

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
鱼粪化石特征对早侏罗世托阿尔期湖泊生态系统的启示:以川东大安寨段为例*
[周小梅1  , 金鑫
, 金鑫1, 2 , 吴强旺1 , 葛禹1 , 时志强1, 2 ]
, 金鑫]
|
|

第一作者简介 周小梅,女,1994年生,硕士研究生,成都理工大学沉积地质研究院古生物学与地层学专业。E-mail: 276378283@qq.com。
早侏罗世托阿尔期早期大洋缺氧事件(Toarcian Oceanic Anoxic Event,~183.8 Ma)是一次全球性的多幕式生物—环境事件,其在四川盆地下侏罗统自流井组大安寨段亦有显示。对采集自川东北地区大安寨段的鱼粪化石进行形态学描述和内含物分析,发现鱼粪化石中含有大量的磷质成分,多为未经消化的鱼骨化石,另有大量介壳类化石与有机质混杂在一起。据露头岩性和显微薄片特征认为: (1)粪化石呈螺旋状,生产该粪化石的鱼类为大型肉食性鱼类,其喜食小型鱼类和软体动物等,推测可能是肺鱼类(角齿鱼);(2)鱼粪化石形成于开放型淡水湖泊中的半深湖—深湖区,其中湖泊表层含氧量丰富,主要由生产者、消费者构成了研究区早侏罗世托阿尔期大型湖泊生态系统,食物链较为复杂,而湖底为水动力条件较弱的还原环境。该研究成果可为早侏罗世托阿尔期大洋缺氧事件在湖泊中的沉积响应及生态影响研究提供依据。
About the first author Zhou Xiao-Mei,born in 1994,is a master degree candidate of paleontology and stratigraphy in Institute of Sedimentary Geology,Chengdu University of Technology. E-mail: 276378283@qq.com.
The Toarcian oceanic anoxic event(T-OAE,Early Jurassic)was an episode of profound environmental changes marked by a global carbon-cycle perturbation,marine mass extinctions,and black-shale deposits. Morphological description and inclusion analysis of fish coprolites collected from the Da’anzhai Member of the Lower Jurassic Ziliujing Formation in the northeast Sichuan Basin were carried out in this paper. We found that the interior of coprolites is composed of abundant organic matters,silts,incomplete clay minerals,phosphorous fossils,undigested fish bone,and a large number of ostracods. According to the morphology and microscopic examination of the coprolites,we speculate that they originated from lungfish. The fish coprolites were deposited in an intermediate to deep lake. During the Da’anzhai depositional period,the surface water of the Sichuan lake was enriched in oxygen,and the lake ecological system was healthy. The bottom of the lake was characterized by deposits of black shale,which implies an anoxic and reducing environment. The T-OAE fish coprolites may suggest a stratified water column in a Toarcian lake. The above results may provide a basis for the study of sedimentary response and ecological impact in the lake of Toarcian oceanic anoxic events of early Jurassic.
粪化石是古生物和古生态信息的重要来源之一, 在过去几十年中, 粪化石的研究取得了重大进展(Brachaniec et al., 2015)。粪化石不仅可以提供关于饮食、消化道结构、生物群多样性的信息, 而且能间接反映动植物生活的环境, 因此可以作为研究深层生态系统功能和结构的重要工具(Nakajima and Izumi, 2014), 是解释生态群落中古代动物的饮食习惯和营养水平的重要信息来源(Khosla et al., 2015)。此外, 粪化石作为重建动物饮食和行为的重要古生物学来源, 直接记录了远古动物之间的捕食— 被捕食关系, 是重建过去生态系统营养结构和能量流模型的优良材料(Eriksson et al., 2011)。从描述的角度可以将粪化石定义为具有浑圆状外形、石化的动物消化物、排泄物和排出物的总称(龚一鸣等, 2009)。在自然界中粪化石较为罕见, 在保存地或多或少地发育, 成分不确定, 但都保存着粪便的原始形态。不同种类的脊椎动物产生的粪便通常具有不同的外形(Luo et al., 2017), 如爬行类动物的粪便有的像松球, 有的像一段香肠, 而鱼肠内的螺旋瓣由于具复杂的结构, 故所产生的粪便是螺旋型, 特别是所有的盾壳鱼、软骨鱼等(McAllister, 1987)。
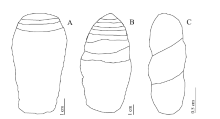
螺旋型粪化石一般包括3种亚型(图 1): 典型异极、异极边缘和双极(Dentzien-Dias et al., 2012)。通常鲨鱼粪便呈异极螺旋形态, 而鱼类粪便呈双极螺旋形态(Diedrich and Felker, 2012)。
 | 图 1 螺旋型粪化石类型(据Dentzien-Dias et al., 2012) A— 异极边缘, 螺旋纹分布于粪化石的末端; B— 典型异极, 螺旋纹集中于粪化石的一端, 间隔紧密, 不同类型的粪化石螺旋纹数量不等; C— 双极, 螺旋纹均匀的布于整个化石, 数量少, 间隔宽阔(Neumayer, 1904)Fig.1 Types of helical coprolites (after Dentzien-Dias et al., 2012) |
保存粪化石的基本条件为: 基本不被搬运, 快速原地埋藏, 埋藏环境贫氧、准厌氧或厌氧, 低水动能条件(对水下环境而言)、水温不能太高、水体的酸碱度适中(王大锐和韩兆宽, 1999)。根据这些条件要求, 亦可反演古代鱼类生活时的生态环境。同时, 粪化石中含有丰富的有机质, 主要成分是磷酸钙, 对其显微组分分析可以了解排泄者的食性(王大锐和韩兆宽, 1999)。
在中国, 粪化石、尤其是鱼粪化石研究较少, 已发现的粪化石产出的年代主要为中生代, 如鄂尔多斯盆地三叠系延长组(刘宪亭, 1953; 李德生等, 1982)、云南禄丰上三叠统(边兆祥等, 1991)、贵州桐梓中侏罗统沙溪庙组(尹赞勋, 1945)。二叠纪末生物大灭绝捕食驱动的中生代海洋革命开始以来, 越来越多的来自海洋地层的粪化石被报道, 显示了粪化石研究在探索海洋生态系统化石食物链中的重要意义(Brachaniec et al., 2015; Luo et al., 2017)。然而却鲜有淡水环境中的粪化石被发现, 因此缺少研究粪化石在淡水湖泊生态系统食物链的化石材料。本次研究描述了在川东北达州地区大槽剖面下侏罗统自流井组大安寨段黑色泥质岩中发现的鱼类粪化石。在四川盆地, 早侏罗世托阿尔期大洋缺氧事件在湖泊沉积中的反映曾被报道(Xu et al., 2017), 本次研究采集的鱼粪化石就产出自响应托阿尔期大洋缺氧事件的湖泊沉积物中。通过对该鱼粪化石研究, 有助于明确湖泊中的捕食— 被捕食关系和重建古生态系统, 为侏罗纪托阿尔期早期大洋缺氧事件在湖泊中的沉积响应及生态影响研究提供依据。
早侏罗世以来, 四川盆地构造活动相对较弱, 古大湖发育, 湖泊内主要沉积砂岩、泥岩及碳酸盐岩(韩永辉和郭正吾, 1984)。在川东北地区, 下侏罗统自流井组发育碎屑岩及湖泊碳酸盐岩(丁一等, 2013), 下伏地层为三叠系须家河组, 二者呈平行不整合接触; 上覆地层为中侏罗统新田沟组, 二者系整合接触(梁西文等, 2014)。自流井组从下至上可分为 4个岩性段: (1)珍珠冲段, 以紫红色泥岩为主; (2)东岳庙段, 灰色、黄灰色粉砂岩、微粒石英砂岩和介壳灰岩, 夹紫红色泥岩; (3)马鞍山段, 以紫红色泥岩为主; (4)大安寨段, 总厚70~100 m, 灰色介壳灰岩与灰黑色、紫红色泥岩互层(张葳等, 2013)。
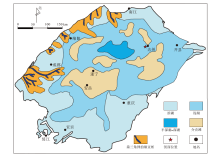
大安寨段沉积可划分为半深湖— 深湖、浅湖、滨湖、介壳滩及扇三角洲前缘亚相5种沉积(图 2) (陈超等, 2020), 主要以滨浅湖泥、浅— 半深湖泥、介壳滩沉积为主, 整体岩性粒度较细, 沉积水体深度较大(张晓鹏, 2005)。大安寨段沉积时期四川盆地处于构造稳定阶段, 但有微弱的湖盆振荡导致湖水的收缩与扩展, 使介壳灰岩、页岩频繁间互或侧向接触(邓燕等, 2017)。该时期物源供应不足, 气候温暖、潮湿(王全伟, 2006), 生物繁多, 以淡水双壳类、介形虫、腹足瓣鳃类为主, 介壳滩呈环状(图 2)分布于水体清澈、能量较强的浅湖区域(张晓鹏, 2005; 黄开伟等, 2010)。根据碳、氧同位素相关性分析(张葳等, 2013; 邓燕等, 2017), 川中地区大安寨段沉积时期的湖泊类型为开放型淡水湖泊。
 | 图 2 四川盆地下侏罗统自流井组大安寨段沉积相(修改自陈超等, 2020)Fig.2 Sedimentary facies distribution of the Da’ anzhai Member of Lower Jurassic Ziliujing Formation in Sichuan Basin(modified from Chen et al., 2020) |
对早侏罗世托阿尔期大洋缺氧事件的研究已有近60年的历程。针对海相地层的研究表明, 该事件发生时期海水温度升高, 海平面上升, 下层水体缺氧、表层水体富营养化, 海相黑色页岩广泛发育, 有机碳普遍富集, 碳、氧、锶、锇和氮同位素、镁/钙值等出现异常, 海洋底栖生物集群灭绝(Jenkys, 1985, 1988)。该事件对中国陆地生态系统也产生了影响(邓胜徽等, 2012), 并在四川盆地及鄂尔多斯盆地陆相沉积中相继被识别出来(Xu et al., 2017; Jin et al., 2020)。托阿尔期大洋缺氧事件发生时陆地气温升高, 在内陆出现了干旱化趋势, 聚煤作用明显减弱或中断, 出现湖相黑色页岩及红色沉积, 植物群出现明显衰退, 喜热型(南方型)植物属种比例增加, 并向高纬度迁移(邓胜徽等, 2012)。
研究剖面位于四川省东北部达州市达川区大槽村附近公路边(图 2), 剖面起点为东经107° 22'9″, 北纬31° 8'31″。大槽剖面大安寨段岩性主要为介壳灰岩与泥页岩, 粪化石产于大安寨段中、上部黑色、灰黑色泥页岩、粉砂质泥页岩层内(图 3)。粪化石的寄主地层发育水平层理, 富含有机质, 反映沉积环境为低能还原环境, 且沉积速率缓慢。粪化石围岩中含有丰富的生物碎屑残骸(植物叶片、生物介壳)和大量个体较大的双壳类实体化石。
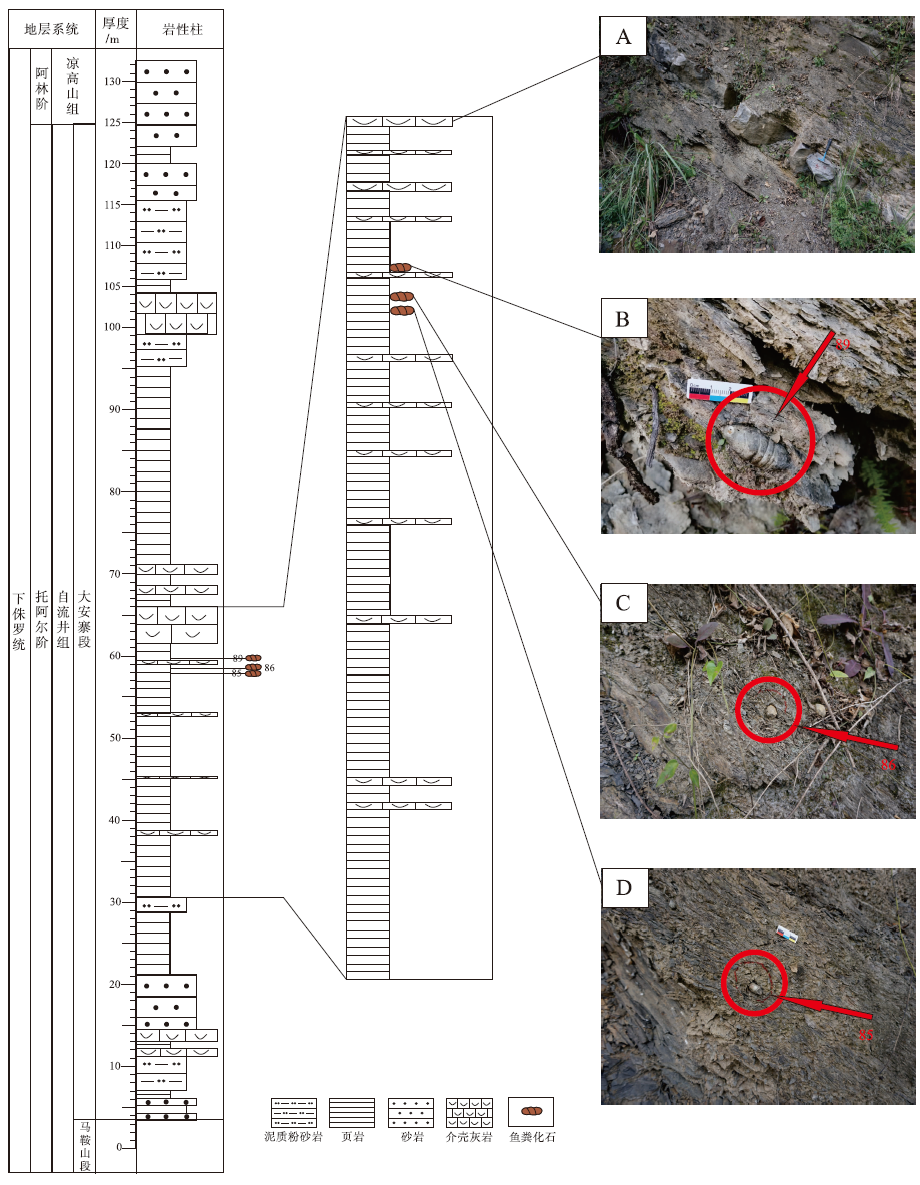 | 图 3 川东北大槽剖面下侏罗统自流井组大安寨段地层柱状图及野外照片Fig.3 Lithological column and field scopes associated with coprolites of the Da’ anzhai Member of Lower Jurassic Ziliujing Formation in Dacao section of northeast Sichuan Basin |
本次共采集到3枚较为完整的鱼粪化石(野外标注样品号为85、86和89)。85、86号样品为保存较好的厘米级大小的粪化石(图 4-A, 4-B), 个体较大, 形态不完整, 推测为沉积时已碎裂的鱼粪形成的粪化石, 其中85号化石呈松球状或圆球状, 而非螺旋状, 本次研究难以准确描述其完整外形(图 4-A, 4-B)。89号样品为1枚保存完好的典型异极螺旋状粪化石, 可见其完整外形(图 4-C, 4-E, 4-F)。
 | 图 4 川东北大槽剖面下侏罗统自流井组大安寨段鱼粪化石宏观照片 A— 85号粪化石; B— 86号粪化石; C— 89号粪化石正面; D— 89号粪化石背面; E— 89号粪化石侧面1; F— 89号粪化石正面2Fig.4 Morphology of fish coprolites of the Da’ anzhai Member of Lower Jurassic Ziliujing Formation in Dacao section of northeast Sichuan Basin |
粪化石(89号样品)呈浅灰色, 表面比较平滑, 整体呈纺锤状, 长51.50 mm、宽20.00 mm、厚17.10 mm。当鱼粪被迅速埋藏而得以保存下来时, 多少会因为受压而变扁, 本枚化石受压较小, 只有轻微变扁现象。该鱼粪化石上有5个明显的旋纹(整体似腹足类的内型), 螺旋纹主要集中在一端且具有螺旋纹的一端稍粗大, 全部螺旋约占全长1/2, 旋距由大变小。因此, 该化石属于heteropolaren式, 即单极式中的典型异极(Dentzien-Dias et al., 2012)。依据这种特殊的螺旋结构, 推测鱼粪化石的产出者为鱼肠内具有较多螺旋瓣的鱼类, 如软骨鱼类中的鲨鱼或者是硬骨鱼中的肺鱼类等(McAllister, 1987)。
现在家养金鱼的粪便细如丝线, 个体较大的鲤鱼、胖头鱼等排泄物的直径也不过2~3 mm(王大锐和韩兆宽, 1999)。本次观察的所有鱼粪化石直径大小在11.5~20.0 mm之间, 故产生此类粪便的肯定是大型鱼类。因此, 推测在大安寨段沉积时期的淡水湖泊中生活着很多的大型鱼类。
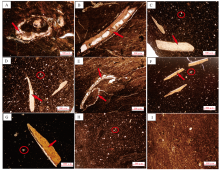
本次研究中, 将85号和86号鱼粪化石及其围岩制作成薄片进行镜下观察。鱼粪化石的围岩为粉砂质泥岩、泥页岩(图 5-A, 5-B, 5-I), 发育大量双壳类及介形类化石(图 5-E), 含大量黑色、炭化的植物碎片化石(图 5-I)。该围岩中含不规则长条状硅质成分, 显微镜下可见褐色物质及不规则浸染状有机质, 推测为破碎的鱼粪成分(图 5-A, 5-B)。
 | 图 5 川东北大槽剖面下侏罗统自流井组大安寨段鱼粪化石及其围岩显微特征 A— 85号鱼粪化石的围岩样品, 粉砂质泥岩, 含有介壳类生物化石(红色箭头所示), 正交偏光; B— 85号鱼粪化石的围岩薄片, 见生物介壳(红色箭头所示), 泥页岩, 单偏光; C, D— 85号粪化石的切片, 可观察到厘米级化石为磷质成分, 应为所食鱼类骨骼化石(红色箭头所示), 单偏光; E— 86号粪化石围岩的切片, 粉砂质页岩, 含介形虫化石(红色箭头所示), 正交偏光; F, G— 86号粪化石的切片, 见大量的磷质鱼刺结构(红色箭头所示), 部分鱼类骨骼较大(如G中红色箭头所示), 单偏光; H— 86号粪化石显微照片, 含较多的粉砂及黑色炭屑, 单偏光; I— 89号鱼粪化石的围岩薄片, 泥页岩, 单偏光Fig.5 Microscopic characteristics of coprolites and surrounding rocks of the Da’ anzhai Member of Lower Jurassic Ziliujing Formation in Dacao section of northeast Sichuan Basin |
从鱼粪化石的显微薄片中可以观察到, 粪化石的内含物具有较多的有机质、大量的粉砂及较多的黏土矿物, 且有机质大量富集(图 5-H); 还包括大量介形类化石, 与有机质混杂在一起(图 5-E)。另有大量的毫米级化石为磷质成分, 推测其为所食鱼类的未经消化或消化不彻底的鱼骨化石(图 5-D, 5-F, 5-G)。较多的褐色物质及均质矿物(图 5-C, 5-D, 5-F, 5-G, 5-H中红色圆圈内)的成分为磷质, 形态不规则, 推测为软体动物(虾、蟹等)的外壳。鱼粪中含有大量介形类化石和磷质鱼骨化石、软体动物外壳等, 表明鱼粪的生产者应为大型肉食性鱼类, 且尤其喜食小鱼类和软体动物等。这2个粪化石没有典型的螺旋纹特征, 推测生产者属于大型肉食性硬骨鱼类(据王大锐和韩兆宽, 1999)。
解剖学研究结果显示, 鱼肠内部多具有结构特殊的螺旋瓣, 未完全消化后的食物残渣通过肠道形成的粪便具有旋纹(王大锐和韩兆宽, 1999)。鱼肠内的螺旋瓣越密集, 鱼粪的螺旋纹就越多, 其中鲨鱼鱼肠的各旋转之间较为密集, 肺鱼较稀疏, 硬鳞鱼各旋转之间距离更远。硬骨鱼体内没有这种结构, 其粪便一般是短棒状(王大锐和韩兆宽, 1999)。但还是不能用来螺旋结构精确推断鱼类的种属, 因为各个种属鱼类的螺旋瓣差别很小, 有时候不同种属的螺旋瓣也会有相同的情况(尹赞勋, 1945)。
文中所研究的鱼粪化石直径在11.5~20.0 mm之间, 直径较大, 其中2个相对较小的鱼粪化石(85号及86号样品)不具完整的外形, 没有明显的螺旋特征, 推测为硬骨鱼类所产(据王大锐和韩兆宽, 1999)。鱼粪化石的内含物中含有小鱼的鱼骨化石和软体动物外壳, 可知其生产者为喜食小鱼和软体动物的大型肉食性硬骨鱼类。
呈现典型螺旋纹的鱼粪化石(89号样品)的生产者为体内具有较多螺旋瓣的大型鱼类, 如鲨鱼类、肺鱼类等(尹赞勋, 1945; 刘宪亭, 1953; 边兆祥等, 1991; 王大锐和韩兆宽, 1999; 龚一鸣等, 2009)。将研究区的鱼粪化石与以往文献中的粪化石比对发现, 其与鲨鱼的粪便外形相差较大, 但与桐梓、富县粪化石的形态相似。然而, 桐梓粪化石只有2条螺旋, 长25 mm、宽12.3 mm(边兆祥等, 1991), 与研究剖面鱼粪化石的大小相差也较大; 而富县粪化石长56.4 mm、宽40 mm、厚18.6 mm(刘宪亭, 1953), 产于陕西富县黑水寺侏罗系上部薄层石灰岩地层中, 其产出的大小、时代及围岩岩性与研究剖面大安寨段的鱼粪石较为接近。另外, 研究剖面大安寨段鱼粪化石具有5条明显的螺旋纹, 也较接近富县的粪化石。富县的粪化石经推测为侏罗纪肺鱼类所产, 且肺鱼类个体较大(刘宪亭, 1953)。肺鱼类在地史上延续的时间较长, 其化石在全球从早泥盆世一直到新生代的地层中均有发现(王俊卿, 1981)。中国迄今发现的相关化石主要产出于泥盆系、侏罗系和白垩系(刘宪亭, 1980; 王俊卿, 1981; 乔妥和朱敏, 2008)。三叠纪以后, 空棘鱼类绝大多数都进入海洋, 而肺鱼类则相反, 绝大多数进入淡水水域(王念忠, 1977a, 1977b)。中生代出现的肺鱼类中, 角齿鱼分布很广, 在多处侏罗系地层中发现有角齿鱼的齿板化石, 如四川广元的四川角齿鱼、江油的玲珑角齿鱼、威远的杨氏角齿鱼等(刘宪亭, 1980)。在中生代的河流、湖泊以及大小池塘中都有角齿鱼的踪迹(马凤珍, 1981)。本次化石的产出地址和时代与已发现的角齿鱼化石的产出地很接近, 都产自四川盆地侏罗系, 据此推测螺旋状鱼粪化石的生产者为肺鱼目角齿鱼科的概率很大。肺鱼类的推测也与大安寨段沉积时期的淡水湖泊环境相对应(王念忠, 1977a, 1977b)。
由于粪化石属遗迹化石, 基本上为原地原位保存, 并且粪化石几乎没被破坏, 故文中所研究的鱼粪化石和围岩为同时代产物(龚一鸣等, 2009), 即化石的时代和大安寨段同属于托阿尔期大洋缺氧事件时期。鱼粪化石的围岩为黑色页岩, 表明沉积时为还原环境, 属半深湖或深湖区域。泥页岩中含有大量的植物叶片化石, 表明当时植物繁盛、气候温暖潮湿、降水丰富。鱼粪化石的内含物中含有大量软体动物和小型鱼类, 表明当时气候适宜生物生存。而在野外剖面中, 大安寨段介壳灰岩与泥页岩呈间互产出, 灰岩中发现较大个体的淡水双壳类实体化石, 表明当时的沉积环境为含氧量丰富的淡水环境。
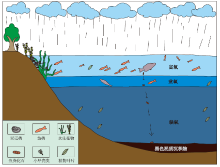
鱼粪化石寄主地层的还原色(灰黑色)形成于高生产力的湖泊沉积环境和还原的早期成岩环境(龚一鸣等, 2009)。研究剖面大安寨段沉积时期, 气候适宜, 降水丰富, 有利于湖泊的生物多样性。由湖泊中有大型肉食性鱼类的存在可推测湖泊中还生活着各种小型的草食性、杂食性鱼类和其他肉食性鱼类等, 已发现的大量介壳类化石、软体动物等表明了消费者丰富多样。高级消费者的存在也显示了食物链的复杂多样性, 据此也可推测湖泊表层是富氧的, 适于各种湖泊动物生存。围岩中植物碎片丰富, 显示研究区早侏罗世开放湖泊中有河流淡水及陆源物质供应, 其不仅带来了植物碎片等有机质, 也将输入大量溶解物质作为营养盐提供给海洋表层藻类等初级生产者消费。在深层湖泊中沉积下来的各种生产者和消费者的尸体以及消费者的排泄物等, 因湖泊底层的缺氧还原环境而保存下来; 且在湖泊底层的缺氧环境中没有分解者, 造成鱼粪等含有丰富有机质的物质很难通过分解者进行分解, 从而得以完整保存, 由此构成了早侏罗世托阿尔期复杂的大型湖泊生态系统。因此, 在托阿尔期早期大洋缺氧事件期间, 研究区湖泊顶层是富氧的复杂生态系统, 而底层为缺氧的粪化石埋藏环境(图 6)。
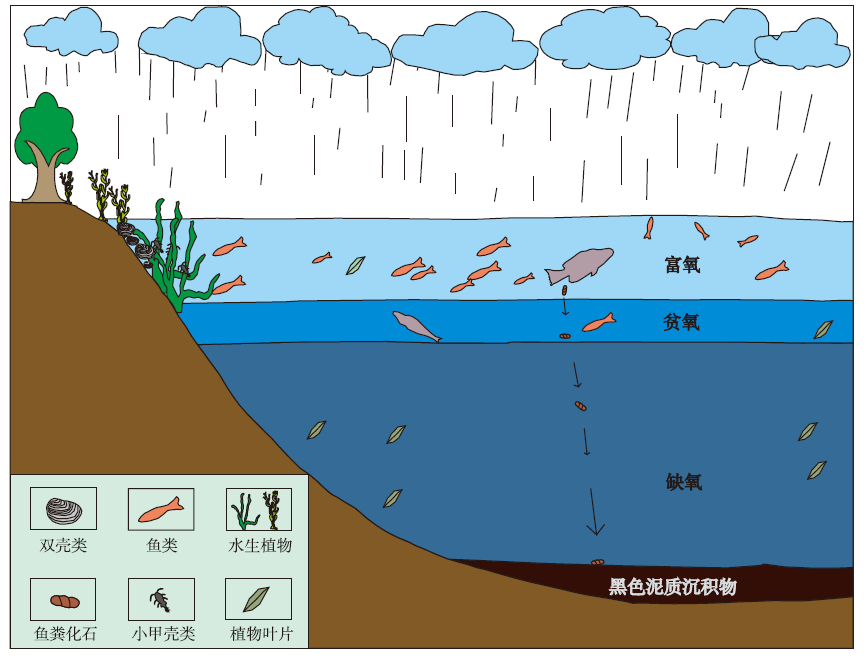 | 图 6 川东北早侏罗世自流井组大安寨段沉积时期湖泊生态系统(据Megard, 1967, 1968; 有修改)Fig.6 Lake ecosystem during the depositional period of Da’ anzhai Member of Lower Jurassic Ziliujing Formation in northeast Sichuan Basin(modified from Megard et al., 1967, 1968) |
1)川东北地区大槽剖面大安寨段鱼粪化石的直径较大, 推测其中2个相对较小鱼粪化石的生产者为喜食小鱼和软体动物的大型肉食性硬骨鱼类, 而呈现典型单极式螺旋纹的鱼粪化石生产者为肺鱼类中的角齿鱼科。
2)早侏罗世托阿尔期大洋缺氧事件发生时, 川东北淡水湖泊下层水体为缺氧、还原的环境, 而表层水体是富氧的, 适宜鱼类、软体动物、双壳类、介形类动物生存, 形成了主要由生产者及包含大型肉食性鱼类的消费者构成的具有复杂食物链结构的大型湖泊生态系统。
致谢 参加野外工作的还有意大利帕多瓦大学的Nereo Preto、意大利蒂利亚斯特大学Marco Franceschi以及成都理工大学陈彬、韩璐、银河、王小端。感谢Nereo Preto教授在野外工作中给予的指导和帮助, 感谢成都理工大学沉积地质研究院李云博士在室内研究中提供的帮助!
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|