1866年Ernst提出生态学(Ecology)的概念, 1935年Tansley(1935)提出生态系统概念。今天, 对于生态学的理解已然发生改变, 理解海洋珊瑚生态系统对于人类的意义是其最大的进步。
珊瑚是海洋环境的晴雨表, 也是最好的天然实验室。作为珊瑚的宿主和化石形态及其表征, 珊瑚礁是死亡珊瑚与活体珊瑚的复合体, 物化为岛、礁、滩、沙, 反哺孕育繁衍珊瑚, 形成珊瑚-珊瑚礁生态系统。几十万种游动的、原地的和死亡的动植物, 以其简单而神奇的关系, 构成这个星球最为庞大的生态系统。作者简要介绍中国南海地区的珊瑚-珊瑚礁, 提出划分南海珊瑚-珊瑚礁为中央区和周缘区2个分布区, 系统报道了2个航次(2009年和2019年)地质调查的发现, 包括科学定名46种六放石珊瑚和6种八放软珊瑚等成果, 阐述了单体环礁和复合环礁的特征及赋存、分布, 指出永乐环礁是南海唯一一个真正的切合达尔文模式的环礁, 构成现代海洋珊瑚-珊瑚礁形成演化研究最好的天然实验室。期望以此抛砖引玉, 推进珊瑚-珊瑚礁的调查与研究。
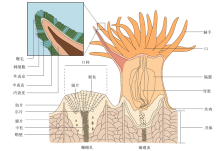
1 珊瑚、珊瑚礁1.1 什么是珊瑚、珊瑚礁珊瑚是古波斯语 sanga和今波斯语xuruhak的汉译, 是动物活体, 可原地活动但不能移动, 个体很小但群体很大。狭义珊瑚指捕食海洋浮游生物最古老最弱势最庞大的低等腔肠动物, 它们原地生长, 以多姿(形态)和多彩(色彩)构成地球最为庞大的海洋生态系统。广义珊瑚泛指珊瑚虫(图 1)与其体内共生虫黄藻分泌体和内色素形成五彩缤纷实物及其化石骨骼的沉积物, 亦即珊瑚依靠自身生物作用、光合作用、钙化作用、埋藏化作用和化石化作用不断长高, 由此孕育繁衍形成的珊瑚礁。
按照达尔文(1842)理论, 珊瑚礁历经裙礁、堡礁和环礁不同演化阶段, 环礁是其进化到最高阶段的产物。因此, 现阶段的西沙群岛八大环礁, 代表了珊瑚-珊瑚礁最高演化阶段的产物, 是研究珊瑚-珊瑚礁最好的天然实验室。
1.2 珊瑚的基本类型以珊瑚虫触手或腔肠数目的6或6的倍数、8或8的倍数, 亦即6只— 8只触手、6个— 8个腔肠为基准, 区分珊瑚为八放珊瑚(Octocorallia)和六放珊瑚(Hexacorallia)2个基本类型。
八放珊瑚: 最早发现于5.6× 108年的前寒武纪(图 1)。它们大都是软珊瑚, 难以形成规模钙质骨骼, 属于附礁生物群落(何心一, 1964; 戴昌凤等, 2013)。八放珊瑚最早发现于1758年, 至今提出了2个分类方案(Hickson, 1931; Bayer, 1981)。发现八放珊瑚赋存繁茂于相对较深的浅海水体(戴昌凤等, 2013)。
六放珊瑚: 最早发现于2.2× 108年前的中三叠世(齐文同, 2007), 而不是Wells(1956)描述的晚三叠世; 这一发现为全球研究者推崇(Qi, 1984; 齐文同, 1989), 展示了中国在珊瑚研究的学术地位, 这得益于乐森璕院士、王鸿祯院士和杨遵仪院士的贡献。中三叠世安尼期 Faviina亚目珊瑚分子的研究论文 1983年在华盛顿第三届国际刺细胞会议宣读, 入选文集(Qi, 1984); 在广西、云南南部、贵州东部野外的发现(邓占球, 2006), 进一步推进了珊瑚起源于中三叠世并得到国际公认(齐文同和斯坦利, 1989)。这是中三叠世印支运动全球大规模海侵、动物门类发生广泛演化进化的结果。2014年, 作者等曾在扬子区四川、浙江和安徽野外发现珊瑚化石露头, 它们富集赋存于那里, 直立于地表的百余米露头剖面之中(许红等, 2016); 但受工作任务限制(王修齐等, 2016; 许红等, 2016), 遗憾至今未采集样品。在广西、云南、贵州等地三叠纪、侏罗纪珊瑚礁的野外发现工作, 开始研究的时间可能更早(邓占球和孔磊, 1984)。
1.3 珊瑚形成珊瑚礁的造礁作用与附礁作用珊瑚分单体和复体, 群居单体珊瑚活体几厘米、几十厘米, 即“ 珊瑚虫” (图 1)。珊瑚虫原地生长, 圆筒状, 以浮游生物为食, 口入同时残骸口出, 由此原地覆盖固着成长扩大。单体珊瑚有环绕状体壁, 表面饰粗细不等皱纹, 进化程度较高的珊瑚体壁退化; 复体珊瑚千姿百态, 有团块状、纤细丛状、枝状、指状、丛状、扇形、鞭形、表覆形等; 有从母体轴部或侧方分芽而出者, 通过体壁上的孔或管, 或各单体之间共骨组织、空位管联结贯通; 珊瑚成长过程中与虫黄藻共生, 吸收后者光合作用合成的碳水化合物、氧、钙或二氧化碳(梁景芬和曾昭璇, 1989; 戴昌凤等, 2013), 形成纤细方解石质或文石质羽针、羽簇或羽榍碳酸钙, 不断加厚强壮长高, 形成珊瑚礁。因此, 所谓珊瑚礁, 是由珊瑚形成的足以抵御最强烈风浪、凸起于周缘的化石长垣, 被誉为会长高的石头; 珊瑚礁可绵延数十至数百千米。
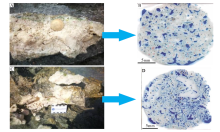
迄今为止, 全球已经发现六放珊瑚和八放珊瑚超过1000种, 估计多达6000种(梁景芬和曾昭璇, 1989), 其中四放珊瑚已经灭绝。珊瑚多数属于刺丝胞动物门珊瑚虫纲, 是很高级的腔肠动物; 造礁珊瑚可以形成碳酸钙骨骼或骨针, 属于六放珊瑚的石珊瑚和黑珊瑚2个目, 少数属于水螅虫纲, 水螅虫纲的水螅珊瑚也是常见的造礁珊瑚, 它们形成砾状珊瑚灰岩(图 2); 八放珊瑚既有软珊瑚也有石珊瑚, 少数种属于石珊瑚的笙珊瑚和蓝珊瑚。附礁珊瑚与其的区别, 是在它们死亡之后, 或形成遗存微小化石碎屑的堆积物灰沙, 八放珊瑚的软珊瑚、柳珊瑚隶属于此类; 还包括其他造礁生物形成的动物屑如软体动物壳、植物屑, 但这些碎屑都太过微小, 被归于附礁生物大类。
因此, 所谓造礁, 是珊瑚等主要造礁生物光合作用、钙化作用、化石化作用、沉积堆积作用和压实胶结成岩作用的结果; 所谓附礁, 是软珊瑚及其他动植物屑归附礁区固结沉积物, 呈松散堆积胶结成岩作用的结果。
1.4 珊瑚生态系统生物多样性特征2001年, Spalding 等按照“ 最高珊瑚覆盖率礁坪、礁顶、浅水礁区等近表层礁” 标准计算全球珊瑚礁总面积, 约为28.43万km2。珊瑚礁生态系统富集海洋生物达数万种, 不但生物多样性特征十分明显(Moore and Best, 2001; 赵美霞等, 2006), 而且物种多样性和高物种多样性首屈一指。事实上, 全球珊瑚礁生态系统只占全部海洋生态系统面积0.2%(David and James, 2000); 但却保有全球生态系统每100 km2最高平均物种数, 因此, 是物种多样性最高的生态系统。通过生态系统物种比较研究, Reaka-Kudla(1997)发现:
a. 全球生态系统生物平均物种数0.36;
b. 全球陆地生态系统生物平均物种数0.85;
c. 全球海洋生态系统生物平均物种数0.08;
d. 全球热带雨林生态系统生物平均物种数11.0;
e. 全球珊瑚礁生态系统生物平均物种数15.5。
由此可见, 珊瑚-珊瑚礁生态系统是超级热带海洋雨林, 是海洋生物活动高度集中的代表, 维持生物多样性和物种多样性最关键最核心生态要素的珊瑚是其主角, 地位不可替代。
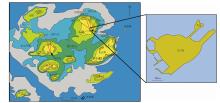
2 南海中央区珊瑚-珊瑚礁南海面积300万km2, 是西太平洋边缘海唯一发育东沙、西沙、中沙和南沙4大群岛珊瑚-珊瑚礁的边缘海, 也包括南海周缘珊瑚-珊瑚礁, 笔者将它们划分为南海周缘区珊瑚-珊瑚礁和南海中央区珊瑚-珊瑚礁。
1)南海周缘区珊瑚-珊瑚礁: 既包括中国大陆沿岸如深圳湾、涠洲岛、香港岛、海南岛和台湾岛近海珊瑚-珊瑚礁, 也包括南海周缘其他各国如印度尼西亚、菲律宾、马来西亚的珊瑚-珊瑚礁。
2)南海中央区珊瑚-珊瑚礁(图 3): 即南海四大群岛珊瑚-珊瑚礁, 这里赋存分布280多个岛、礁、滩、沙, 绝大多数属于深海台地型珊瑚-珊瑚礁。
公元2世纪的汉代, 中国先民们发现了南海诸岛星罗棋布的珊瑚-珊瑚礁, 今天知道它们全部属于碳酸盐岩台地型珊瑚礁, 位于深水。其中, 东沙群岛位于南海北部大陆坡折海域, 周缘海水最浅, 仅350 m; 西沙群岛位于南海西北部大陆坡海域, 周缘水深超过1200 m; 中沙— 南沙群岛周缘海域水深超过2000 m; 它们孤悬于南海, 陆地面积奇小。
南海区发现石珊瑚571种(Huang et al., 2014), 只有部分具有专业鉴定描述, 最为经典者当属公开报道的东沙群岛285种(戴昌凤等, 2013), 西沙群岛的173种(黄晖, 2018); 以及在香港、海南岛等地发现的石珊瑚和各地软珊瑚。目前中国海域总计发现珊瑚726种(刘瑞玉, 2008; 戴昌凤等, 2013; 戴昌凤和秦启翔, 2017; 黄晖, 2018), 表现出高物种多样性的显著特征。其中, 南海中央区亦即中国珊瑚-珊瑚礁区280余个岛、礁、滩、沙及全部礁盘的珊瑚-珊瑚礁面积38 065 km2(赵焕庭, 1999; 戴昌凤等, 2013)。珊瑚礁面积大于地球总面积1%的国家21个(Wilkinson, 2004), 中国排名第8位, 位列印度尼西亚、澳大利亚、菲律宾、法国、巴布亚新几内亚、斐济和马尔代夫之后, 因此, 中国是世界上最为重要的主要珊瑚-珊瑚礁国家。
值得进一步强调的是, 南海中央区280余个岛、礁、滩、沙与水下沉积盆地赋存分布珊瑚-珊瑚礁, 二者总面积可达17.26× 104 km2(许红等, 2015)①(① 许红,等.2015.大型油气田重大科技专项:南海新生代生物礁储层特征对比研究成果报告.); 它们都已有2千万年发育史(许红和王玉净, 1999)。其中, 中国在珠江口盆地发现了8个新近纪生物礁油气田(陈斯忠和胡平忠, 1987; 许红, 1992), 迄今南海发现的最大油田和气田储集层都是生物礁; 通过南海沉积盆地生物礁和最新西沙群岛井— 震资料联合解释, 确认南海中央区珊瑚-珊瑚礁基底的确是火山岩, 地质年代为侏罗纪, 同时发现古元古代(18~24)× 108年分子(Xu et al., 2018)。
南海中央区4大群岛出露海面珊瑚-珊瑚礁分为珊瑚岛、珊瑚礁、珊瑚滩和珊瑚沙4种类型, 其中绝大多数属于珊瑚礁, 个别属于火山岛, 如西沙群岛东岛环礁的高尖石岛。下面重点介绍西沙群岛珊瑚-珊瑚礁和东沙群岛珊瑚-珊瑚礁的生物多样性, 简单介绍南沙和中沙的基本情况。
2.1 西沙群岛珊瑚-珊瑚礁2.1.1 概况
中国西沙群岛珊瑚-珊瑚礁早期科考始于建国前, 规模科考始于20世纪70年代, 总计钻科探井7口, 总进尺约4800 m。2014年5月, 科探井西科1井历时2年3个月钻穿西沙群岛生物礁, 实现了200年前达尔文钻穿太平洋环礁的梦想, 揭露生物礁1257.52 m, 与西永1井1251 m差6.52 m; 其井底火山岩之上第1块岩心出现的造礁生物是珊瑚藻(许红等, 2015①(① 许红,等.2015.大型油气田重大科技专项:南海新生代生物礁储层特征对比研究成果报告.), 2017②(②许红.2017.第六届全国沉积学大会A1分会场重点报告:碳酸盐(岩)沉积学基础理论与进展.)), 下伏古老辉长岩闪长岩花岗片麻岩(Xu et al., 2018)。西沙群岛历经2千万年发育史(许红和王玉净, 1999), 是南海四大群岛自然成因陆域面积最多的群岛, 属于退积型珊瑚礁, 但它所经历退积成礁作用与澳大利亚大堡礁3.2万年以来的5期退积作用(Jody et al., 2018)不同。西沙群岛形成深水台地型珊瑚藻礁、仙掌藻礁和珊瑚礁, 期间先后经历新近纪晚期多期全球性冰川型海退事件, 曾经记录的海退规模为135.51 m(Xu et al., 1999), 最新数据超过了160 m(许红, 2017)②(②许红.2017.第六届全国沉积学大会A1分会场重点报告:碳酸盐(岩)沉积学基础理论与进展.)。此外, 西沙群岛位于大陆坡, 属于典型孤立碳酸盐岩台地深水环境, 距它最近的大陆海南岛330 km, 远于大堡礁与澳大利亚海岸的距离。
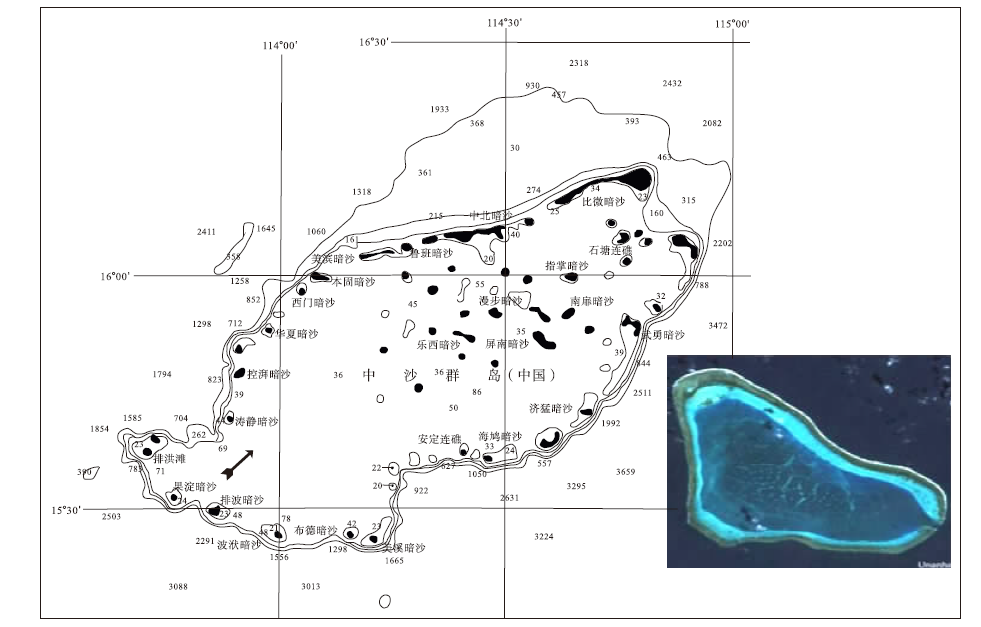
西沙群岛由北向南、由西向东依次形成的8大环礁分别是: 北礁, 永乐环礁, 华光礁, 盘石礁, 玉琢礁, 宣德环礁, 东岛环礁, 浪花礁。不难发现, 真正的环形珊瑚礁只有永乐环礁, 宣德环礁也算1个, 但它呈马蹄形(图 4); 二者周缘都发育10余个岛、礁、滩、沙。西沙群岛东北为西渡滩, 东南浪花礁, 正南嵩焘滩, 西南中建岛, 西北北礁; 域内发育唯一一座成因于早更新世的火山岩小岛— 高尖石岛, 位于东岛环礁。西沙群岛出露海面的岛屿在自然形成的陆地面积最大, 宜居环礁也最多, 含台地面积1836.4 km2, 礁坪面积223.6 km2, 礁湖面积1612.8 km2; 出露海面的岛、礁、滩、沙计41座, 自然干出的陆地总面积9.703 km2; 组成复合环礁3个、单体环礁5个、台礁1个和暗滩2个。
在西沙群岛, 天文大潮礁盘裸露, 2009年科考航次的调查记录了那里绵延数十千米珊瑚群落分布的模样, 那种多姿多彩、千姿百态和繁茂的景观, 颠覆了随行研究生的专业选择。此外, 也在西沙群岛多口科探井观察到上新世、中新世化石珊瑚(许红和王玉净, 1999; 周小康等, 2018); 在西沙群岛不同岛礁潮间带, 发现多种类型海滩岩(孙志鹏等, 2010), 其中富集大量混杂堆积的鹿角珊瑚化石。
2.1.2 生物多样性特征
2009年中国实施了建国以来最大规模西沙群岛珊瑚礁科考, 涉及12条科考与保障舰船, 包括浅潜、摄像、差分GPS与动植物学、沉积学和旅游学专业, 登陆27个岛、礁、滩、沙(西沙工委暨中南西沙办事处, 2009)③(③ 西沙工委暨中南西沙办事处.2009.西沙群岛多学科联合科考报告.), 发现永乐环礁晋卿岛礁盘绵延数十千米(图 5), 形成可抵御最强烈风浪、波基面以下向海一侧低能透光带发育的抗浪性珊瑚高生物多样性和高物种多样性珊瑚礁长垣, 天文大潮期间, 那里珊瑚重见天日, 那种美丽令人震撼并流连忘返; 当然, 那里也存在海底潜流冲沟与死亡珊瑚(图 5-F)。
2012年, 许红等发现石岛浅水潟湖区赋存富集活体仙掌藻(Xu et al., 2015a), 这一发现引发与晚新近纪钻井化石仙掌藻对比的研究(Xu et al., 2015b; 许红, 2017)④(④许红.2017.第六届全国沉积学大会A1分会场重点报告:碳酸盐(岩) 沉积学基础理论与进展.), 夯实千万年以来西沙群岛绿藻延续至今的藻礁造岩作用; 2019年, 在“ 永乐龙洞” 发现采集活体仙掌藻(马晓等, 2021), 实证钙藻的造礁作用对于西沙群岛多个环礁产生实际贡献。
2016年, 赵美霞等发现西沙群岛特殊海绵生物群落(赵美霞和余克服, 2016)。2018年, 黄晖报道了汇集十年调查的系列发现, 涉及西沙群岛236种珊瑚, 173种石珊瑚, 27种海绵, 16种红绿藻, 309种鱼类; 同年, 汪品先⑤(⑤汪品先.2018.央视新闻、综合频道及新华视点等有关中国科学院深海科学与工程研究所“探索一号” 科考船“深海勇士” 号西沙海 域下潜报道及视频.)深潜发现西沙北礁深水柳珊瑚; 2018年底, 在西沙石岛海域发现自然恢复8种珊瑚(许红等, 2019)⑥(⑥许红,等.2019.中国环境科学学会2019年科学技术年会第八分会场口头报告:“中国西沙人居环礁珊瑚-珊瑚礁生态环境现状科考”. 西安.)。2019年3-4月份, 许红率队实现西沙群岛宣德环礁、永乐环礁和东岛环礁多学科综合科考, 发现六放珊瑚46种, 八放珊瑚6种; 鱼类40余种, 海龟两类(图 6, 表 1); 其中有45种石珊瑚是国家二级重点保护动物(占了南海石珊瑚物种的近四分之一)。该项发现时间很短, 而且是随机的, 没有刻意回避及选择, 半月内每天都能够拍到, 并且最后总数如此之多的珊瑚及其生态系统中的其他生物, 说明西沙群岛珊瑚-珊瑚礁生物多样性极高。
表 1
Table1
表 1(Table1)
 表 1 2019西沙群岛科考航次发现珊瑚属种一览表 Table1 List of coral species found in Xisha Islands during the 2019 expedition| 序号 | 珊瑚科名 | 珊瑚种名 | 种数 |
|---|
| 六放珊瑚 | 杯形珊瑚科 | 多曲杯形珊瑚Pocillopora meandrina; 埃氏杯形珊瑚 Pocillopora eydouxi; 疣状杯形珊瑚Pocillopor averrucosa | 3种 | | 2 | 滨珊瑚科 | 滨珊瑚 Porites sp.; 澄黄滨珊瑚Porites lutea; 角孔珊瑚 Goniopora | 3种 | | 3 | 蜂巢珊瑚科 | 刺星珊瑚Cyphastrea.; 锯齿刺星珊瑚 Cyphastrea serailia; 蜂巢珊瑚 Favia; 宝石刺孔珊瑚 Echinopora gemmacea; 角蜂巢珊瑚 Favites; 标准蜂巢珊瑚 Favia speciosa; 薄片刺孔珊瑚 Echinopora lamellosa; 同双星珊瑚 Diploastrea heliopora; 扁脑珊瑚Platygyra; 交错扁脑珊瑚Platygyra crosslandi; 弗尼吉亚肠珊瑚 Leptoria phrygia | 11种 | | 4 | 菌珊瑚科 | 西沙珊瑚 Coeloseris mayeri; 牡丹珊瑚 Pavona; 小牡丹珊瑚Pavona minuta; 皱纹厚丝珊瑚Pachyseris rugosa | 4种 | | 5 | 鹿角珊瑚科 | 壮实鹿角珊瑚Acropora robusta; 简单鹿角珊瑚Acropora austera; 鼻形鹿角珊瑚Acropora nasuta; 栅列鹿角珊瑚Acropora palifera; 风信子鹿角珊瑚 Acropora hyacinthus; 中间鹿角珊瑚Acropora intermedia; 巨锥鹿角珊瑚Acropora monticulosa; 美丽鹿角珊瑚Acropora formosa; 强壮鹿角珊瑚Acropora valid; 鹿角珊瑚 Acropora sp.; 多孔鹿角珊瑚Acropora millepora; 芽枝鹿角珊瑚 Acropora gemmifera; 多星孔珊瑚Astreopora myriophthalma; 繁锦蔷薇珊瑚Montipora efflorescens; 蔷薇珊瑚Montipora sp. | 15种 | | 6 | 裸肋珊瑚科 | 阔裸肋珊瑚Merulina ampliata | 1种 | | 7 | 木珊瑚科 | 盾形陀螺珊瑚Turbinaria peltata; 皱折陀螺珊瑚Turbinaria mesenterina | 2种 | | 8 | 枇杷珊瑚科 | 丛生盔形珊瑚Galaxea fascicularis; 盔形珊瑚 Galaxea fascicularis | 2种 | | 9 | 石芝珊瑚科 | 石芝珊瑚Fungia sp.; 绕石珊瑚 Herpolitha limax; 楯形石芝珊瑚Fungia scutaria | 3种 | | 10 | 铁星珊瑚科 | 毗邻沙珊瑚Psammocora contigua | 1种 | | 11 | 褶叶珊瑚科 | 菌状合叶珊瑚Symphyllia agaricia | 1种 | | 八放珊瑚 | | 多孔螅 Milleporina; 苍珊瑚Heliopora; 笙珊瑚Tubipora; 肉芝软珊瑚Sarcophyton; 短指软珊瑚Sinularia; 海葵 Heteractis | 6种 |
注: 本表含46种造礁石珊瑚, 其中, 45种是国家2类重点保护动物, 占南海有记录石珊瑚物种近四分之一。 | 表 1 2019西沙群岛科考航次发现珊瑚属种一览表 Table1 List of coral species found in Xisha Islands during the 2019 expedition |
2.1.3 讨论
将2019年调查所见(图 6)与前期研究成果对比, 发现珊瑚个体体积、群落繁茂程度等均不如2009年科考航次在晋卿岛礁盘的发现(图 5), 也不如黄晖等更早前在西沙群岛多岛的调查发现(图 7)。
这是一个全球性现象。2019年, 自然杂志报道河流输入对于澳大利亚大堡礁珊瑚-珊瑚礁白化作用产生影响, 该认识来自一个加拿大团队调查的结论, 指出其或是珊瑚白化的主要影响因素(Tollefson, 2019)。与澳大利亚大堡礁大陆架浅水比较, 西沙群岛深蓝海水能见度更好, 可达30 m, 大陆坡深水生态环境条件当然更胜一筹。不能说西沙群岛目前的现象是因河流影响珊瑚-珊瑚礁环境, 但有一定的启示意义, 可以引起我们对环保意识与生态治理的足够重视。保护好西沙珊瑚-珊瑚礁及其他区域的珊瑚-珊瑚礁, 海洋科技工作者责无旁贷。
2.2 东沙群岛珊瑚-珊瑚礁2.2.1 概况
东沙群岛像指环, 发育一个环礁及其干出的海岛— 东沙岛(图 8), 含神狐暗沙、一统暗沙、周缘的北卫滩和南卫滩等岛礁滩沙。东沙群岛距离广东陆丰— 惠来海岸最近, 为240 km; 距离万山群岛担杆岛280 km, 距离香港、深圳超过300 km, 距离高雄450 km。比较南海其他3个群岛, 东沙群岛珊瑚礁沉积厚度最小, 推测仅为350 m, 面积也小, 约为500 km2。
2.2.2 生物多样性特征
1995年, 中国学者实施东沙群岛珊瑚-珊瑚礁系统调查研究, 迄今已经发现记录了573种珊瑚, 详细描述了285种六放珊瑚, 288种八放珊瑚(戴昌凤等, 2013; 戴昌凤和秦启翔, 2017)。涉及六放珊瑚亚纲2目14科。石珊瑚目13科: 鹿角珊瑚科, 轴孔珊瑚科, 微孔珊瑚科, 莲珊瑚科, 亶珊瑚科, 刺叶珊瑚科, 真叶珊瑚科, 菊珊瑚科, 圆星珊瑚科, 贝壳珊瑚科, 树珊瑚科; 黑角珊瑚目鞭角珊瑚科。涉及八放珊瑚亚纲3目8科。匍根珊瑚目2科: 笙珊瑚科和羽珊瑚科; 蓝珊瑚目蓝珊瑚科; 软珊瑚目5科: 软珊瑚科, 穗珊瑚科, 柳珊瑚科, 扇珊瑚科和鞭珊瑚科。此外, 还有水螅虫纲千孔珊瑚目千孔珊瑚科和柱星珊瑚目的柱星珊瑚科等(戴昌凤等, 2013)。
从上面的成果可以看出, 台地型大陆坡珊瑚礁环境诠释的珊瑚高物种多样性特征, 同样构成珊瑚生态系统奇迹(图 9)。
详尽的调查科考证实, 东沙群岛珊瑚礁生态系统的发育和珊瑚属种分布具有规律性, 在环礁外缘, 北部、东部赋存软珊瑚亦即发现更多八放珊瑚, 却与水深有关, 5 m水深主要以石珊瑚占优势, 10~25 m乃至30 m以软珊瑚占优势; 西部多发育石珊瑚亦即六放珊瑚, 它们从5 m水深开始直到25 m都是优势种; 在南部, 5 m和25 m以石珊瑚为主, 10~20 m却大量发现软珊瑚; 在潟湖区, 发现较多六放石珊瑚; 在航道位置, 则是软珊瑚居多(戴昌凤等, 2013; 戴昌凤和秦启翔, 2017)。由此可见, 今天已经具有近6× 108年历史的八放珊瑚群落仍然还是能够很好适应环境能力的珊瑚物种, 生命力顽强, 继续诠释着它们统治珊瑚生态系统的能力。
2.3 中沙群岛珊瑚-珊瑚礁中沙群岛是南海4大群岛位置居中靠北的群岛(图 3), 古称红毛浅; 西距西沙群岛永兴岛200 km。在中沙大环礁及耸立于深海盆之上的环礁礁盘发育26个暗沙: 比微暗沙、中北暗沙、鲁班暗沙、美滨暗沙、本固暗沙、西门暗沙、华夏暗沙、控湃暗沙、涛静暗沙、排洪滩、果淀暗沙、排波暗沙、波洑暗沙、布德暗沙、美溪暗沙、安定连礁、海鸠暗沙、济猛暗沙、武勇暗沙、隐矶滩、石塘连礁、指掌暗沙、南扉暗沙、漫步暗沙、屏南暗沙、乐西暗沙; 加上周缘的宪法暗沙、中南暗沙和黄岩岛(图 10)。其中, 中沙大环礁是南海2大巨型环礁之一, 长约 140 km, 宽约60 km, 面积6900 km2; 距海面约10~26 m, 故多隐没于海面之下, 礁坪的1/6在天文大潮最低潮干出时, 可见珊瑚、珊瑚礁规模分布。所以, 中沙大环礁珊瑚大量发育, 生物量很高(图 11), 大量珍贵石珊瑚形成五光十色的海底花园; 附近海域则是南海重要渔场, 盛产金带梅鲷、旗鱼、箭鱼、金枪鱼等。
2.4 南沙群岛珊瑚-珊瑚礁在南沙群岛, 钻井揭露了2000余米岩心(张偲, 2018)⑦(⑦张偲.2018.在第十三届全国生物多样性科学与保护研讨会上的大会报告.内蒙古呼和浩特.), 井底见花岗片麻岩和玄武岩。
南沙群岛面积巨大, 辖239个岛礁滩沙, 其中自然岛屿11个, 5个沙洲, 20个干出水面的礁; 距祖国大陆约1000 km的礼乐滩大环礁是南沙也是南海的最大的巨型环礁。面积约为台湾岛的四分之一, 达7000 km2; 一般水深18 m, 最深40 m, 最浅9 m, 天文大潮时干出水面; 造礁珊瑚星罗棋布(图 2, 图 12)。钻探发现礼乐大环礁古近系和新近系巨厚, 沉积一套砂泥岩+碳酸盐岩地层, 厚达5000 m, 形成礼乐盆地。它被评价为南沙石油远景区诸盆地之一, 基底为白垩纪火成岩及更老变质岩, 位于中国南海九段线内。目前, 中国有效控制南沙群岛11座岛礁: 永暑礁、美济礁、渚碧礁、赤瓜礁、南薰礁、 东门(垦南)礁、华阳礁、 五方礁、仁爱礁、 信义礁、仙娥礁。
4 结论1)划分中国海洋的珊瑚-珊瑚礁为南海中央区珊瑚-珊瑚礁和南海周缘区珊瑚-珊瑚礁, 指出中国是全球排位第8的主要珊瑚-珊瑚礁国家, 地位举足轻重; 南海中央区珊瑚-珊瑚礁即南海4大群岛珊瑚-珊瑚礁, 280余个岛、礁、滩、沙在基底火山岩上历经20 Ma发育, 与沉积盆地生物礁一起组成南海珊瑚-珊瑚礁; 简介了西沙、东沙、中沙和南沙群岛美丽的珊瑚-珊瑚礁属种。
2)中国海域调查发现记录了近600种造礁、附礁珊瑚, 系统汇集描述了10余年来南海中央区珊瑚-珊瑚礁不同科考航次调查发现于浅水礁盘、至25 m水深浮潜、浅潜、深潜和来自四大群岛不同岛礁的珊瑚调查航次发现与水下拍摄照片, 最新科学定名发现46种六放石珊瑚, 6种八放软珊瑚。
3)对比研究珊瑚生态系统生物多样性、物种多样性统计学认识, 超过热带雨林生态系统, 是地球上最为庞大的生态系统, 是珊瑚生态系统构建海洋金山银山的最大优势; 指出珊瑚-珊瑚礁对于践行生态治国理念举足轻重的地位; 珊瑚-珊瑚礁造岛、固礁、护鱼、防护岛岸流失, 属于无可替代、最大的海洋生态资源, 是珍贵的南海海洋国土。
4)南海珊瑚-珊瑚礁形成单体环礁和复合环礁, 对比分析了二者沉积学特征, 指出永乐环礁是南海唯一1个真正切合达尔文模式环礁, 是环礁发展到最高阶段产物, 构成现代海洋珊瑚-珊瑚礁形成演化研究最好的天然实验室与人居环礁, 珍视保护它们我们责无旁贷。
致谢 国家自然科学基金 41872114、科技部基础科技调查专项2017FY201407课题、大型油气田和煤成气开发重大科技专项2011ZX05025-002-04联合资助。






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 , 史国宁
, 史国宁