

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
贵州开阳龙水地区震旦系洋水组沉积特征及生物成磷作用*
[吴文明1, 2, 3  , 杨瑞东
, 杨瑞东1 , 刘建中3, 4 , 王泽鹏2, 3 , 罗朝坤1 , 叶太平5 , 冯康宁1 , 杨光海1 , 汪龙波1 , 刘松2 , 王大福2 , 黄毅2 , 潘启权2 , 张丞2 ]
, 杨瑞东, 刘建中|
|

第一作者简介 吴文明,男,1984年生,博士研究生,矿床学及地球化学专业。E-mail: 272180889@qq.com。
贵州开阳磷矿以其厚而富的磷矿石资源备受关注。对开阳龙水地区含磷岩系(洋水组)剖面进行地层学、岩石学和沉积学研究,并讨论其沉积环境和成磷作用。区内含磷岩系岩性组合为黏土岩—白云岩—碎屑状磷块岩—硅质岩—泥晶磷块岩—叠层石磷块岩。龙水地区在震旦纪陡山沱期整体为靠近古陆边缘的滨岸潮坪沉积环境,并经历了2次海侵—海退。其沉积环境由初期的无障壁海滩转变为晚期的潮坪环境,表明陡山沱期古海洋处于较为动荡的环境,海平面升降频繁。富含生物化石的致密状泥晶磷块岩、圆柱状叠层石磷块岩和磷质生物团粒,均属于原生磷块岩。三者磷酸盐含量极高,属于高品位磷块岩,其形成环境可能是弱酸性至弱碱性的浅海陆棚。生物与高品位磷块岩的密切关系说明生物在高品位磷块岩形成过程中起重要作用。其形成具有3个必要条件: 海水富磷,生物大量繁殖,有利于生物繁殖及磷酸盐结晶的沉积环境。
, Yang Rui-Dong, Liu Jian-ZhongAbout the first author Wu Wen-Ming,born in 1984,is a Ph.D. candidate of Guizhou University. He is mainly engaged in sedimentary deposits and solid mineral exploration. E-mail: 272180889@qq.com.
The Kaiyang phosphate mine has attracted much attention due to its thick and rich phosphate ore resources. In this paper,stratigraphy,petrology and sedimentology are studied for the phosphorous-bearing Yangshui Formation in Longshui,Kaiyang,and the sedimentary environment and phosphorus formation are discussed. The lithologic assemblages of the phosphorous-bearing rock systems in the area are clay rock-dolomite-clastic phosphate rock-siliceous rock-micritic phosphate rock-stromatolite phosphate rock. In the Doushantuo age,the Longshui area was a coastal tidal flat close to the old landmass,and experienced transgression-regression twice. The sedimentary environment of the Doushantuo age changed from the barrier-free beach in the early stage to the tidal flat in the late stage,indicating a relatively dynamic environment with frequent sea-level fluctuations. The dense micritic phosphate rock,cylindrical stromatolite and phosphate biological aggregates rich in biological fossils belong to the original phosphate rock. The phosphate content of the three lithofacies is high,belonging to the high grade phosphate block rock. The depositional environment was likely a low-energy subtidal zone with medium-acidity to weak alkalinity. The close relationship between organisms and high grade phosphate rock suggests that organisms played an important role in the formation of high grade phosphate rock. Its formation has three prerequisite conditions: seawater rich in phosphorus,biological mass reproduction and conducive to biological reproduction and phosphate crystal environments.
贵州开阳磷矿是世界上典型的沉积型磷块岩成矿区, 由于其磷块岩沉积厚度大、品位高而闻名。磷矿层赋存于震旦系洋水组(与陡山沱组同期异相)(刘鸿允和沙庆安, 1963)。以往的成磷作用主要有生物成矿说、无机沉淀说、交代说和物理富集说等。关于生物成磷, 前人从岩石学、矿床地球化学和模拟化学实验等方面进行了论述。岩石学方面, 各地质时期磷矿成矿带中广泛分布有磷质叠层石(朱士兴和王砚耕, 1982; Martí n-Algarra and Sá nchez-Navas, 2001; 张伟等, 2015)、微体化石组合(袁训来, 1999; Liu et al., 2014)、藻类化石(赵东旭, 1986, 1990; 张昀和袁训来, 1995; Chen et al., 1995)和动物胚胎化石(Xiao et al., 1998), 说明磷块岩的形成与生物密切相关。地球化学方面, 瓮安及开阳地区磷块岩富集了Cr、Co、Pb、Ga及Sr等与生物密切相关的微量元素(郭庆军等, 2003; 施春华, 2005; 邓克勇等, 2015), 其中亲硫元素Pb表现最为明显; 瓮安地区磷块岩稀土元素配分模式呈现了中稀土略富集的“ 帽型” 配分模式, 表明了微生物对成矿的作用(陈其英等, 2000; Liu et al., 2014; 杨海英等, 2020; 张亚冠等, 2020)。陡山沱期海水的碳同位素发生负漂移, 磷块岩中的δ 13C平均为-1.76× 10-3, 远低于与之共生的白云岩(0.985× 10-3), 磷块岩中比白云岩较低的δ 13C值表明碳来源为生物
在中国地质调查局项目“ 贵州省开阳以东磷矿整装勘查区矿产调查与找矿预测” 实施过程中, 在瓮安县和开阳县交界的开阳县龙水乡发现了沉积序列自下而上依次为黏土岩— 白云岩— 碎屑状磷块岩— 硅质岩— 泥晶磷块岩— 叠层石磷块岩的含磷岩系剖面, 特别是发现了致密状富含生物化石的泥晶磷块岩(生物磷块岩), 对深入研究其成因及认识开阳地区高品位磷矿石的成因研究具有重要意义。黔中磷矿经历了由海平面变化影响导致的复杂成矿过程, 但对原生的生物成矿作用研究欠缺。作者对该剖面开展了岩石学、沉积学和生物特征等方面的研究, 重点研究生物成磷作用。
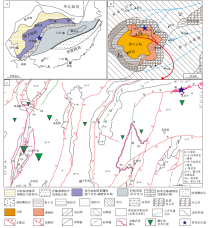
研究区大地构造位于扬子陆块区上扬子陆块鄂湘渝黔前陆褶皱冲断带黔北隆起务川— 开阳北北东向变形区(贵州省地质调查院, 2017)。该区域在陡山沱期总体位于扬子陆块陆架边缘(Jiang et al., 2011)(图 1-a), 并处于黔中古陆北缘的无障壁滨岸浅滩环境(陈国勇等, 2015; 国土资源部矿产勘查办公室, 2015; 吴文明等, 2017; Zhang et al., 2019)(图 1-b)。贵州开阳富磷矿聚集区主要构造为走向近南北的洋水背斜、翁昭背斜、近南北向的逆冲断层以及系列北东东的断裂(图 1-c), 研究剖面位于北东东走向的龙水断层以南的龙水乡。区内出露地层由老到新依次有青白口系清水江组变余粉砂岩、南华系澄江组陆源碎屑岩、下震旦统洋水组(与陡山沱组同期异相)黏土岩— 白云岩— 磷块岩— 硅质岩— 磷块岩岩性组合、上震旦统灯影组白云岩等地层。
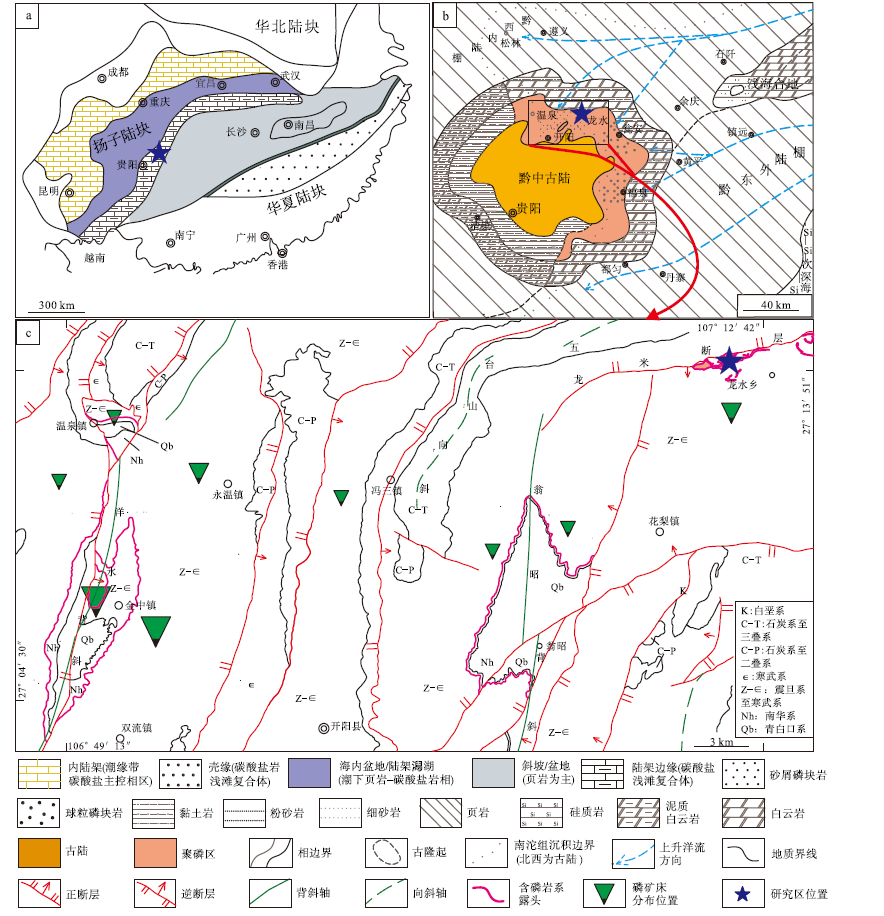 | 图 1 贵州开阳地区区域地质背景图 a— 扬子陆块陡山沱期岩相古地理图(据Jiang et al., 2011; 有修改); b— 黔中地区陡山沱期岩相古地理图(据陈国勇等, 2015; 有修改); c— 开阳地区地质简图①②Fig.1 Regional geological background of Kaiyang, Guizhou Province |
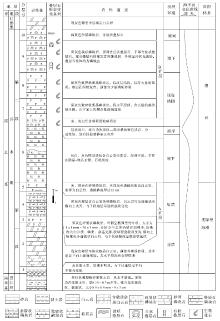
本次研究的开阳龙水地区洋水组剖面, 由位于小河口处的第1— 5层(坐标: E107° 13'33″, N27° 14'29″)、姊妹岩处的第6层(坐标: E107° 13'20″, N27° 14'31″)和大坪山附近的第7— 10层(坐标: E107° 11'23″, N27° 14'05″)3个区段组合而成。根据岩性组合, 洋水组可分为上下2段(图 2)。第一段为碎屑岩— 白云岩— 磷块岩建造(图 2中第1— 5层), 厚度为6.7 m。其下部(第1层)为灰色薄层含白云质黏土岩; 中部(第2层)为浅灰色中厚层白云岩, 具水平层理、交错层理; 上部(第3— 5层)自下而上分别为深灰色碎屑状、条带状和砂屑磷块岩, 为A矿层。第二段为硅质岩— 磷块岩建造(图 2中第6— 10层), 厚度大于11 m。其下部(第6层)为含硅质团块白云岩、含磷质团粒硅质岩, 夹砾岩、砂砾岩(其位于剖面的具体位置有待进一步研究); 上部(第7— 10层)自下而上分别为深灰色厚层致密状泥晶磷块岩、圆柱状叠层石磷块岩、丘状叠层石磷块岩和波状叠层石磷块岩, 为B矿层。与开阳龙洞沟剖面(盛章琪, 1984)及瓮安大塘矿段剖面(密文天等, 2010)类似, 富含生物的磷块岩均分布于含磷岩系上部(图 3)。
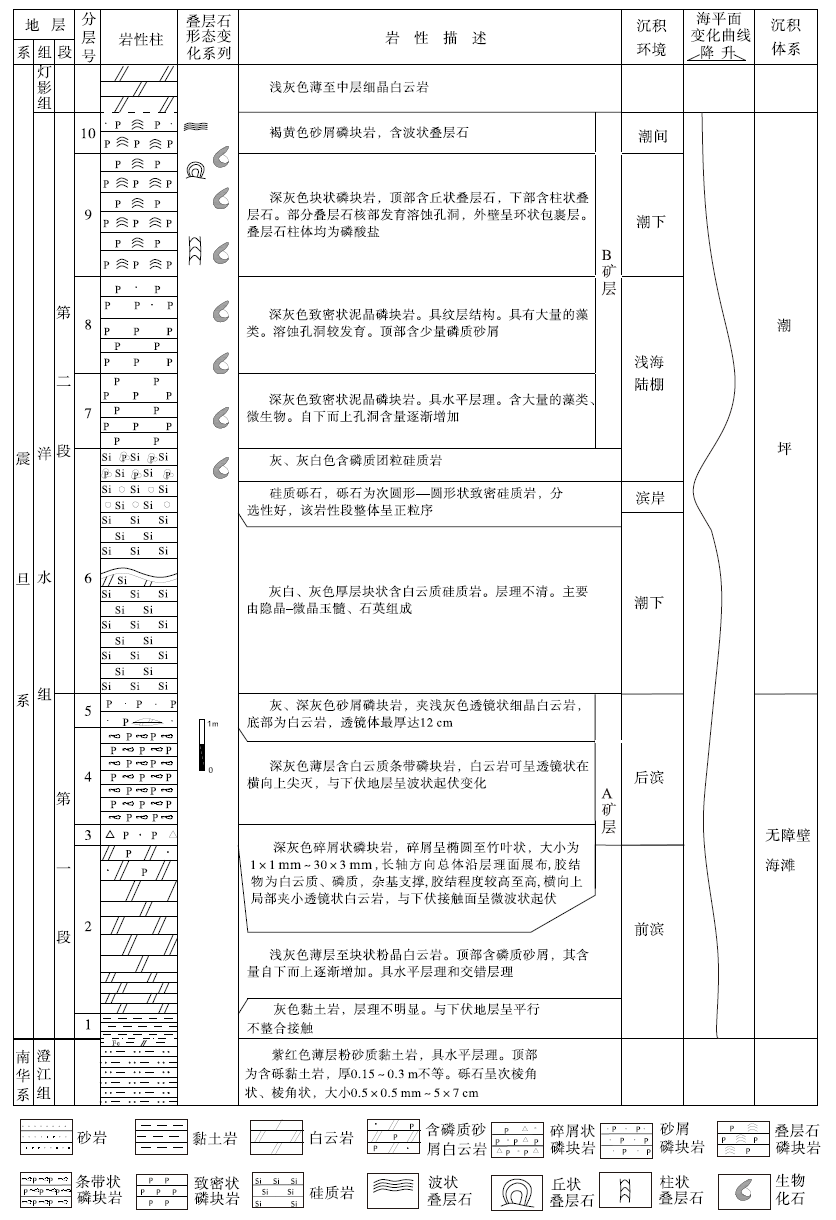 | 图 2 贵州开阳龙水地区震旦系洋水组综合柱状图Fig.2 Synthetic column of the Sinian Yangshui Formation in Longshui area, Kaiyang, Guizhou Province |
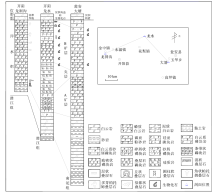
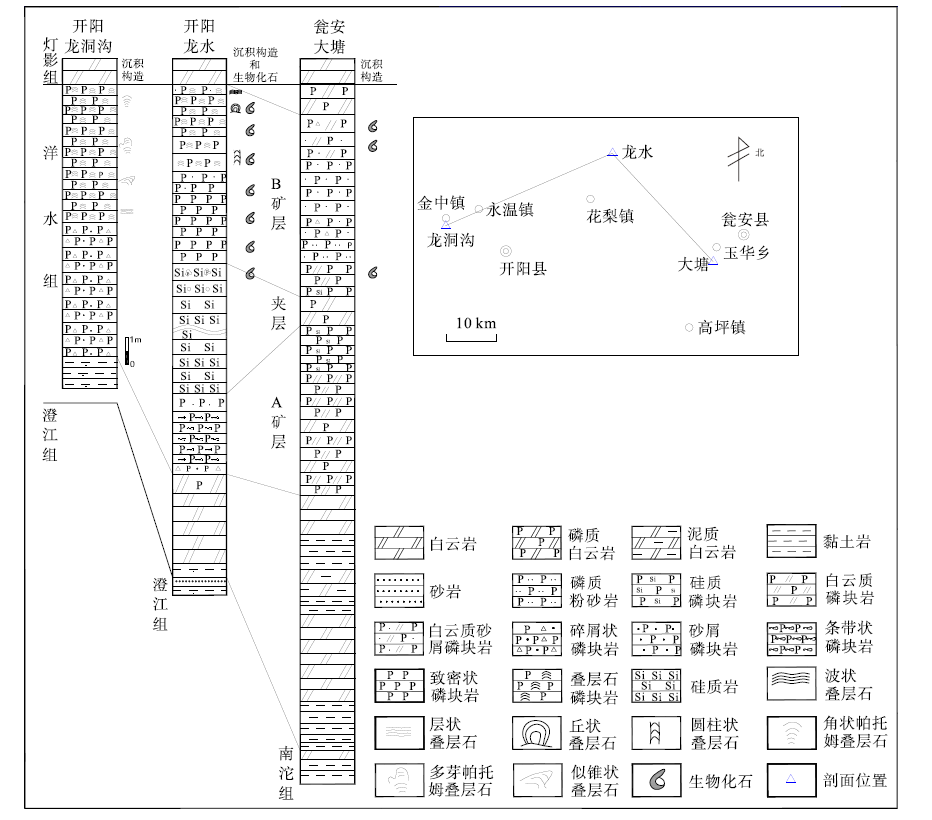 | 图 3 贵州开阳、瓮安震旦系洋水组地层对比 开阳龙洞沟剖面(盛章琪, 1984), 瓮安大塘矿段剖面(密文天等, 2010)Fig.3 Stratigraphic correlation of the Sinian Yangshui Formation in Kaiyang and Weng’ an areas, Guizhou Province |
开阳龙水地区矿石自然类型有致密块状磷矿石、砂屑状磷矿石、条带状磷矿石和团块状磷矿石等。B矿层以致密块状及砂屑状磷矿石为主, 条带状磷矿为次, 再次为团块状磷矿石; A矿层以条带磷矿石及致密块状磷矿石为主, 其次为砂屑状磷矿石, 再次为团块状磷矿石。矿石的工业类型属碳酸盐型白云质磷块岩。
2.2.1 碎屑结构
1)角砾结构。主要分布在A矿层底部(第3层), 为角砾状磷块岩(图 4-a)。其角砾由磷质砂屑紧密堆积而成, 呈长条状、竹叶状、棱角状和次棱角状, 直径2~30 mm, 分选性较差, 磨圆程度较低, 角砾含量大于30%。胶结物以磷质为主, 白云石次之。以杂基支撑为主。
 | 图 4 贵州开阳龙水地区洋水组剖面典型岩石结构照片 a— A矿层底部的角砾状磷块岩; b— A矿层下伏的含磷质砂屑白云岩; c— 夹层上部的砾岩; d— 夹层上部的砂砾岩; e、f— 夹层顶部的含磷质团粒硅质岩; g、h— 泥晶磷块岩显微镜下的生物结构; i— 磷质团块显微镜下的生物结构Fig.4 Typical rock features of the Yangshui Formation section in Longshui area, Kaiyang, Guizhou Province |
2)砂砾结构。分为砂状结构和砾状结构。砂状结构主要是指分布于洋水组底部白云岩的顶部(第2层, 图 4-b)、A矿层中上部(第4、5层)和B矿层中部(第8层顶部)的砂屑磷块岩。其砂屑主要由隐晶质磷灰石矿物组成, 呈不规则椭圆状为主, 大小多在0.1~0.3 mm之间, 局部具长轴定向分布。砂屑边缘多具层纤状磷灰石分布。粒间胶结物主要为隐晶质胶状胶磷矿, 局部为白云石。少量白云石呈自形— 半自形粒状, 粒径0.01~0.2 mm, 聚集呈条带状分布粒间胶结物中。石英呈他形粒状, 粒径小于0.4 mm, 星散分布于磷质砂屑中, 少量分布于粒间胶结物中。黄铁矿呈自形— 半自形粒状, 粒度小于0.1 mm, 呈星点状或聚集成细小团块状分布于粒间胶结物中。少量水云母呈细小鳞片状, 粒度(长轴)小于0.05 mm, 零星分布于粒间胶结物中。砾状结构主要指分布在硅质白云岩上部的砾石层(第6层上部, 图 2-c)。砾石成分主要为硅质, 少数为白云质。角砾呈圆形、椭圆形, 磨圆度较好, 直径0.5~20 cm, 分选性较好。多为颗粒支撑, 砾间充填细小砂砾、泥晶质胶结物。根据砾石大小, 可分为中砾岩和细砾岩。根据其成分较单一和分选性好的特性, 应属于滨岸砾岩。
2.2.2 生物结构
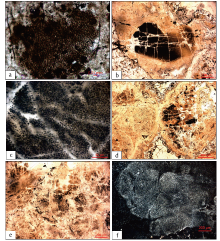
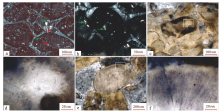
指B矿层的致密状泥晶磷块岩、圆柱叠层石磷块岩和含磷质团粒硅质岩中分布的大量生物结构。前两者在显微镜下可发现大量红藻类和绿藻类化石(图 4-g, 4-h, 4-i), 以红藻类多细胞叶状体(Zhang, 1989)为主(图 5)。叶状体直径从几微米至数毫米不等。其内部细胞直径小于10 μ m, 呈定向或非定向排列。生物化石含量大于70%。此外, 其还含有部分单细胞藻体, 根据其形态判断, 它们可能为红藻和绿藻。含磷质团粒硅质岩的磷质团粒呈灰白至深灰色, 块状构造, 该层厚度约0.6 m。其磷质团粒为白色, 呈近圆球状(图 4-i)或不规则云团状, 直径0.3~10 mm。团粒或孤立或呈集合体分布。团粒间的充填物为硅质岩(石英和玉髓)。部分团粒内包含定向或非定向排列的细胞颗粒。这种结构是磷酸盐化藻类生物颗粒被硅质胶结形成。
 | 图 5 贵州开阳龙水磷矿致密状泥晶磷块岩和圆柱状磷块岩的红藻类生物显微结构照片(采自龙水大坪山) a— 具细胞岛的红藻; b— 具无定向分布细胞结构的红藻; c— 照片b的局部放大; d— 两种不同结构的红藻体, 右部具细胞结构, 左部未见细胞; e— 具放射状细胞结构的红藻体; f— 呈扇子状的红藻体Fig.5 Microstructure of algal of dense micrite phosphorus block and cylindrical phosphorus block in Kaiyang Longshui phosphate ore, Guizhou Province(from Longshui Dapingshan) |
2.3.1 水平层理
主要分布于B矿层泥晶磷块岩底部(图 6-a), 由深灰和浅灰色泥晶磷块岩互层组成, 单层厚度为2~15 cm。剖面底部的白云岩层也发育有水平层理(图 6-b)。
 | 图 6 贵州开阳龙水地区洋水组剖面典型岩石沉积构造照片 a— 具水平层理的泥晶磷块岩; b— 具水平层理和交错层理的细晶白云岩; c— 具交错层理的条带状磷块岩; d— 具类似鸟眼构造的泥晶磷块岩; e、f— 波状叠层石; g、h、i— 柱状叠层石Fig.6 Photographs of typical sedimentary structure of rock in the Yangshui Formation in Longshui area, Kaiyang, Guizhou Province |
2.3.2 交错层理
主要分布于剖面底部的白云岩层(图 6-b)和A矿层中部的白云岩质条带状磷块岩(图 6-c)。
2.3.3 鸟眼构造
主要分布于B矿层泥晶磷块岩上部(图 6-d)以及圆柱状叠层石柱体内部(图 6-i), 呈不规则孔洞大致沿层理面和叠层石凸起纹层定向排列, 呈近圆状、不规则状, 其边缘一般呈弧状。其孔径0.2~5.0 mm, 横向延伸最大达40 mm。这可能是红藻类生物遗体等有机质腐烂分解后形成的。圆柱状叠层石柱体仅内部发育孔洞, 而其外壁未见, 说明其成因与沉积暴露无关。
2.3.4 叠层石构造
1)圆柱状叠层石。主要分布于含磷岩系B矿层(第9层)中部。叠层石(图 6-g, 6-h, 6-i)为浅灰至深灰色, 呈圆柱状, 具有外壁。柱体高为0.3 m以上, 直径为4~8 cm。柱体纹层不发育。叠层石与围岩界线明显。柱体之间由强硅化含砂质砂屑磷质岩充填。充填物由磷质砂屑和少量陆源碎屑石英组成。磷质砂屑呈不规则椭圆状为主, 大小多在0.2~1.0 mm之间。碎屑石英呈磨圆状, 粒度0.2~0.6 mm, 零星分布。胶结物为磷质, 后期硅化较强。胶结程度一般, 往往被风化淋滤, 形成蜂窝状孔洞或空槽。叠层石柱体可能形成于水体比较深的区域, 而柱间充填物可能形成与水动力较强的潮下带。
2)波状叠层石。主要分布于含磷岩系B矿层(第10层)顶部。叠层石(图 6-e, 6-f)纹层形态为小型波纹状, 波纹不规则。单个纹层呈浅灰色, 厚度约为2 mm, 近于等厚叠置。少量自生石英呈自形— 半自形粒状, 粒度小于0.1 mm, 呈星点状或聚集成不规则团块状、条带状分布。少量黏土矿物水云母呈细小鳞片状, 零星分布。
根据贵州省开阳县龙水磷矿普查报告(贵州地质局106队, 1983)①(贵州省地质局106队. 1983. 贵州省开阳县龙水磷矿普查地质报告.), 区内主要磷矿体主要赋存于A矿层和B矿层。B矿层为区内主要矿层, 除局部沉积尖灭外, 在全区发育良好, 走向延伸长达17.5 km。其厚度变化较大、分布零星。矿体厚度0.69~17.99 m, 平均厚度3.90 m, 矿石P2O5含量为8.05%~40.08%, 平均33.24%。受区内复杂构造影响, 共分布大小矿体27个。A矿层位距上部B矿层3~12 m, 矿层分布极不稳定, 其厚度和品位变化很大, 矿体小。厚度0.60~3.55 m, 矿石P2O5含量为12.18%~30.60%。
龙水地区洋水组由于受富禄间冰期时复杂古地理地形控制, 沉积厚度在各局部区域有很大差异。洋水组一段总体为无障壁海滩环境, 其与洋水背斜一带的磷矿层类似, 以砂屑磷块岩为主(张亚冠等, 2016)。其下部的中层至厚层泥晶白云岩具水平层理和斜交层理, 应为前滨环境。中部的磷质砾屑呈长条状和竹叶状, 为高能水动力环境产物。上部主要以白云质条带状磷块岩为主, 条带状磷块岩被认为是海平面不断变化、导致海水化学条件不断变化引起磷质和白云质的交互沉积(柴华等, 2014); 当海水磷酸盐含磷充足时, 沉积磷质条带、磷质沉积物受水流破碎影响形成磷质颗粒堆积; 当海水磷质输入不足时, 会形成白云质沉积, 磷质颗粒会受白云质胶结, 且在磷质条带沉积间歇发育白云质条带沉积。顶部为砂屑磷块岩, 砂屑磷块岩一般为原生磷块岩受高能波浪或潮汐水流破碎、簸选、搬运和再沉积的产物(叶连俊等, 1989)。
洋水组二段总体为潮坪环境。其下部的含白云岩团块硅质岩夹层总体为潮下环境, 但顶部的硅质砂砾岩表明该时段海平面下降, 致使该区变为滨岸砂砾滩环境。推测花梨— 龙水一带存在半岛。中下部的致密状泥晶磷块岩(生物磷块岩)具水平纹层, 显示其可能形成于浅海陆棚; 其上部为叠层石磷块岩, 自下而上为柱状叠层石— 丘状叠层石— 波状叠层石序列。根据叠层石柱体的形态可判断其形成环境。柱状叠层石和丘状叠层石一般出现在潮下带(Reid et al., 2011; 梅冥相和孟庆芬, 2016)。波状叠层石一般出现在潮间带(Delfino et al., 2012)。B矿层自下而上出现的致密状泥晶磷块岩— 柱状叠层石— 丘状叠层石— 波状叠层石序列, 展示了该时段海水深度由深变浅、沉积环境由浅海陆棚过渡为潮间带的变化过程。
从整个剖面岩相组合特征来看, 该地区在陡山沱期整体为靠近古陆边缘的滨岸潮坪沉积环境, 并经历了2次海侵— 海退。其沉积环境总体经历了无障壁海滩到潮坪环境的变迁。表明陡山沱期古海洋处较为动荡的环境, 海平面升降频繁(密文天等, 2010; 杨爱华等, 2015)。
龙水剖面B矿层的致密状泥晶磷块岩, 包括层状泥晶生物磷块岩和柱状叠层石磷块岩, 岩石坚硬, 质地紧密。两者在显微镜下均显示为隐晶质胶磷矿, 磷酸盐含量大于95%。说明这些磷块岩没有经过后期水动力破坏, 应为原生磷块岩。夹层顶部含磷质团粒硅质岩中的团粒多具有保存完整的藻类生物细胞结构。开阳洋水背斜大量的颗粒磷块岩, 是构成高品位磷块岩的主体。颗粒中也发现大量的微生物结构, 可能是生物磷块岩被改造形成的。因此, 生物成磷作用是初始磷块岩形成的重要作用, 对形成高品位、巨量磷块岩具有重要的控制作用。
4.1.1 矿石结构指示的环境
泥晶磷块岩通常被认为是磷灰石自生沉降形成的原生富磷沉积产物(刘魁梧和陈其英, 1994)。研究区的泥晶磷块岩发育水平纹层, 指示其沉积环境为较低能的浅海陆棚。
4.1.2 生物指示的环境
开阳龙水地区的B矿层富含生物化石, 与瓮安地区的B矿层相似, 均富含红藻类化石(陈孟莪和刘魁梧, 1986)。致密状泥晶磷块岩、柱状叠层石磷块岩及含磷质团粒硅质岩中含大量磷酸盐化红藻(图 6)或绿藻化石, 红藻类占比大于70%。厚度如此大的原生磷块岩与生物密切相关, 说明以红藻为主的生物群的生活环境很适合磷酸盐的沉积和结晶。现代藻类学研究认为红藻主要生活在30~60 m深水域, 最深可达200 m(段德麟等, 2012)。地质时代早期的红藻生活在弱酸性水体中(Oliveira and Bhattacharya, 2000)。
4.1.3 矿物组合指示的环境
致密状泥晶磷块岩与圆柱状叠层石磷块岩均由隐晶质磷灰石和自形石英晶体组成。从这些石英晶体与磷酸盐矿物的紧密程度来看, 两者可能是同生或准同生沉积物。这表明原生磷块岩的形成环境既有利于磷灰石的结晶, 也有利于石英晶体的形成。沉积磷灰石一般形成在浅水区域, 溶解状态的磷酸盐在水体pH值达到8± 0.5时可大量沉积(朱士兴等, 1982)。晶型很好的自生石英, 可能是沉积磷块岩短暂暴露地表、准同生淋漓作用形成的产物, 说明磷块岩沉积环境属于水体较浅的潮坪环境。
经过光学显微镜下观察, 开阳龙水地区内发育的致密状泥晶磷块岩和圆柱状叠层石磷块岩均为富含生物化石的隐晶质磷酸盐; 含磷质团粒硅质岩的磷质团粒为藻类生物化石。它们属于初始成磷作用的原生磷块岩, 或者改造的生物磷块岩。这表明研究区磷块岩的形成与生物有着极其密切的关系。
前人认为生物成磷作用有5种方式: (1)生物吸收介质中的磷, 形成磷质菌藻细胞或磷质介壳, 死亡后直接堆积成矿(蒋志文, 1982; 陈其英, 1990); (2)生物体被磷酸盐化, 随后机械富集成矿, 形成研究区内的磷质团粒, 以及瓮安地区以磷酸盐化球形藻、湖北宜昌地区以磷酸盐化兰绿藻丝构成的核形石(赵东旭, 1990)和秘鲁海岸鸟粪化石、鱼骨化石等(Fö llmi, 1996; Filippelli, 2011); (3)藻类活动可以捕捉、粘黏各种磷质碎屑, 形成各种叠层状磷块岩(朱士兴和王砚耕, 1982; 王砚耕和朱士兴, 1984; 赵东旭, 1990)和颗粒磷块岩(She et al., 2013); (4)藻体在沉积物中分解使微环境成碱性, 促使磷酸盐结晶(朱士兴等, 1982; 赵东旭, 1990), 与磷酸盐化生物化石一起形成研究区内的泥晶磷块岩; (5)藻类的光合作用需要从水介质中吸收大量的CO2, 从而使水介质中的pH值不断增高, 当pH值达到8± 0.5时, 溶解状态的磷酸盐就大量沉积(朱士兴等, 1982)。
笔者认为研究区高品位原生磷块岩的形成具有3个必要条件: 海水富磷, 生物大量繁殖和有利于生物繁殖与磷酸盐结晶的沉积环境。三者缺一不可, 相互作用。陡山沱期是生物大爆发的时期, 目前已发现3个大的生物群, 即瓮安生物群、苗河生物群和兰田植物群。其中瓮安生物群不仅包含具细胞组织结构和组织分化的多细胞藻类原叶体、多细胞藻类集合体、丝状藻类、球状藻类、疑源类等化石, 还包括后生动物休眠卵和胚胎化石以及早期后生动物的遗体或遗迹等化石。据作者初步研究认为, 龙水地区磷块岩中的生物化石种类也极为丰富。海水中高浓度的磷和营养物质为这些生物群的繁衍提供了物质保障。在有利的沉积环境中, 生物可能以捕捉、粘粘磷质、改变水体介质促使磷酸盐大量沉淀成矿。
根据致密状泥晶磷块岩具有的水平层理构造以及富含红藻类化石, 该类型磷块岩品位高, 形成于上升洋流携带深部的磷等营养物质大量滞留和贮藏于陆架边缘的浅海陆棚地带。该地带极利于藻类生物繁殖, 在藻类生物化学作用下, 大量生物被磷酸盐化并伴随巨量磷质沉淀结晶形成生物磷块岩。
开阳地区磷块岩具有高品位特征, 特别是具有磷质包壳的砂屑磷块岩含磷最高(吴文明等, 2017)。通过对开阳永温磷矿的砂屑磷块岩研究, 发现砂屑中含有藻类生物化石(图 7-a, 7-b), 部分地区的磷质砂屑具有生物结构(图 7-c, 7-d); 净砂屑磷块岩中还保留了类似生物磷块岩中的自形石英晶体(图 7-a, 7-b)。这说明砂屑磷块岩是生物磷块岩改造形成的。因此, 开阳地区高品位磷块岩的形成, 均与初始生物磷块岩密切关系, 说明藻类生物化学作用是主要成磷控制因素。
 | 图 7 贵州开阳地区砂屑磷块岩显微结构照片 a、b— 磷质砂屑中的藻体(红色箭头)和自形石英晶体(绿色箭头), 正交偏光, 样品采自开阳永温; c— 疑似为单细胞藻体的磷质砂屑, 单偏光, 样品采自开阳翁昭; d— 照片c局部放大, 疑似细胞核; e— 具有疑似放射状鞭毛结构的磷质砂屑, 单偏光, 样品采自开阳翁昭; f— 照片e的局部放大Fig.7 Microstructure of sands phosphorus rock of the Kaiyang Yongwen area, Guizhou Province |
1)贵州开阳龙水地区在陡山沱期整体为靠近古陆边缘的滨岸潮坪沉积环境, 并经历了2次海侵— 海退。其沉积环境由初期的无障壁海滩转变为晚期的浅海陆棚, 表明陡山沱期古海洋处于较为动荡的环境, 海平面升降频繁。
2)富含生物化石的致密状泥晶磷块岩、圆柱状叠层石磷块岩均属于原生磷块岩, 含磷质团粒硅质岩中的团粒是磷酸盐化生物化石。三者磷酸盐含量极高, 属于高品位磷块岩, 其形成环境为藻类生物极为繁盛的弱酸至弱碱性浅海陆棚。
3)开阳地区高品位磷块岩形成与藻类生物化学作用密切相关, 生物在高品位磷块岩形成过程中起重要作用。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|