



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国贵州中生代四足类足迹化石的回顾与新发现*
[邢立达1, 2  , 曾嵘
, 曾嵘3 , 张羽1 , 邱亮1 , 佟柏霖1 , 王董浩1 ]
, 曾嵘|
|

第一作者简介 邢立达,男,1982年生,博士,中国地质大学(北京)地球科学与资源学院副教授,主要从事古脊椎动物学的教学与科研工作。E-mail: xinglida@cugb.edu.cn。
贵州省拥有丰富的中生代脊椎动物化石记录,多为三叠纪海生爬行类化石,侏罗纪和白垩纪的脊椎动物化石较少。目前,贵州省已发现中生代爬行类足迹点 9处,分别为贞丰牛场、贞丰龙场、安顺幺铺、赫章辅处、毕节响水( 2处)、仁怀茅台、赤水宝源和习水同民足迹点。文中除描述了贞丰龙场新增的一批足迹化石外,整理并总结了所有足迹点的发现史、研究现状、足迹类型与特征以及所反映的古生态信息。已发现的遗迹记录由 69条行迹和 420个孤立足迹组成,足迹总数达 991个。所有的遗迹化石指示了至少 477个造迹者,包括海生爬行类、手兽足迹类、非鸟兽脚类、鸟类、蜥脚类和鸟臀类等,其中三叠纪以海生爬行类主导,侏罗纪和白垩纪以蜥臀类主导。这个数据集为贵州中生代古生物类群的多样性提供了重要的信息,是晚中生代稀缺骨骼化石记录的良好补充,为研究贵州省中生代脊椎动物群的古生态与古环境提供了更多的证据。
About the first author Xing Li-Da,born in 1982,is an associate professor in School of Earth Sciences and Resources,China University of Geosciences(Beijing). He is mainly engaged in teaching and scientific research of paleovertebrates. E-mail: xinglida@cugb.edu.cn.
Guizhou Province is rich in records of Mesozoic vertebrate fossils,mostly Triassic marine reptile fossils,while Jurassic and Cretaceous vertebrate fossils are scarce. There are nine Mesozoic tetrapod track sites discovered in Guizhou Province,which are Niuchang and Longchang sites from Zhenfeng,Yaopu site from Anshun,Fuchu site from Hezhang,Xiangshui sites I and II from Bijie,Maotai site from Renhuai,Baoyuan site from Chishui and Tongmin site from Xishui. This paper described more tracks of Longchang site from Zenfeng,and try to summarize the discovery and research history,track types and their characteristics,and the palaeoecological information reflected in all nine track sites. All these records consist of 69 trackways and 420 isolated tracks for a total of 991 tracks of tetrapod remains indicative of at least 477 trackmakers,including marine reptiles,chirotheriids,non-avian theropods,birds,sauropods,and ornithischian dinosaurs. Among them,the Triassic track records are dominated by marine reptiles,while the Jurassic and Cretaceous records show the dominance of saurischian. This dataset is an important supplement to the scarce bone fossils of the Late Mesozoic that contributed to the diversity of Mesozoic paleontological taxa in Guizhou Province,and provides more evidence for understanding the palaeoecology and palaeoenvironment of the Mesozoic vertebrate fauna of Guizhou Province.
开放科学(资源服务)标识码(OSID) 
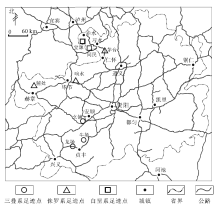
贵州省地处云贵高原东部, 地质历史悠久, 沉积地层较为完整, 已发现大量古生物化石, 以埃迪卡拉纪瓮安群(Yin et al., 2017)、中寒武世凯里群(Zhao et al., 2005)、晚三叠世关岭群(汪啸风等, 2003)等多个生物群最为著名。近十余年来, 随着地质与古生物工作者对脊椎动物, 尤其是四足类足迹化石认知的加深, 贵州省接连发现了多处中生代四足类骨骼化石点与足迹点(图 1), 并以侏罗纪与白垩纪的记录为主。骨骼化石点包括大方、毕节、息烽和平坝等地(曾嵘, 2020), 足迹点包括三叠系的贞丰牛场等3处, 侏罗系的赫章辅处等4处, 以及白垩系的赤水宝源等2处。这些发现在很大程度上补充了该地区中生代四足类动物群的多样性, 在一些区域足迹化石甚至成为了解该地区古动物群的唯一途径。此外, 四足类足迹化石还提供了动物足部形态、生活习性以及运动方式等多方面的信息, 对研究贵州省中生代的古生态、古地理等都具有重要的意义。
 | 图 1 贵州省中生代四足类足迹化石点分布Fig.1 Distribution of the Mesozoic tetrapod track sites in Guizhou Province |
贵州省构造位置属扬子地台及其东南大陆边缘, 地层自元古宙至第四系均有出露, 以古生界和中生界三叠系最为发育, 同时古生物化石也十分丰富。文中所描述的足迹化石均来自中生代。
贵州省三叠系以海相沉积为主, 既有台地相浅水碳酸盐岩, 又有深水广海盆地陆源碎屑岩(贵州省地矿局, 1987)。发现脊椎动物化石的层位主要是中三叠统下部的关岭组。关岭组岩性组合为石灰岩、白云岩夹少量泥岩, 厚440~810 m。根据岩性组合特征, 关岭组可分为松子坎段(下段)和狮子山段(上段), 贞丰龙场、牛场和安顺幺铺足迹点都发现于松子坎段(吕洪波等, 2004; Xing et al., 2013)。松子坎段厚130~240 m, 主要岩性为白云岩与泥岩:底部为厚0.1~3 m的黄绿色、灰绿色、灰白色斑脱岩化玻屑凝灰岩(即“ 绿豆岩”); 下部以灰色中厚层白云岩为主, 夹泥质白云岩和角砾状白云岩等; 中上部为紫红色、灰绿色、褐黄色粉砂质泥岩、白云质泥岩与泥质白云岩以及泥质灰岩互层(吕洪波等, 2004)。
贵州省侏罗系地层发育不如三叠系, 只零星分布于贵州北西部的道真、遵义、贵阳、贞丰一线, 东部的天柱附近亦有零星出露, 以赤水至习水一带发育最好。侏罗系主要为陆相红色岩系, 以河流相及湖相最为发育, 自下而上可分为自流井组、沙溪庙组、遂宁组和蓬莱镇组, 其中自流井组发育最为齐全, 为一套滨湖及浅湖沉积(贵州省地矿局, 1987; 张晓诗等, 2016)。毕节响水大转弯足迹点和仁怀茅台岩滩足迹点都发现于自流井组马鞍山段(张晓诗等, 2016; Xing et al., 2019a, 2019b), 该段为一套灰白色、铁红色薄—中厚层细粒石英砂岩、长石石英砂岩与褐红色薄层泥质粉砂岩或粉砂质泥岩、泥质互层, 局部夹褐色薄—中厚层砾岩。赫章辅处足迹点发现于下侏罗统龙头山群, 岩性为紫红色、黄绿色泥岩、页岩, 夹浅灰色、紫褐色、灰绿色中厚层至厚层砂岩、长石石英砂岩、钙质粉砂岩, 大致与四川盆地下侏罗统珍珠冲组或川南香溪群上部相当(Xing et al., 2021)。
贵州省白垩系仅零星出露, 面积小, 主要分布于水城、晴隆、贞丰一线北东广大地区, 缺失早白垩世早、中期沉积。其中, 赤水、习水地区的白垩系与四川盆地毗连, 属大型内陆拗陷盆地边缘河流沉积(邢立达等, 2016; 曾嵘, 2020)。目前, 仅在四川盆地东南缘的贵州赤水宝源和习水同民发现有脊椎动物足迹化石记录(Xing et al., 2011)。赤水和习水地区的白垩系红层属白垩系中部夹关组(窝头山组), 为一套陆相沉积, 主要岩性为紫红色、砖红色、紫灰色厚层中细粒长石石英砂岩, 夹多层紫红色泥岩, 底部为砾岩层(四川省地质矿产局, 1997)。
迄今为止, 在贵州省已发现的恐龙足迹点共9处, 包括三叠系的贞丰牛场、贞丰龙场、安顺幺铺共3处, 侏罗系的赫章辅处、毕节响水(2处)、仁怀茅台共4处, 以及白垩系的赤水宝源和习水同民2处。对上述9处足迹点, 笔者整理和总结了它们的发现史、研究现状、足迹类型与特征及其所反映的古生态信息, 并分别予以介绍。
贞丰牛场足迹点位于贵州省黔西南布依族苗族自治州贞丰县北盘江镇(原牛场乡)青杠林村上坝组自然村, 足迹化石产出于中三叠统关岭组松子坎段泥裂构造发育的泥质白云岩层面上, 共发现有3条行迹, 计60个足迹(Xing et al., 2013)。目前暴露的含足迹化石的层面面积约100 m2, 3条行迹分别长10 m、9 m和2 m。其中行迹A保存最好, 后足迹五趾, 多数后足迹保存有对应的三趾型前足迹。另外2条行迹(B与C)稍浅, 多数未保存前足迹(图 2; 表 1)。
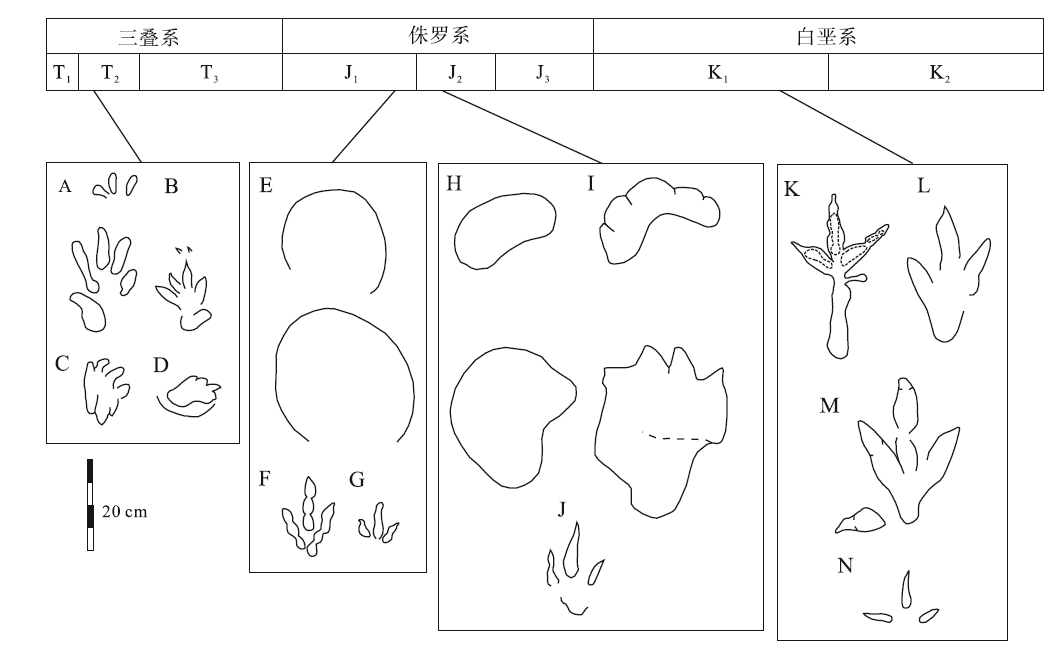 | 图 2 贵州省中生代四足类足迹化石形态简图 A—贞丰牛场的Chirotherium barthii; B—贞丰龙场的Chirotherium barthii; C—安顺幺铺的Anshunpes aquacursor; D—安顺幺铺的Dikoposichnus luopingensis; E—赫章辅处的Brontopodus-like; F—赫章辅处的Eubrontes-type; G—赫章辅处的Anomoepus-type; H—毕节响水的cf. Brontopodus; I—仁怀茅台的Liujianpus shunan; J—仁怀茅台的Grallator; K—赤水宝源的cf. Irenesauripus isp.; L—习水同民的Gigandipus chiappei; M—习水同民的Eubrontes-type; N—习水同民的Wupus isp.Fig.2 Morphological schematic diagram of the Mesozoic tetrapod tracks in Guizhou Province |
| 表 1 贵州省中生代四足类造迹者丰度统计 Table 1 Abundance statistics of dinosaur trackmakers from the Mesozoic of Guizhou Province |
20世纪60年代, 当地村民在建造粮食晒场时, 在地表发现了这些奇怪的爪印, 认为是传说中的神鬼所留(Xing et al., 2013)。1988年4月, 贵州省博物馆自然部王新金根据贞丰县城建局张英群提供的线索, 赴上坝村考察了这些足迹, 对此批脚印进行了翻模并在之后以模型展出过。1988年5月, 贵州省地质矿产局区域地质调查队的王雪华和马骥对这些足迹进行了考察, 并进行了简要的描述, 认为其应属于恐龙足迹(王雪华等, 1989)。之后, 吕洪波等(2004)描述了该批足迹, 认为其保存在近岸浅水环境的泥裂泥质白云岩层面上, 为Chirotherium(手兽足迹); 这是在中国首次记录的Chirotherium, 但作者没有给出具体遗迹种。Lockley和Matsukawa(2009)对比了贵州与欧洲各地发现的Chirotherium, Klein和Lucas(2010)在讨论全球三叠纪四足动物足迹的生物地层和地质年代意义时, 认为牛场的Chirotherium可归入C. barthii。
邢立达等于2011年考察并详细记录了牛场的Chirotherium, 从形态特征与系统分类等方面详细描述了这批足迹, 亦将其归入C. barthii(Xing et al., 2013)。鉴定基于以下特征: 长22~25 cm、中等大小的chirotheriids(手兽足迹类), 行迹窄, 步幅角143° ~164° , 复步长也相对较低; 前足迹比后足迹更强烈地向行迹中线外转, 分别为11° 与20° ; 后足迹的第Ⅰ ~Ⅳ 趾较短且细, 其中第Ⅱ 与第Ⅳ 趾对称, 近乎等长, 中间的第Ⅲ 趾最长, 第Ⅰ 趾较短较细且位于稍后的位置, 第Ⅰ ~Ⅳ 趾的近端趾垫形成后凹的边界; 后足迹的第Ⅴ 趾有1个大的椭圆形近端趾垫, 位于第Ⅳ 趾的中线上, 并且具有明显弯曲的、收窄的趾骨区; 前足迹的第Ⅲ 趾最长, 第Ⅱ 趾和第Ⅳ 趾较短且等长, 第Ⅰ 趾和第Ⅴ 趾短但极少保存, 第Ⅳ 趾和第Ⅴ 趾横向伸展(Xing et al., 2013)。
贞丰龙场足迹点位于黔西南布依族苗族自治州贞丰县龙场乡北石灰窑村新修的关(岭)兴(义)公路西侧, 与牛场足迹点相距十几千米, 层位与牛场一致, 为中三叠统关岭组松子坎段。龙场的足迹由至少3个造迹者所留, 但行迹较为凌乱, 保存较差。这批足迹由吕洪波等于2003年11月发现, 并于次年描述, 亦将其归入Chirotherium(吕洪波等, 2004)。Xing 等(2013)详细描述了这批足迹, 记录了至少7个后足迹和1个前足迹。从形态上, 龙场足迹与牛场足迹相似, 但略小, 后足迹长15 cm, 保存亦较差, 各个趾之间的隔离度较低, 其中保存较好的标本显示了第Ⅰ 趾有圆形的趾垫, 第V趾近端膨大。这批标本亦被归入C.barthii(Xing et al., 2013)(图 2; 表 1)。
安顺幺铺足迹点位于安顺市经济技术开发区么铺镇红龙村将军山, 距红龙村西南约542 m(曾嵘, 2020)。幺铺足迹点的分布范围长约26 m、宽约5.7 m。足迹化石产自中三叠统关岭组下段薄至中厚层泥质白云岩的层面上, 层面泥裂发育, 属于潟湖或浅海沉积。
2009年, 贵州省博物馆蔡回阳等在开展第三次全国文物普查工作期间, 在安顺市文物局工作人员的带领下考察发现了该足迹点。2017年7月, 贵州省博物馆与中国地质大学(北京)的学者对该足迹点进行详细调查, 记录了400多个海生爬行类足迹。2018年底, 贵州省博物馆、中国地质大学(北京)与德国古爬行及两栖动物博物馆的学者再次对足迹点进行详细考察。
安顺幺铺足迹点发现有22条行迹和200多个孤立的足迹以及一些不确定的痕迹(Xing et al., 2020)。所有的足迹都明显不同于陆生造迹者的足迹, 而是属于海生爬行类在水底留下的足迹。其中, 12条行迹和1个孤立的足迹被归入Dikoposichnus luopingensis(罗平双桨迹)(Xing et al., 2020)(图 2, 表 1)。D. luopingensis是由中国地质调查局成都地调中心的学者在云南省罗平县首次发现并命名的, 属于中生代海生爬行类——幻龙类的觅食迹, 是幻龙类用前肢在海床上同步移动造成的(Zhang et al., 2014)。幺铺足迹点发现的Dikoposichnus亦呈半月—新月形的槽状凹陷, 单个足迹最长轴长5~20 cm, 有3~4个侧向尖锐的爪印, 后缘有1圈向后推移的沉积物堆。幺铺Dikoposichnus基本上成对出现, 且相对于行进路线对称分布, 足迹与行迹的整体形态与云南产出的Dikoposichnus相同(Xing et al., 2020)。
层面上另外一类足迹的形态和行迹模式与Dikoposichnus明显不同, Xing 等(2020)将其命名为Anshunpes aquacursor(奔波安顺足迹)(图 2, 表 1)。Anshunpes为半跖行—跖行性的四趾型足迹, 足迹长约15 cm, 宽9 cm, 整体形态较长, 各趾粗壮, 其中第Ⅱ 趾和第Ⅲ 趾最长, 而第Ⅳ 趾和第Ⅴ 趾较短, 足迹后部通常有1个拉长的双叶状或三叶状的“ 足跟” 。行迹整体较宽, 为45.8 cm, 左右足迹对称分布, 全部向外转, 是典型的拥有水下游泳或海底行走习性的四足动物所留。Anshunpes的行迹规律表明其造迹者采用同步运动、划行或撑行(punting)的运动方式——即对称的左、右肢同时运动, 潜在的造迹者为龙龟类和楯齿龙类。这是迄今最早的关于楯齿龙或龙龟类动物游泳或“ 撑行” 运动的遗迹化石记录(Xing et al., 2020)。
层面上还有部分遗迹很难确定来源, 这些遗迹包括4种类型: 第1种呈短而深的平行沟槽, 类似Anshunpes, 但边缘较为锋利, 可能为鳍行迹; 第2种为零散分布的凹痕, 形态类似于鱼巢或鱼类觅食迹, 但不涉及任何足迹模式, 不能确定是否与前述2种足迹的造迹者存在关联; 第3种为“ 多节” 索状堆积结构, 可能是鱼类的粪便; 第4种形如沙漏, 代表一些钙质沉积物(Xing et al., 2020)。
赫章辅处冒沙井足迹点位于赫章县辅处彝族苗族乡南部的冒沙井, 在农舍附近约20 m2、坡度约30° 的斜坡上分布有恐龙足迹, 相邻区域也有零星发现。足迹包括31个兽脚类足迹、1个疑似鸟臀类足迹和9个蜥脚类足迹, 均保存于下侏罗统龙头山群的细砂岩层面上。
2018年5月, 贵州省博物馆、中国地质大学(武汉)和赫章县文物事业管理局组成的联合考察组对赫章县古基乡恐龙化石点进行勘察时, 根据群众提供的线索, 在距古基乡化石点以西50 km处的赫章县辅处发现了恐龙足迹化石。2018年12月, 贵州省博物馆邀请中国地质大学(北京)和自贡恐龙博物馆学者再次详细考察赫章恐龙足迹点。
31个兽脚类足迹形成4条行迹, 保存最好的1条行迹包括4个足迹, 平均长16.8 cm, 平均长宽比为1.5, 爪印尖锐, 有2个清晰的跖趾垫, 趾印式为x-3-3-4-x, 第Ⅱ 趾与第Ⅳ 趾之间的趾间角平均为53° , 平均步幅角为166° , 足迹长与单步长之比为1︰2.91。其他3条行迹的足迹平均长度分别为19.5 cm、15.1 cm和14.3 cm。这些足迹的形态可归入Eubrontes type。有1个孤立足迹比较特殊, 仅9 cm长, 长宽比1.0, 第Ⅱ 趾和第Ⅳ 趾之间的夹角更宽, 达79° , 形态上与鸟臀类足迹Anomoepus type非常相似。大部分蜥脚类足迹是孤立存在的, 只有2个足迹构成一单步: 前足迹长15 cm, 呈弱U形, 长宽比为0.7, 没有明显的爪印, 跖趾区成凹形; 后足迹为椭圆形, 平均长35.8 cm, 长宽比为1.4, 无明显的爪印, 跖趾区后缘平滑弯曲; 整体形态特征可归入Brontopodus type(Xing et al., 2021)(图 2, 表 1)。
毕节响水大转弯足迹点位于毕节市金海湖新区响水乡北西向直线距离7 km处的大转弯南东简易公路南西侧岩壁的砂岩层面上。可细分为相距300 m的2个足迹点, 都产于下侏罗统自流井组马鞍山段中厚岩屑石英砂岩层面上。
大转弯足迹点一号点于2016年由成都理工大学张晓诗等在区域地质调查中首次发现, 并认为由蜥脚类恐龙所留(张晓诗等, 2016)。同年11月, 毕节市文物局戴犁和中国科学院古脊椎动物与古人类研究所张立召在响水乡进行野外考察时, 亦发现了路边岩石上的足迹。2016年12月, 毕节市文物局邀请中国地质大学(北京)学者考察该足迹点, 并在附近发现二号点。
大转弯足迹点一号点共有3条行迹, 其中保存最好的S1行迹共有17个足迹, 包括5个前足迹和12个后足迹, 前足迹平均长15.2 cm, 后足迹平均长35.2 cm。保存较好的前足迹呈椭圆形至弱U形, 没有明显的爪印, 跖趾区成凹形, 而后足迹为椭圆形, 长宽比为1.3, 无明显的爪印, 远端的不规则凹陷可能代表趾或爪印, 跖趾区后缘平滑弯曲。后足三角宽/后足长的值为1.5, 表明该行迹间距较宽, 整体形态可归入宽间距的Brontopodus type, 指示一类相对小型的蜥脚类造迹者(Xing et al., 2019a)(图 2, 表 1)。
大转弯足迹点二号点有1条不完整的行迹, 包括3个足迹, 其中只有1个前足迹完整, 余下1对前后足迹皆不完整。完整的前足迹呈U形, 足趾Ⅰ 、Ⅱ +Ⅲ 、Ⅳ 、Ⅴ 上有可识别的爪印, 长宽比为0.8。这批足迹从形态上亦可归为蜥脚类(Xing et al., 2019a)。
大转弯足迹点是贵州首次记录的侏罗纪恐龙足迹。该发现亦再次表明, 中国西南地区早侏罗世的蜥脚类恐龙足迹似乎具相当大的多样性, 包括窄间距的Parabrontopodus、宽间距Brontopodus type、以及basal sauropodomorph(基干蜥脚型类)足迹(Xing et al., 2019a)。
仁怀茅台岩滩足迹点位于贵州省仁怀市茅台镇岩滩村一酒厂内。足迹化石保存于下侏罗统自流井组马鞍山段中厚泥质岩屑石英砂岩层面上, 面积约350 m2, 足迹至少250个, 其中仅发现1个兽脚类足迹, 其他都是蜥脚类足迹(Xing et al., 2019b)。
2013年夏, 茅台镇赤水河畔一酒厂在建造制酒车间时, 考虑到其后侧边坡地带岩层不够牢固, 于是对边坡进行清理, 清理至下部时发现一个完整的岩石面, 质地坚硬, 有许多深浅不一的印痕。2017年夏, 该公司员工郭松波怀疑后山凹坑为恐龙足迹, 向贵州省博物馆和中国地质大学(北京)报告。2017年8月, 中国地质大学(北京)等单位学者前往化石点考察。2018年10月, 仁怀市政府对化石点用钢化玻璃覆盖保护。
岩滩足迹点至少有3层:最底层的砂岩上至少有10个足迹, 但只保留了不清晰的卵圆形轮廓; 中部砂岩层约保存了250个足迹, 这些足迹保存完好, 轮廓清晰; 上部砂岩层只有1个孤立的后足迹。在中部砂岩层共识别出行迹14条, 包括足迹97个, 均属于蜥脚类足迹。所有行迹近于平行, 反映恐龙为群居生活(Xing et al., 2019b)。
岩滩蜥脚类足迹可分为2个类型, 类型A的后足迹长32.5~39.0 cm, 平均长宽比为1.3, 趾的方向与后足中轴平行, 足迹在前部约46%~50%部位横向分割。前足迹为五趾型到半圆形, 长9~18 cm, 平均长宽比为0.6。前足迹以63° 向行迹中线外转, 明显大于后足迹的外转角度(约32°)。前足迹平均步幅角为87° , 小于后足迹的105° 。类型A与四川省古蔺县椒园足迹点下侏罗统自流井组的蜥脚类Liujianpus shunan(Xing et al., 2016)非常相似, 两足迹点亦相距较近, 椒园位于岩滩以西43 km处。Xing等(2019b)将类型A归入Liujianpus shunan(图 2, 表 1)。
类型B的形态特征与类型A截然不同, 缺乏前足迹和后足迹脚跟印, 只保留有细长、逐渐变细、平行的趾迹。保存较好的后足迹平均长31.5 cm, 长宽比为1.4。四趾中, 第Ⅱ 、Ⅲ 、Ⅳ 趾的趾迹较长较深, 第Ⅰ 趾的趾迹较短较浅。足迹后端保留有突起, 表明基底沉积物被足掌耙起并堆积在跟部。这种特殊的形态此前并没有被记录过, Xing 等(2019b)认为有3种可能性: (1)造迹者四足向两足的转换; (2)水体环境下的游泳行为; (3)幻迹。
该足迹点保存有舌状波痕、具有独特褶皱痕迹的微生物构造等沉积构造, 表明此处为不利于无脊椎动物栖息的半干旱环境。这与早侏罗世蜥脚类足迹通常与半干旱古环境有关(Xing et al., 2019b)相一致。
岩滩足迹点仅发现1个小型三趾型足迹。该足迹长20 cm, 长宽比为1.67, 形态上可归属于经典的兽脚类足迹Grallator(跷脚龙足迹)(Xing et al., 2019b)
赤水宝源足迹点位于贵州省赤水市宝源乡联华村, 属于赤水桫椤国家级自然保护区范围。足迹产自下白垩统上部夹关组(窝头山组)1块面积14 m× 11 m、几乎垂直的砂岩坍塌体上。
多年前, 当地人发现在联华村森林中的一些岩石上有形状像鸡爪的足迹, 村民们叫它“ 天鸡石” 或“ 仙鸡石” (Xing et al., 2011)。2009年, 邢立达赴实地考察, 在1块大型的、几乎垂直的砂岩坍塌体上发现多个恐龙足迹。2019年7月, 贵州省博物馆和中国科学院古脊椎动物与古人类研究考察队再赴化石点考察, 在周边又有新发现。
Xing等(2011)描述了7条兽脚类行迹和一些孤立的足迹。足迹共72个, 包括三趾和四趾型, 足迹的长度在15~19 cm之间, 长宽比为1, 每个足趾都有锋利的爪印, 可能的趾垫式为2-2-4-4-x, 步幅角为165° 。从形态上, 这批足迹暂时被归入cf. Irenesauripus isp.(图 2, 表 1)。这批足迹中大多数标本的跖趾区都保存了不同大小的、延长的跖骨印, 重复出现的跖骨印可能代表了一种特殊的沉积环境, 比如地面非常泥泞致恐龙行走困难才留下的独特痕迹。但是, 来自四川汉溪足迹点的同类足迹却表明, 这种行走步态对恐龙的速度并无明显的影响(Xing et al., 2015)。基于2011年及之前的夹关组足迹化石记录, Xing等(2011)认为当地早白垩世的恐龙动物群缺乏蜥脚类, 以兽脚类和鸟脚类为主, 但此后夹关组新发现的多处足迹点均发现了蜥脚类足迹(邢立达等, 2016)。
习水同民蔺江足迹点位于贵州省遵义市习水县同民镇蔺江村, 属于习水国家级自然保护区范围。足迹产自下白垩统上部夹关组, 分布在蔺江村同民河畔红色砂岩层面上。足迹共64个, 其中56个足迹可识别出10条行迹, 另外8个为孤立的足迹。
2016年7月, 同民镇干部袁锐在蔺江村调查时发现了这些奇怪的印记, 向相关部门报告。2017年5月, 贵州省博物馆自然部蔡回阳等前往考察。同年7月, 由贵州省博物馆、中国地质大学(北京)地球科学与资源学院、自贡恐龙博物馆组成的联合考察队考察该足迹点。
蔺江足迹点10条行迹中, 有1条保存良好的四趾型足迹行迹包括3个足迹, 平均长32.0 cm, 平均长宽比为1.3, 第Ⅲ 趾最长, 其次是第Ⅱ 趾和第Ⅳ 趾, 后中指向的第Ⅰ 趾平均长为10 cm, 趾垫式为1- 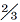
蔺江足迹点的其他兽脚类足迹的长度均在10~25 cm之间, 与经典的Eubrontes type足迹相似, 因此都可归入此类。其中1个孤立的足迹位于足迹层面西南端约12 m处, 该足迹长19.4 cm, 长宽比为1.0, 具有异常延伸的、长达35.2 cm的跖骨垫。
该足迹点另有9个三趾型鸟类足迹, 保存较好的足迹长11.2 cm, 长宽比为0.6~0.7, 第Ⅱ 趾与第Ⅳ 趾之间的趾间角为130° ~147° , 步幅角为134° ~154° 。该形态学与重庆綦江莲花保寨足迹点的Wupus非常相似, 但与Wupus agilis有一定差别, Xing 等(2018)暂且将其归为Wupus isp.。
2019年, 在上述9个足迹点之一的贞丰龙场又发现4个足迹, 对这4个新发现的足迹笔者开展了详细的考察和研究。
2019年8月, 贵州省博物馆曾嵘和中国科学院古脊椎动物与古人类研究所技术人员张鹏杰、桂友、王松在贞丰县龙场乡进行野外考察时, 在关兴公路西侧的贞丰县龙场镇第二中学对面岩壁层面上发现了4个趾痕明显的爬行动物足迹。同年9月, 贵州省博物馆邀请中国地质大学(北京)邢立达课题组前往现场考察。
考察发现, 该足迹点位于吕洪波等(2004)首次记录的龙场足迹点的上部约10 m处, 原足迹点可能在施工过程中被剥去, 因此无法确定与原行迹的关系。足迹点仍在关岭组松子坎段的中上部泥质白云岩表面, 该岩层表面发育泥裂构造, 并被后期2组节理截切。足迹点所在崖壁的地层褶皱变形, 出露一系列向斜、背斜以及逆断层等构造(图 3; 图 4)。
 | 图 3 贵州省贞丰县龙场乡中三叠统关岭组松子坎段龙场二号足迹点的卫星与野外照片 A—2016年4月卫星图, 绿点为龙场旧足迹点; B—2020年11月卫星图, 绿点为龙场旧足迹点, 红点为新足迹点; C——白圈内为新发现的龙场足迹Fig.3 Satellite and field images of the Songzikan Member of Middle Triassic Guanling Formation at No.2 Longchang track site in Longchang Township, Zhenfeng County, Guizhou Province |
 | 图 4 贵州省贞丰县龙场乡中三叠统关岭组松子坎段龙场二号足迹点的现场与足迹图 A—足迹层面; B—龙场新发现足迹的1个单步; C—与B对应的轮廓图Fig.4 Photographs and interpretative outline drawings of the Songzikan Member of Middle Triassic Guanling Formation at No.2 Longchang track site in Longchang Township, Zhenfeng County, Guizhou Province |
本课题组使用iPhone 7拍摄照片。使用三维图片重建软件Agisoft Metashape Professional Edition(v. 1.5.0)、数据分析可视化软件Paraview(v. 5.0.0)和三维点云处理软件CloudCompare(v. 2.6.1)制作了三维图片, 绘制了轮廓图。
在龙场新足迹点(文中称之为龙场二号足迹点)发现有至少4个足迹, 其中3个构成1条行迹, 编号为LC-T1-L1、LC-T1-R1和LC-T1-L2, 另1个为孤立足迹, 编号为LC-TI1(图 5)。4个足迹保存情况都不佳:LC-T1-L1长19.5 cm, 缺失第V趾末端, 第Ⅰ ~Ⅳ 趾宽15 cm; LC-T1-R1缺失第V趾跟部, 宽20 cm, 第Ⅰ ~Ⅳ 趾宽16 cm, 单步为74 cm; LC-TI1缺失第V趾, 第Ⅰ ~Ⅳ 趾宽10 cm。从尺寸上看, 龙场二号点行迹T1的足迹长约20 cm, 大于下部足迹点保存最好的足迹的长度(13 cm), LC-TI1则接近下部足迹点足迹的尺寸。从形态上看, 二号点足迹与先前发现的下部足迹点保存不好的足迹非常相似, 各个趾之间的隔离度较低, 第Ⅰ ~Ⅳ 趾较短且细, 第Ⅱ 趾与第Ⅳ 趾对称, 近乎等长, 中间的第Ⅲ 趾最长。龙场二号足迹点足迹的单步与下部足迹点的足迹基本一致, 表明其运动状态相似。综合以上形态学特征, 龙场二号足迹点的足迹可归入Chirotherium。LC-T1这种狭窄的行迹亦与德国图林根南部Hildburghausen附近Heß berg 地区的Chirotherium barthii模式标本行迹一致。
 | 图 5 贵州省贞丰县龙场乡中三叠统关岭组松子坎段龙场二号足迹点新发现足迹的照片(A, D, G), 三维照片(B, E, H)和轮廓图(C, F, I)Fig.5 Photographs(A, D, G), 3D images(B, E, H)and interpretative outline drawings(C, F, I) of newly found tracks of the Songzikan Member of Middle Triassic Guanling Formation at No.2 Longchang track sitein Longchang Township, Zhenfeng County, Guizhou Province |
对贵州贞丰龙场二号足迹点Chirotherium足迹的3D化信息采集, 为这些保存不佳的足迹提供了更多的形态学细节, 如各趾的深度等。龙场二号足迹点与龙场足迹点足迹尺寸的差异, 亦表明该地区有更多个体的Chirotherium造迹者活动, 增加了其丰度。
从形态上看, Chirotherium的造迹者代表了chirotheriid(手兽科)中最发达的后足迹形态, 并展示了向功能性三趾型足迹演化的趋势(Haubold and Klein, 2002; Klein and Lucas, 2021; Klein et al., in press)。其潜在的造迹者被认为是早期的avemetatarsalians(鸟跖类)和/或crocodylian-stem archosaurs(主龙类鳄类干群), 它们演化出类似的后足结构, 例如一些poposauroids(波波龙超科)成员(Farlow et al., 2014; Klein and Lucas, 2021)。
贵州贞丰牛场、龙场和龙场二号足迹点的Chirotherium足迹总数为72个, 造迹者为6个, 后足迹长度13~25 cm, 具有较高的丰度(表 1)。就化石记录而言, Chirotherium目前在中国南部的分布亦较为广泛, 除了贵州的记录外, 还包括云南会泽和四川攀枝花的发现。
云南会泽Chirotherium发现于中三叠统关岭组松子坎段, 包括了4条Chirotherium barthii行迹, 足迹长10~19 cm, 大部分足迹成对出现, 形态与欧洲经典的C. barthii相一致, 另外该足迹点还发现有亚洲首次记录的Rhynchosauroides足迹(Xing and Klein, 2021)。四川攀枝花的Chirotherium发现于上三叠统宝顶组, 足迹尺寸较大, 后足迹长达41 cm, 明显大于贵州标本, 但形态与中三叠世的Chirotherium非常相似, 如后足迹的第Ⅱ ~Ⅳ 趾紧凑, 呈对称分布, 第Ⅲ 趾最长, 还有1个指向后侧方、细长的第V趾。Xing 等(2014)将其暂归入Chirotherium, 不排除这批足迹代表了1个新的遗迹种。云贵川的足迹记录表明, Chirotherium广泛活跃于该区域, 尤其是关岭组的记录最为丰富。
关岭组的沉积环境为炎热干燥气候下的浅海潮间带和潟湖, 与德国西南部中三叠统Muschelkalk(Anisian-Ladinian)碳酸盐岩斜坡相似, 后者发现了丰富的海生爬行类骨骼化石与陆生动物足迹, 但缺乏海生爬行类足迹(Klein and Lucas, 2018)。
目前, 贵州省已发现的9个足迹点记录了近991个恐龙等四足类的足迹, 这代表着约477个造迹者, 包括海生爬行类、手兽足迹类、非鸟兽脚类、鸟类、蜥脚类和鸟臀类。这个数据集很好地补充了贵州省中生代的四足类动物群面貌, 尤其是贵州省目前除了海生爬行类之外的脊椎动物化石记录较少, 故足迹化石的作用更加明显, 这点尤其体现在无骨骼化石发现的白垩系“ 哑地层” 中。
牛场和龙场三叠系关岭组记录的Chirotherium是亚洲首次记录的此类足迹, 其中Chirotherium常见于欧洲、北美、南美以及北非的中三叠统, 正如上文所分析的, 这些发现是Chirotherium造迹者在欧亚大陆广泛分布的重要证据之一。但关岭组更难得的足迹记录是安顺幺铺的海生爬行类足迹。由于保存条件苛刻, 海生爬行类的水下足迹很少保存, 这彰显了安顺幺铺足迹动物群的重要性, 而大量的足迹忠实记录了海生爬行类的水下活动, 更是首次发现的关于楯齿龙类或龙龟类动物水下行为的记录, 反映了海生爬行类早期采用“ 撑行” 这种肢体协同运动的游泳方式。同时, 足迹化石组合表明, 幻龙类曾与楯齿龙类或龙龟类动物一同出现在东特提斯海沿岸的潟湖等浅海环境中觅食, 这为了解海生爬行类在中三叠世浅水或潟湖底部的生活习性和运动情况提供了证据(Xing et al., 2020)。
侏罗纪的恐龙骨骼化石目前在贵州省有零星发现, 其面貌与四川盆地相似, 恐龙足迹的记录亦体现了这个特点。下侏罗统龙头山群(相当于四川盆地珍珠冲组)发现的蜥臀类足迹, 以保存完好的中型Eubrontes兽脚类足迹为主, 并伴有一些蜥脚类和可能的鸟臀类恐龙足迹。这种以蜥臀类为主导的足迹动物群(表 1)常见于中国与世界各地的早侏罗世生物带, 也与当地极少发现的骨骼化石记录一致。这个发现也表明, 较小的足迹点更有利于留存那些活跃的造迹者, 比如兽脚类恐龙, 而蜥脚类恐龙在同样面积的足迹点中的丰度往往更低。茅台岩滩发现的大面积蜥脚类足迹是中国目前发现的早侏罗世规模最大的蜥脚类恐龙足迹化石群, 该处发现的Liujianpus足迹形态介于基干蜥脚类与蜥脚类之间, 具有明显的时代特点。这种形态亦发现于四川古蔺。毕节响水大转弯足迹点的足迹则再次表明, 中国西南部早侏罗世的蜥脚类恐龙足迹似乎具有相当大的多样性, 包括窄间距的Parabrontopodus、宽间距Brontopodus type、以及Liujianpus这类可能的basal sauropodomorph足迹(Xing et al., 2019a)。而且, 这一生态面貌与同期四川盆地的记录一致, 表明中国西南早侏罗世有着类似的蜥脚类恐龙动物群(Xing et al., 2019a)。
白垩纪, 贵州的恐龙足迹记录与四川盆地非常相似。赤水宝源的非鸟兽脚类足迹化石是贵州省首次记录的白垩纪恐龙足迹, 填补了贵州省该时期的足迹化石记录空白(Xing et al., 2011)。虽然夹关组的足迹记录在四川盆地已有相当丰富的发现, 但蔺江足迹点Gigandipus形态类型的出现, 依然丰富了该组的兽脚类足迹的多样性, 也表明Grallator、Eubrontes、Anomoepus、Gigandipus等北美侏罗纪常见的因子在中国早白垩世仍然有广泛的存在。蔺江的Wupus isp.表明, Limiavipedidae这种大型古鸟类造迹者在中国西南地区的分布比之前想象的更广泛, 其与加拿大的Limiavipes以及南澳大利亚的大型水鸟足迹相似。这些发现再次表明, 夹关组保存了高度多样化的兽脚类足迹, 其仍然是四川盆地四足动物记录主要的、重要的来源, 很好地弥补了该盆地极少发现白垩纪骨骼化石的遗憾。
1)由69条行迹和420个孤立足迹组成的、总数达991个足迹的四足类遗迹化石指示了至少477个造迹者, 这些造迹者包括海生爬行类、手兽足迹类、非鸟兽脚类、鸟类、蜥脚类和鸟臀类等。这一数据集为贵州省中生代古生物类群的多样性研究提供了重要的信息。
2)三叠纪, 贵州省的四足类遗迹化石中的Chirotherium无对应的骨骼记录, 但鉴于该遗迹属的广泛分布, 因此具有全球对比意义; Dikoposichnus和Anshunpes为同时期海生四足类骨骼化石记录增加了可对应的运动学记录, 后者是世界首次记录的楯齿龙或龙龟类遗迹化石记录。未来工作重点是在寻找更多的足迹记录的同时, 深挖已发现遗迹化石的信息, 如模拟古动物运动方式等。
3)侏罗纪, 贵州省的足迹集中在中下侏罗统, 数量明显多于该地区稀少的骨骼化石记录。足迹化石提供了更丰富的多样性, 且表现为确凿的蜥臀类主导, 尤其是蜥脚型类的集合, 体现了由基干蜥脚型类主导转化至真蜥脚类主导的演化趋势。
4)白垩纪, 贵州省的足迹记录与四川盆地的遗迹记录基本一致, 但亦有新的形态作为有益的补充。
5)贵州中生代的四足类遗迹化石与与四川盆地等西南地区整体的足迹群组合十分相似, 在沉积良好的区域仍有潜力发现新的足迹形态, 从而增加四足类足迹的多样性, 最终结合骨骼化石, 展现更加全面的贵州中生代古动物群面貌。
致谢 感谢贵州省博物馆的诸位同仁邀请并协助了多次野外工作, 澳大利亚昆士兰大学的安东尼· 罗米里奥(Anthony Romilio)对本文数据亦有贡献。
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|