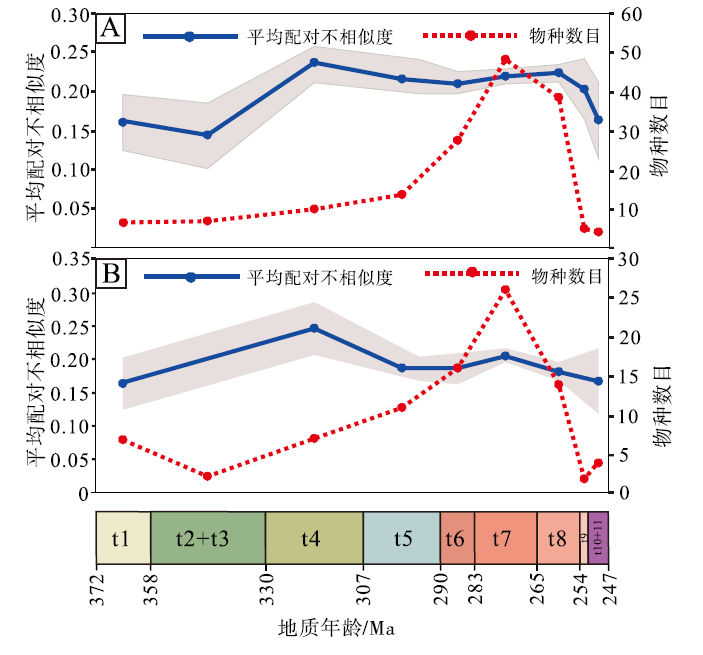 | 图 7 中国晚泥盆世至早三叠世楔叶类植物的物种数目与平均配对不相似度的对比 A— 总体物种; B— 首现种; 时段缩写同图 3: t2、t3可编码的物种数目较少, 因此合并; t10、t11同理Fig.7 Comparison of species richness and mean pairwise dissimilarity of the Late Devonian-Early Triassic sphenophytes from China |
4.2.3 楔叶目和木贼目的形态空间
通过主成分分析, 楔叶目和木贼目在主成分轴1上可清楚区分(图 8-A, 8-B)。参考不同变量对主成分的荷载, 认为主成分轴1主要受每轮叶的数量、叶尖角度、叶连合以及叶形等性状的影响。由楔叶目至木贼目(主成分轴1), 每轮叶的数量逐渐增多, 叶尖角度由钝角变为锐角, 叶形由楔形变为倒披针形, 叶片逐渐连合。不同时段的楔叶目叶片在形态空间上的范围较为重合, 反映它们的特征变化较为连续。楔叶目植物的叶形变化主要表现在主成分轴2和轴3上。参考不同变量对主成分轴2的荷载可以发现, 楔叶属(Sphenophyllum)的叶形态具有以下演化模式: 叶尖形状由分裂变为圆形或钝圆, 叶面积由小变大, 由基部叶裂到不裂, 出现不等叶和叶镶嵌的现象(图 8-A, 8-B)。瓣轮叶属(Lobatannularia)和轮叶属(Annularia)的大部分物种在主成分轴2和轴3上能够清楚区分; 由轮叶属至瓣轮叶属, 不等叶和叶连合更为明显, 叶面积更大, 叶尖趋于圆形或钝圆(图 8-A, 8-B)。
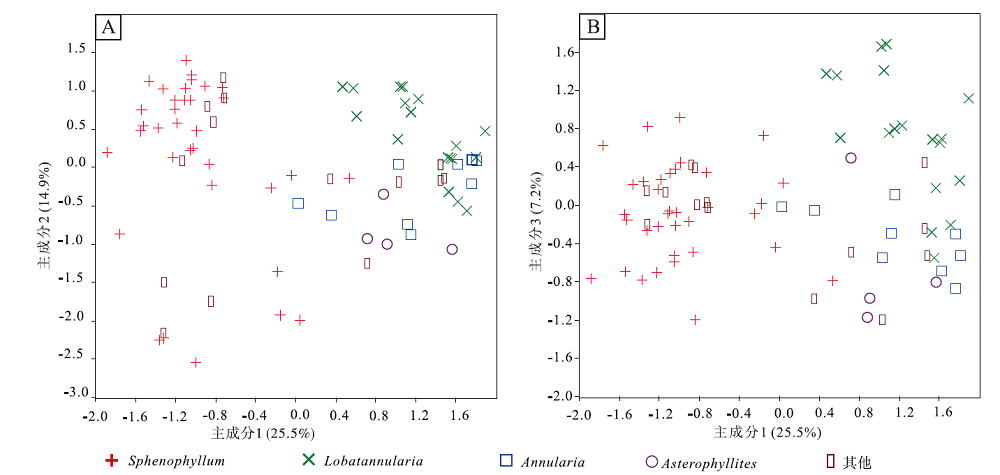 | 图 8 中国晚泥盆世至早三叠世楔叶类植物叶片的主成分轴投影 A— 主成分轴1和主成分轴2; B— 主成分轴1和主成分轴3。主成分分析依据90个种、51个形态学特征。主成分轴1、主成分轴2、主成分轴3为51个形态学特征的不同的线性组合, 反映植物叶片整体形态的变化。叶片整体形态的变异程度(方差)在主成分轴1上最大, 在主成分轴2、主成分轴3上依次递减Fig.8 Morphospace plots using PC axes, of the Late Devonian-Early Triassic sphenophytes from China |
中国楔叶目植物叶片形态空间的投影显示, 楔叶目在晚泥盆世法门期占据了一定的空间, 谢尔普霍夫期— 莫斯科期达到最大, 随后缩小, 晚二叠世长兴期和早三叠世, 楔叶目已基本灭绝(图 9-A, 9-B)。木贼目植物叶片的形态空间投影显示, 该目在谢尔普霍夫期— 莫斯科期开始占据一定的形态空间, 随后扩大, 空谷期— 沃德期达到最大, 卡匹敦期— 吴家坪期稍有缩小, 长兴期和早三叠世, 残存的木贼目分子仍占据一定的形态空间范围(图 9-C, 9-D)。
 | 图 9 中国晚泥盆世至早三叠世楔叶目、木贼目植物叶片的主成分分析(多边形框定了不同时段的物种投点) A, B— 楔叶目植物叶片的主成分分析及二维投影; C, D— 木贼目植物的主成分分析及二维投影Fig.9 PCA analysis of leafy species of the Late Devonian-Early Triassic Sphenophyllales(A and B)and Equisetales(C and D) from China(the convex hulls delimit taxa from different time bins) |
王自强(1989)研究表明, 华北晚古生代至三叠纪初维管植物总体多样性的主要特征是: 多样性持续增长, 在早二叠世亚丁斯克期达到最高, 空谷期至吴家坪期稍有下降, 长兴期明显下降。文中研究显示, 华北的楔叶类植物在该时期的多样性曲线与维管植物的总体多样性特征稍有不同, 楔叶类植物在空谷期— 沃德期达到最高峰, 稍晚于维管植物。但华北及中国总体楔叶类植物每百万年种级多样性均在亚丁斯克期达到最高值, 同时, 在长兴期及附近下降幅度不如总体多样性剧烈。Wang等(2010)认为早泥盆世到早石炭世是华南陆生维管植物分化和分异的阶段, 多样性起伏较大, 植物类型更替快, 从二叠纪早期开始植物多样性迅速发展, 并在中、晚二叠世达到最高峰, 二叠纪末陆生维管植物受到生物大灭绝事件的影响多样性极度下降并发生根本的更替。Xiong等(2013)研究了志留纪至石炭纪华南陆生维管植物的多样性, 在法门期至巴什基尔期多样性有2次起伏, 峰值分别出现在法门期和韦宪期。Xiong 和Wang(2011)的研究结果显示华南二叠纪至三叠纪的陆生植物, 其多样性峰值出现在吴家坪期, 并且长兴期显著下降。文中研究表明, 楔叶类植物在华南地区的多样性变化与华南维管植物群的整体多样性是同步的。
华北的楔叶类植物化石记录相对较晚, 但发展很快, 辐射期早于华南。而华南的辐射期较晚, 其衰退也晚于华北。这一现象可能是由古地理、古气候以及植物群演化所导致的。晚泥盆世至早石炭世(约380— 340 Ma), 华南板块位于古赤道位置, 处于湿热的气候带, 华北板块则位于赤道以北, 并逐渐向北漂移(Huang et al., 2018)。宾夕法尼亚亚纪及二叠纪之初(约320— 290 Ma), 华北板块渐渐向赤道靠近, 或近于赤道, 并多为海陆交互相沉积, 华南几乎全部为海相沉积, 所以华南的植物化石记录稀少, 而华北在热带— 亚热带的燥热潮湿环境的滨海沼泽地带丛生茂密的树林(Li et al., 1995)。这使得华南植物群发育的进程受到了一定的影响, 从而导致华南楔叶类植物辐射的相对迟缓。中二叠世至早三叠世(约270— 250 Ma), 由于华北板块向北漂移逐渐离开赤道湿润带, 干旱的环境来临, 而华南地区的纬度较低, 在吴家坪期尚保持着湿润多雨的生态环境(Li et al., 1995), 这使得华南楔叶类植物的衰退晚于华北。
晚泥盆世法门期的楔叶类植物均为楔叶目, 其中赫氏三叉穗(Eviostachya hoegii)、轮生钩蕨(Hamatophyton verticillatum)、假弱楔叶(Sphenophyllum pseudotenerrimum)、松滋轮叶蕨(Rotafolia songziensis)的叶片分裂程度非常高, 裂片呈线形, 前人认为这种叶形代表了楔叶类植物比较原始的类型(杨关秀等, 1994)。但该时期也存在龙潭楔叶(Sphenophyllum lungtanense)和大型西湖叶(Xihuphyllum megalofolium)具有片化叶子且面积较大的分子, 表现出了较为进化的性状。石炭纪叶片形态空间扩展的原因有两点, 一是更多楔叶属物种的出现, 如弱楔叶(Sphenophyllum tenerrimum)、椭圆楔叶(Sphenophyllum oblongifolium)和微缺楔叶(Sphenophyllum emarginatum)等; 二是芦木科的星叶属与轮叶属分别首现于密西西比亚纪和宾夕法尼亚亚纪。石炭纪的长叶星叶(Asterophyllites longifolius)、细星叶(Asterophyllites tenuifolius)、假星轮叶(Annularia pseudostellata)的共同特点是叶为细线形, 叶长远大于叶宽, 叶面积小。
早二叠世, 叶裂程度高的类型消失, 新出现的以畸楔叶(Sphenophyllum thonii)为代表的楔叶目成员叶较大, 最大者长5.5 cm, 叶大小相近, 两侧不对称, 也出现了叶三对型排列的中朝楔叶(Sphenophyllum sino-coreanum)、襄垣楔叶(Sphenophyllum xiangyuanense)等。该时期出现的纤细轮叶(Annularia gracilescens)、尖头轮叶(Annularia mucronata)等在叶形上与石炭纪线形叶的芦木科分子相比, 长宽比变小, 叶面积增大, 叶为披针形和倒披针形, 并且出现了同一叶轮中的叶大小不等的情况。华夏植物群的代表类群瓣轮叶属(Lobatannularia)出现在这一时期, 该类植物的特点是叶轮形成明显的两瓣, 叶缺明显, 每一瓣中的叶大小不等, 如舌瓣轮叶(Lobatannularia lingulata)、剑瓣轮叶(Lobatannularia ensifolia)等。
中二叠世至晚二叠世早期出现的楔叶目成员大多为三对型分子, 如不等楔叶(Sphenophyllum imparium)、薄楔叶(Sphenophyllum tenuie)等。新出现的轮叶属分子高平轮叶(Annularia gaopingensis)则具有叶缺。中二叠世也新出现多个瓣轮叶属分子。晚二叠世由于有不少中二叠世的物种延续, 总的形态空间收缩不明显。但没有新的楔叶目成员产生, 新出现的物种大部分属于瓣轮叶属, 使得首现物种的形态空间非常小。
楔叶目和木贼目植物叶片在晚古生代展现了相似的演化模式, 即叶面积由小变大, 叶轮中的叶由大小相等到大小不等, 由均匀排布到出现上下叶缺, 叶轮的生长角度也发生改变, 出现叶镶嵌的现象。这些改变可能反映了植物接收到的光照强度的变化。根据植物与光照强度的关系, 植物被分为阳性植物、阴性植物和耐阴植物三大生态类型, 而阴性植物的叶柄常或长或短, 叶形叶或大或小, 使叶子成镶嵌状排列在同一平面上以充分利用阴暗不足的光线(曲仲湘等, 1984)。中二叠世至晚二叠世早期, 很多楔叶目和木贼目新分子出现了符合阴性植物特征的性状。作为林下层的楔叶类植物, 这种形态上的改变, 可能暗示它们所能接收到的光照强度的减弱, 可能指示当时的森林变得更为郁闭, 因此, 增大叶片面积和出现叶镶嵌等更有利于它们获取更多的阳光。
1)晚泥盆世至早三叠世, 中国楔叶类植物属种多样性经历了法门期的初始辐射、杜内期至萨克马尔期的缓慢上升、萨克马尔期至吴家坪期的快速上升、以及长兴期的急剧下降和其他的低谷期。每百万年属种多样性在二叠纪— 三叠纪之交亦有明显降低。华北楔叶类植物种级多样性在空谷期— 沃德期达到峰值, 而华南楔叶类植物的种级多样性峰值出现在卡匹敦期— 吴家坪期。楔叶类植物在长兴期的灭绝幅度最大。楔叶目的属种多样性在早二叠世早期上升较快, 晚二叠世呈现下降趋势。木贼目的类群分化和辐射更为快速, 多数时段的种级多样性高于楔叶目。
2)与属种丰富度的剧烈变化模式不同, 中国楔叶类植物叶片的形态多样性在演化初期时较小, 宾夕法尼亚亚纪达到峰值, 随后稍微下降并保持平稳, 在二叠纪的大部分时段内变化不大。晚古生代, 楔叶目和木贼目植物叶片展现了相似的演化模式, 即叶面积由小变大, 叶轮中的叶由大小相等到大小不等, 由均匀排布到出现上下叶缺, 叶轮的生长角度也发生改变, 出现叶镶嵌的现象。林下层可接收光照强度的改变可能是这些改变的原因之一。
该文共包含3个附加材料, 均为excel格式。附件材料下载见中国知网或扫描文章OSID码。
附加材料1:晚古生代楔叶属种分类多样性名录-华北
附加材料2:晚古生代楔叶属种分类多样性名录-华南
附加材料3:中国晚古生代楔叶形态编码
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|