

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
云南早泥盆世植物稳定碳同位素特征及其古气候意义*
[张世辉1  , 肖良
, 肖良2 , 薛进庄1 , 孟美岑3 , 秦敏4 , 崔莹5 , 王德明1 ]
, 肖良, 王德明]
|
|

第一作者简介 张世辉,男,1995年生,北京大学地球与空间科学学院硕士研究生,主要从事古环境研究。E-mail: zsh16390@163.com。
受到植物化石材料局限性与时代久远性的限制,目前用古生代植物化石δ13C值来反演古环境的研究程度较低。文中通过对云南省曲靖市下泥盆统徐家冲组和文山州下泥盆统坡松冲组的部分植物化石进行稳定碳同位素测试,分析了早泥盆世植物的光合作用途径以及古环境。结果表明: 徐家冲组镰蕨属(Drepanophycus)δ13C值为-29.0‰~-22.8‰,平均值为-24.5‰,其中镰蕨属茎轴的δ13C值高于叶片0.1‰~0.3‰;坡松冲组工蕨属(Zosterophyllum)和始叶蕨属(Eophyllophyton)δ13C值分别为-24.8‰~-22.1‰和-23.0‰~-22.6‰,平均值分别为-23.2‰和-22.7‰;三者的δ13C值分布范围都与现代C3植物一致。自徐家冲组第57至第79沉积旋回,δ13C值总体呈上升趋势,达到2‰左右,推测可能与早泥盆世大气CO2浓度下降和气候变冷相关。镰蕨属和工蕨属的Ci/Ca值较为稳定,说明这2种植物可能对大气CO2浓度变化较为敏感,曲靖市古气候可能为当今的热带疏林草原气候或热带季风气候。
About the first author Zhang Shi-Hui,born in 1995,is a postgraduate in School of Earth and Space Sciences,Peking University. He is mainly engaged in research of palaeoenvironment. E-mail: zsh16390@163.com.
Because of the availability of plant fossil materials in the deep past,palaeoenvironment inversion using δ13C value of Paleozoic plant fossils is relatively poorly studied. In this paper,the stable carbon isotopes of some fossil plants are analyzed from the Lower Devonian Xujiachong Formation in Qujing area and from the Posongchong Formation in Wenshan area,Yunnan Province,China. The δ13C values of Drepanophycus from the Xujiachong Formation range from-29.0‰ to -22.8‰ with an average value of-24.5‰. The δ13C values of Zosterophyllum and Eophyllophyton from the Posongchong Formation range from -24.8‰ to -22.1‰ and -23.0‰ to -22.6‰,and their average values are -23.2‰ and -22.7‰,respectively. All values fall within the distribution range of the δ13C values of modern C3 plants. The δ13C values of different organs of Drepanophycus are dissimilar,and the δ13C value of the axis is 0.1‰ to 0.3‰ higher than that of the leaf. From the 57th to the 79th sedimentary cycles of the Xujiachong Formation,the δ13C value of Drepanophycus increases by about 2‰,which is interpreted as relating to a decrease in the palaeoatmospheric CO2 concentration and to the climate cooling. The calculated Ci/Ca values of Drepanophycus and Zosterophyllum are relatively stable across the studied interval. Thus,it is inferred that these two plants are sensitive to the change of palaeoatmospheric CO2 concentration and the palaeoclimate in Qujing area may be the present tropical savanna climate or tropical monsoon climate.
开放科学(资源服务)标识码(OSID) 
气候变化是当今最为引人关注的环境问题之一, 众多学者通过各种途径对它进行解释与预测, 以便为人类社会的发展提供参考依据, 但人类气象记录的历史不过几百年, 难以从中找出长时间尺度的规律, 因此古气候和古环境的研究逐渐受到学术界普遍关注(刘贤赵等, 2014)。目前, 多种地质记录被用来分析古气候与古环境变化, 如冰芯(Petit et al., 1999; 段克勤等, 2012; 徐陈鹏等, 2019)、古土壤(Kelly et al., 1998; 刘瑾等, 2015)、树木年轮(Chen et al., 2011, 赵少军等, 2017)及湖泊和海洋沉积物(袁子能等, 2012; 钟阿娇等, 2020)等。
植物叶片化石一直是重建古气候的重要材料之一, 前人主要利用叶片的气孔参数(包括气孔密度和气孔指数)、叶相特征、植物群组合等获取古环境信息(肖良等, 2017a)。近年来, 越来越多的学者将有机地球化学方法用于对现代植物的研究, 其中叶片稳定碳同位素对环境变化响应的研究方法已相对比较成熟(Foster et al., 2017)。植物δ 13C值常受到内因与外因的共同影响, 内因是自身的遗传因素, 外因是环境, 如温度(Wang et al., 2013)、水分(Kohn, 2010)、大气CO2浓度(Schubert and Jahren, 2012, 2018; Chapman, 2019; Cui et al., 2020)等。目前已开展了多种植物的δ 13C值与环境因子之间关系的研究, 这为利用植物化石δ 13C值进行古气候和古环境研究提供了参考依据(肖良等, 2017b), 但由于影响植物碳同位素组成的各环境因子之间关系错综复杂, 两者之间的关系也存在许多争议(冯虎元等, 2000; 郑淑霞和上官周平, 2006; 刘贤赵等, 2014)。
泥盆纪是植物登陆后演化的一个关键时期, 陆生植物的先驱分子开始出现, 初步形成复杂多样的生态系统, 使得陆地环境得到极大改变; 同时, 植物属种多样性倍增, 一些关键性状如根、叶等快速演化(Hao and Xue, 2013; 薛进庄和郝守刚, 2014)。
近年来, 众多学者对中国泥盆纪地层不断进行研究, 发现了越来越多的植物化石种类, 对植物谱系关系与分类方案的认识愈加完善。其中早泥盆世的植物大多矮小、根系不发达、受限于潮湿低洼的生长环境, 而到了晚泥盆世, 陆生维管植物已拥有了叶片、复杂根系等, 并将栖息地扩展到高地(Wan et al., 2019)。这一巨大的改变, 导致了有机碳埋藏、大气中O2含量升高和CO2浓度降低, 引发全球气候变冷(Le Hir et al., 2011)。
迄今为止, 对植物化石稳定碳同位素的研究相对很少(肖良等, 2017a), 且利用植物化石δ 13C值来反演古环境的研究主要集中在中生代、新生代。由于受到植物化石材料的局限性与时代久远性的限制, 用古生代植物化石δ 13C值来反演古环境的研究程度较低, 尚有许多空白(冯净等, 2014)。前人对于泥盆纪陆生植物稳定碳同位素进行了少量的研究, 研究对象主要为真菌(Boyce et al., 2007)、地衣(Fletcher et al., 2004)、苔类(Graham et al., 2010)和维管植物(Beerling et al., 2002; 冯净等, 2014)。Wan等(2019)对泥盆纪和早石炭世大量维管植物进行了系统研究, 但研究区主要集中在国外, 国内早泥盆世维管植物稳定碳同位素的研究十分罕见。
中国早泥盆世非海相地层主要分布在西南地区的云南、贵州、四川、广西等省自治区, 以滇东和滇东南最为发育(刘振峰等, 2004)。徐家冲剖面与纸厂剖面是较为典型的2个剖面(图 1), 地层出露较好, 植物化石众多、形态保存良好。其中, 镰蕨属在滇东徐家冲组中保存最多且最完整(李星学和蔡重阳, 1978), 能为逐层分析同一属种的植物化石稳定碳同位素变化提供丰富材料; 滇东南坡松冲组植物群以丰富的形态明显分异的工蕨类植物为特征(郝守刚和王德明, 2003)。因此, 笔者选取镰蕨属与工蕨属为研究对象, 并辅以同时采集到的始叶蕨属, 开展稳定碳同位素组成及其与环境之间关系的研究, 探讨这3类植物的光合作用途径、不同器官间稳定同位素的差异以及早泥盆世碳同位素的变化趋势。
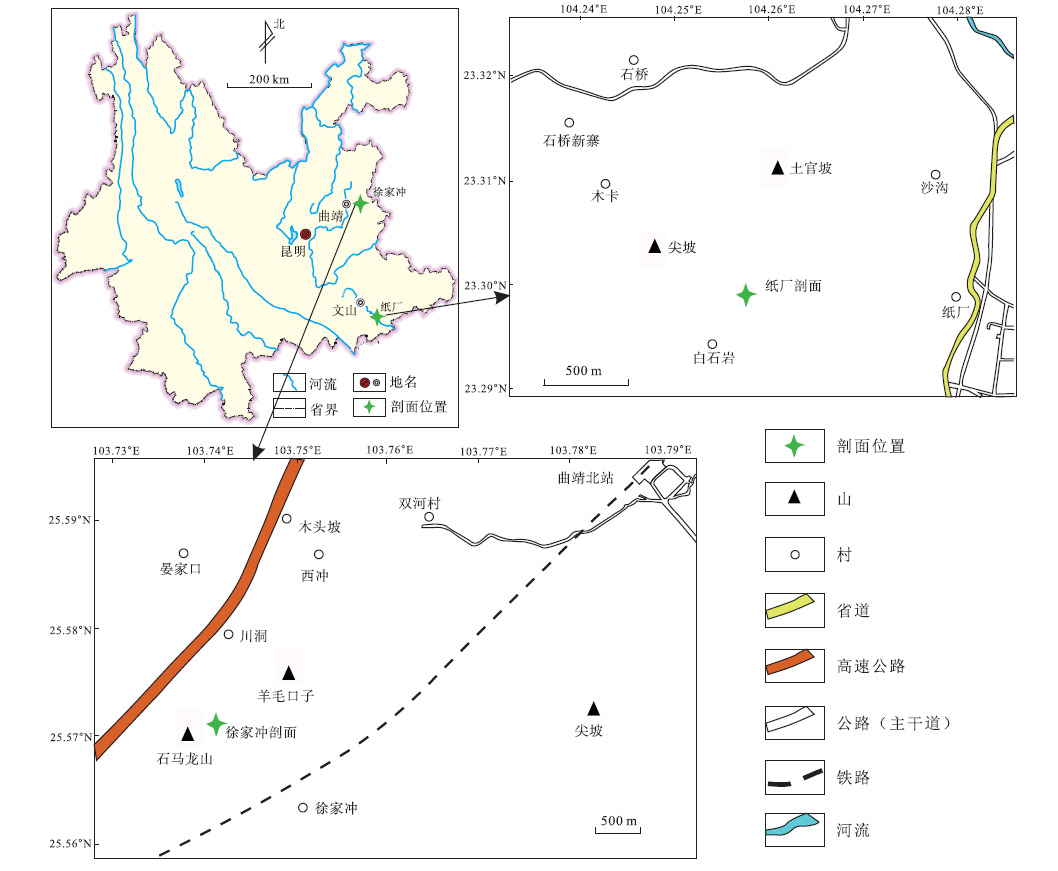 | 图 1 云南省早泥盆世植物化石产地Fig.1 Location of the Early Devonian plant fossil in Yunnan Province |
中国早泥盆世非海相地层在滇东和滇东南出露最为完整。经过众多学者近一个世纪的努力, 云南省东部曲靖地区成为该时期地层的1个标准剖面所在地(刘振峰等, 2004)。
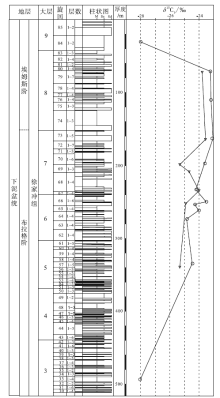
下泥盆统徐家冲组出露于曲靖市西北约13 km的徐家冲等剖面。徐家冲剖面位于昆明市东北约167 km的曲靖市西山乡徐家冲村北的山丘中, 地层沿着徐家冲村至川洞村的山路分布, 出露较好。刘振峰等(2004)测得的徐家冲剖面徐家冲组厚841.84 m, 基于岩性被划分为93个沉积旋回, 并可合并成9个大层(图 2)。李星学和蔡重阳(1978)最早测制了该套地层, 共划分为8大层; 同时在徐家冲村东北约6 km的沾益县龙华山剖面发现了相应的地层, 称作龙华山组。龙华山组下部被掩盖, 仅厚223.8 m, 岩性和植物组合相当于徐家冲组的上部。
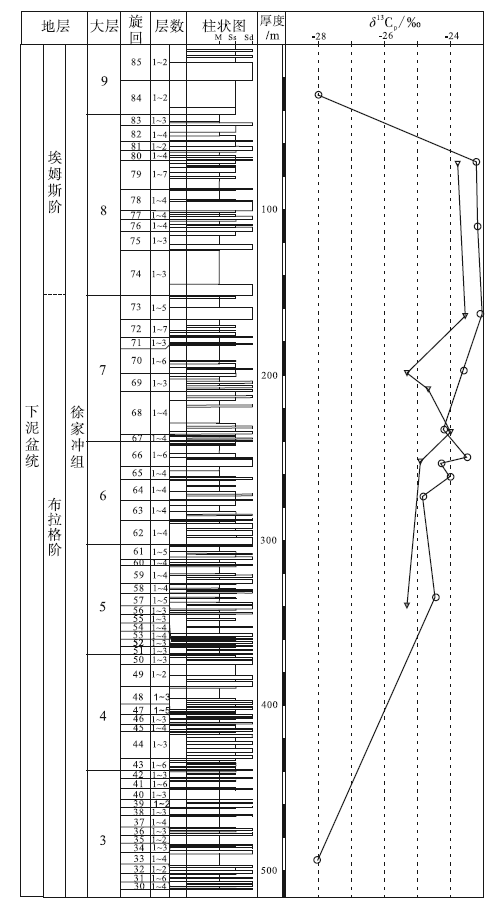 | 图 2 云南省曲靖市徐家冲剖面下泥盆统徐家冲组地层柱状图(改自刘振峰等, 2004)及镰蕨属δ 13C值(旋回依据岩性划分, 层数为每个旋回所包含的地层数)Fig.2 Stratigraphic column of the Lower Devonian Xujiachong Formation(modified from Liu et al., 2004)and δ 13C values of Drepanophycus at Xujiachong section in Qujing City, Yunnan Province(cycles are divided by lithology, layer is the number of strata contained in each cycle) |
徐家冲组与下伏的下泥盆统桂家屯组呈整合接触, 与上覆的中泥盆统穿洞组呈平行不整合接触。徐家冲组由砂岩、粉砂岩、泥岩和粉砂岩构成四元结构的沉积旋回, 每个旋回从砂岩开始, 以粉砂岩或泥岩结束, 可能指示三角洲平原靠陆部分的河流环境(刘振峰等, 2004; Xue et al., 2016)。对动植物大化石(Wang et al., 2002)、孢子组合(Wellman et al., 2012)以及生物地层系数(biostratigraphic coefficient)(Wang, 2007)的研究表明, 徐家冲组的中下部属于布拉格期晚期, 上部属于埃姆斯期早期。
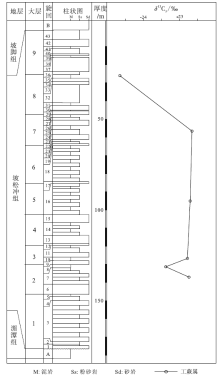
坡松冲组广泛分布在滇东南文山、广南、蒙自、砚山等地区, 并且延伸到越南境内。该组在文山州古木镇纸厂村保存最好, 众多植物化石被发现于此。在纸厂剖面, 坡松冲组与下伏的下奥陶统湄潭组呈角度不整合接触, 与上覆的下泥盆统坡脚组呈整合接触(图 3)。坡松冲组下部地层为灰色、灰黄色粗粒和细粒砂岩, 代表河流沉积; 上部为大量的黑色页岩或泥质粉砂岩, 多有泥质透镜体, 代表近岸的湖相或潟湖沉积(郝守刚和王德明, 2003)。对植物化石(郝守刚和王德明, 2003; Hao and Xue, 2013)、孢子组合(Cascales-Miñ ana et al., 2017)及生物地层系数(Gerrienne, 1996)的研究表明, 坡松冲组地层属于早泥盆世的布拉格期晚期。
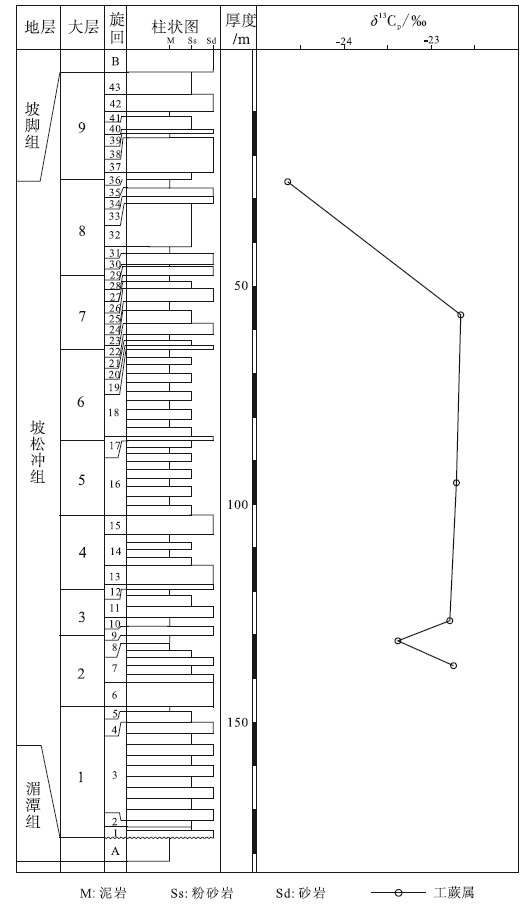 | 图 3 云南省文山州纸厂剖面下泥盆统坡松冲组地层柱状图(改自Hao and Xue, 2013)及工蕨属δ 13C值Fig.3 Stratigraphic column of the Lower Devonian Posongchong Formation(modified from Hao and Xue, 2013)and δ 13C values of Zosterophyllum at Zhichang section in Wenshan Prefecture, Yunnan Province |
植物化石采自中国云南省曲靖市徐家冲剖面徐家冲组和文山州纸厂剖面坡松冲组。徐家冲剖面徐家冲组镰蕨属样品共计105个, 分别采自第33-2层、57-3层、64-2层、65-2层、66-1层、66-2层、68-2层、70-1层、73-1层、76-2层、79-7层和84-2层(层号据刘振峰等, 2004)。文山州纸厂剖面坡松冲组工蕨属样品共计28个, 采自第2大层、第3大层、第5大层、第7大层、第8大层; 始叶蕨属样品4个, 采自第3大层(层号据Hao and Xue, 2013)。
在选取植物化石进行有机碳稳定同位素分析之前, 首先对样品进行预处理: (1)用针将植物炭膜取下(必要时, 在体视显微镜下进行操作), 装进玻璃瓶中封存; (2)将储存的炭屑放入蒸馏水中浸泡约1 h, 以去除附着的沉积物; (3)将样品转移入盐酸中, 浸泡约12 h, 在反应过程中用玻璃棒多次搅拌, 至无机碳酸盐与盐酸充分反应; (4)将样品反复用蒸馏水冲洗至中性, 倒入玻璃培养皿中自然干燥约10 h, 再转移到干燥的玻璃瓶中储存。
本次样品共分3批进行测试。第1批和第2批样品于中国石油勘探开发研究院石油地质实验研究中心测试, 使用仪器为Finngan MAT-252, 测试条件为将样品制备成纯CO2气, 用MAT-252双进样法与GBW04405参考气比较测试给出相对PDB的值, 执行标准为《有机物和碳酸盐岩碳、氧同位素分析方法》SY/T5238-2008。第3批样品于核工业北京地质研究院测试, 仪器型号为MAT-253气体同位素质谱仪, 检测方法与依据为GB/T 18340.2-2010《地质样品有机地球化学分析方法 第2部分: 有机质稳定碳同位素测定同位素质谱法》。
碳同位素(δ 13C)的定义如下:
植物光合作用在合成有机质的过程中优先吸收大气中碳的轻同位素12C, 因此碳同位素在植物体内外产生分馏, 植物体内相对富集轻同位素12C, δ 13C值相对于大气偏负。光合作用中植物的碳同位素分馏不仅与植物自身因素有关, 而且还受到周围环境条件的影响, 这是利用碳同位素δ 13C值分析环境信息的基础。植物光合作用产生的碳同位素分馏主要受3个因素控制: 光合作用的方式、物理化学变化过程中造成的分馏和环境因素。
1)光合作用的方式。根据光合作用方式的不同, 将植物分为C3植物、C4植物和CAM植物。这3类植物碳同位素值分布范围不同(Deines, 1980; 马晔和刘锦春, 2013; 冯净等, 2014), 其中C3植物δ 13C值一般为-20‰ ~-32‰ , C4植物δ 13C值一般为-7‰ ~-15‰ , CAM植物δ 13C值一般为-10‰ ~-22‰ (马晔和刘锦春, 2013)。泥盆纪时植物刚登陆不久, 正处于演化的关键时期, 光合作用的方式一般被认为是C3途径。至石炭纪, 陆生植物繁盛, 大气处于低CO2、高O2浓度的状态, 这可能会促使植物演化出C4或CAM光合作用方式, 然而目前并未在石炭纪发现C4植物和CAM植物的证据, 两者直到新生代才开始大量出现(Keeley et al., 2012)。
2)物理化学变化过程中造成的分馏。Farquhar等(1980, 1989)提出了C3植物的碳同位素分馏公式:
其中,
3)环境因素。环境的变化会引起植物光合作用效率等的变化, 从而影响植物δ 13C值。环境因素很多, 包括土壤、降水、海拔、光照、温度、大气CO2等。单一环境因子对植物δ 13C值的影响目前未有明确的定论, 以水分为例, 一般认为在缺水的环境下, 植物的气孔关闭, 细胞间的CO2分压降低, 导致δ 13C值升高, 但也有例外(Diefendorf et al., 2010; 刘贤赵等, 2014), 因此对植物δ 13C值与环境因素之间的关系仍需进一步研究。
在环境因素中, 大气CO2浓度和
基于公式(1)和公式(2), 可以通过测量植物化石的碳同位素值
然而, Schubert和Jahren(2012)通过在控制CO2浓度的实验装置中培养拟南芥Arabdopsis和萝卜Raphanus, 观察到碳同位素的分馏(Δ 13C=(
该研究结果已被用于重建冰期— 间冰期的大气CO2浓度(Schubert and Jahren, 2015)、过去23 Ma的CO2浓度变化(Cui et al., 2020)、古新世— 始新世CO2浓度(Cui and Schubert, 2018; Chapman et al., 2019)以及白垩纪CO2浓度(Barral et al., 2017)。Cui 等(2020)认为, 该公式具有应用到重建早古生代CO2浓度的潜力。
对采自徐家冲组和坡松冲组的植物化石(图 4)镰蕨属、工蕨属和始叶蕨属的形态描述, 参考孙克勤等(2010)的《中国植物化石志》进行。
 | 图 4 云南早泥盆世植物化石照片 A— 镰蕨属, 箭头所指为叶, 刺状, PKUB21001; B— 始叶蕨属, 茎轴等二叉或不等二叉分枝, PKUB21007; C— 始叶蕨属, 箭头所指为顶端回弯的末级附属物, PKUB21008; D— 工蕨属, 箭头所指为二歧分叉的直立枝, PKUB21004Fig.4 Photos of plant fossils of the Early Devonian in Yunnan Province |
镰蕨属, 草本。根状茎平卧而分叉, 常二歧分枝。叶刺状, 螺旋排列或散生, 直或镰刀形, 长1~2 cm, 具1条维管束; 基部扩大呈圆锥状。茎中央的维管束纤细而简单(图 4-A)。本次采集的样品多保存茎轴, 少数保存有刺状叶, 保存情况不理想, 难以区分器官。
工蕨属, 植物体矮小, 簇状丛生, 基部分叉很多, 形式特殊, 成“ H” 形或“ K” 形, 并由此伸出二歧分叉的直立枝(图 4-D)。孢子囊圆形至肾形, 具短柄, 螺旋排列于直立枝的顶部, 常呈穗状。本次采集的样品多保存簇状丛生的枝, 难见孢子囊。
始叶蕨属, 植物体矮小。茎轴等二叉或不等二叉分枝, 三维生长。一些茎轴具营养叶和生殖叶, 另一些具一次或二次分叉的、顶端回弯的末级附属物(图 4-B, 4-C)。本次采集的样品多保存等二叉或不等二叉分枝的茎轴, 偶见末级附属物。
测试结果(表 1)表明, 云南早泥盆世植物化石δ 13C值分布范围大多位于-22‰ ~-25‰ 之间, 不同剖面的植物化石δ 13C值分布有所差异。
| 表 1 云南早泥盆世植物化石δ 13C值汇总表 Table 1 δ 13C values of plant fossil of the Early Devonian in Yunnan Province |
徐家冲组的植物化石为镰蕨属, 其δ 13C值除了在第33与第84旋回为-28.1‰ 外, 在第57至第79旋回中分布在-24.8‰ ~-23.0‰ 之间, 平均值为-23.8‰ (图 2)。自下而上, 徐家冲组植物化石的δ 13C值总体呈现上升趋势。对镰蕨属不同器官之间的δ 13C值进行测试发现, 叶片的δ 13C值范围为-24.4‰ ~-23.1‰ , 平均值为-23.7‰ , 而茎轴的δ 13C值范围为-24.2‰ ~-22.95‰ , 平均值为-23.6‰ 。相比于茎轴, 叶片更富集12C, 其δ 13C值偏负。
坡松冲组工蕨属的δ 13C值分布范围为-24.8‰ ~-22.1‰ , 平均值为-23.2‰ (图 3)。在自下而上的垂向序列中, 工蕨属δ 13C值先后经历了下降、上升、缓慢上升和再下降的过程, 总体未有明显的变化趋势。始叶蕨属δ 13C值分布范围为-23.0‰ ~-22.6‰ , 平均值为-22.75‰ 。
根据光合作用途径的不同, 可将现代陆生植物分为C3植物、C4植物与CAM植物(Deines, 1980)。陆生植物大多为C3植物, 主要包括灌木、乔木和寒冷地区的草本植物, δ 13C值一般在-20‰ ~-32‰ 之间(平均值为-26‰ ); 少数为C4植物, 主要包括热带的草本(禾本科、莎草科)和小部分灌木(大戟科、藜科)等, δ 13C值一般为-7‰ ~-15‰ (平均值为-12‰ ); 其余为CAM植物, 主要包括沙漠肉质植物和热带附生植物, δ 13C值一般为-10‰ ~-22‰ (平均值为-16‰ )(马晔和刘锦春, 2013)。从这3类植物的δ 13C值分布范围可以看出, C3、C4植物的δ 13C值分布范围不重合, 而CAM植物极为少见, 因此一般可以用δ 13C值来判断某种植物的光合作用途径。
研究区徐家冲组植物化石的δ 13C值为-29.0‰ ~-22.8‰ , 坡松冲组植物化石的δ 13C值为-24.8‰ ~-22.1‰ , 都位于现代C3植物δ 13C值分布范围区间, 这表明早泥盆世的镰蕨属、工蕨属和始叶蕨属光合作用途径与现代C3植物相同或相似。
Wan(2012)测定了泥盆纪不同时期不同地区众多植物化石的δ 13C值, 其中不同地区镰蕨属的碳同位素δ 13C平均值分别为: -25.8‰ (德国)、-24.8‰ (中国新疆)、-25.9‰ (加拿大魁北克)、-26.5‰ (加拿大新不伦瑞克)、-25.3‰ (美国缅因)、-25.6‰ (美国纽约)。可以看出, 不同地区同一属的δ 13C值最大值与最小值相差1.7‰ , 大多集中在-25‰ ~-26‰ 之间。除此之外, Wan(2012)还以魁北克为例, 测定了泥盆纪时期同一地区的不同属植物之间的碳同位素δ 13C平均值, 发现其均值存在差异: -25.9‰ (Drepanophycus)、-27.1‰ (Sawdonia)、-24.2‰ (Psilophyton)和-28.1‰ (Archaeopteris), 其中数据差异最大的近4‰ , 最小的也有1‰ 。上述资料表明, 在同一时期, 地理环境因素可以对同一个属的植物化石δ 13C值产生影响, 并且同一地区不同属植物化石δ 13C值也有所差异。相对于植物属种而言, 地理位置影响程度较低, 即同一地区不同属之间的碳同位素值差异远大于不同地区同一属之间的差异, 这说明相对于环境因素来说, 植物本身遗传因素对碳同位素的影响更大。冯净等(2014)对新疆北部泥盆纪植物化石的研究也表明了这一观点。
由于徐家冲组样品所在的层位岩性条件非常接近, 都为灰绿色粉砂岩或石英砂岩中的灰绿色粉砂岩夹层, 沉积环境类似, 因此若成岩作用对δ 13C值产生影响, 那么其影响应该是相似的, 故成岩作用对本次研究的影响可以不予考虑。同样, 坡松冲组样品的成岩作用亦可忽略。在这一前提下, 笔者对测试样品的δ 13C值进行进一步分析。
测试样品中, 徐家冲组镰蕨属的δ 13C平均值为-24.5‰ , 坡松冲组工蕨属的δ 13C平均值为-23.2‰ , 坡松冲组始叶蕨属的δ 13C值为-22.7‰ , 这表明同一地理区域不同属植物化石的碳同位素δ 13C值存在差异, 其中始叶蕨属的δ 13C值高于工蕨属。但三者的δ 13C平均值差别较小, 最大约1‰ , 小于Wan(2012)所列举的植物化石δ 13C值差异, 推测这可能是由于笔者研究的植物属在亲缘关系上相对于Wan(2012)所列举的植物属更接近。依据文中的测试结果与Wan(2012)的数据, 可知镰蕨属δ 13C值已知的范围大致在-27‰ ~-23‰ 之间。
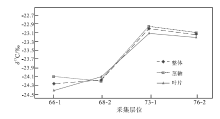
同一现生或化石属种植物的不同器官亦表现出不同的δ 13C值(Benner et al., 1987; Martinelli et al., 1998; 冯净等, 2014)。本次研究的植物化石部分保存不好, 导致对植物器官的区分较为困难。以徐家冲组植物化石为例, 在已区分不同器官的4个层位中(图 5), 除了第68-2层中植物茎轴的δ 13C值比叶片偏负0.1‰ 外, 其余3个层位中, 叶片δ 13C值均比茎轴偏负。其中, 第66-1层、第73-1层、第76-2层的叶片比茎轴的δ 13C值分别偏负约0.3‰ 、 0.1‰ 、 0.1‰ 。
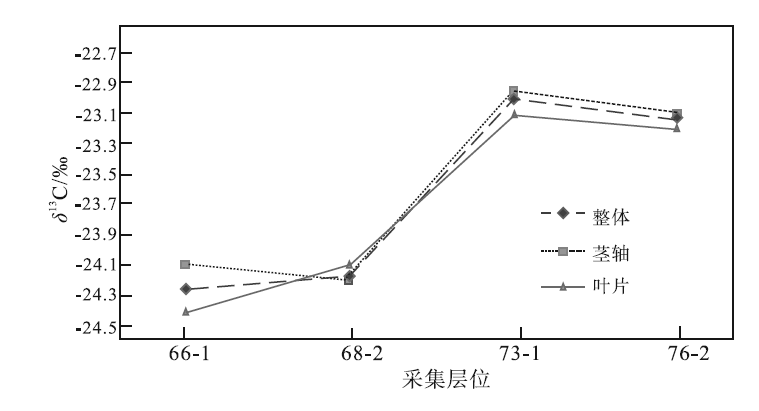 | 图 5 云南省曲靖市徐家冲剖面下泥盆统徐家冲组镰蕨属不同部位的δ 13C值 虚线表示镰蕨属整体的δ 13C值, 不区分器官; 点线表示镰蕨属茎轴的δ 13C值; 实线表示镰蕨属叶片的δ 13C值Fig.5 δ 13C value of different parts of Drepanophycus of the Lower Devonian Xujiachong Formation at Xujiachong section in Qujing City, Yunnan Province |
植物不同器官具有不同的生物化学组分, 进而表现出不同的生理生态特征, 并具有不同的δ 13C值(李明财等, 2007)。其中植物的光合作用发生在叶片中, 合成的有机产物主要为糖类, 而茎轴中的有机物则以木质素、半纤维素为主。相对来说, 木质素、半纤维素的δ 13C值高于可溶性糖的δ 13C值, 因此, 植物叶片的δ 13C值会表现为偏负于茎轴的δ 13C值(Francey et al., 1985)。至于第68-2层镰蕨属茎轴的δ 13C值比叶片偏负0.1‰ , 有可能是测试误差引起的。
从第66-1层至第76-2层, 植物叶片与整体的δ 13C值变化趋势相似, 而茎轴的δ 13C值在第68-2层至第76-2层与植物整体的变化趋势相似, 在第66-1层至第68-2层的变化趋势则相反(图 5)。可以推测, 相对于茎轴, 叶片δ 13C值变化趋势更接近植物整体δ 13C值的变化趋势, 说明植物整体的碳同位素变化主要是由叶片的碳同位素分馏效应决定。因此, 在了解植物整体的变化趋势时, 可以用叶片δ 13C值来进行研究。
由于植物叶片、茎轴与整体的δ 13C值差距最大为0.3‰ , 其余差距均为0.1‰ , 难以说明差异是否是由测试误差造成的, 故对3组数据进行Kruskal-Wallis H 检验。利用origin、SPSS这2种软件对3组数据进行显著性检验分析, 表明3组数据并不存在显著性差异(p> 0.05)。这与本次所测数据结果(图 5)相符, 即3组数据存在一定的差异, 但不显著。
在徐家冲组自下而上的垂向序列中, 植物δ 13C值总体呈现上升的趋势: 第33层、第57-3层、第79-7层植物δ 13C值分别为-28.1‰ 、-24.5‰ 、-23.2‰ , 上升约5‰ 。但到了徐家冲组的最上部, δ 13C值开始下降(图 2)。Wan(2012)认为, 自泥盆纪早期至中期植物化石的δ 13C值上升, 而中期至晚期则下降, 泥盆纪镰蕨属的δ 13C值在3个时期分别为-26.2‰ (早期)、-25.4‰ (中期)和-26.0‰ (晚期)。本次研究的镰蕨属位于泥盆纪早期, 碳同位素δ 13C值总体呈上升趋势, 中间略有波动, 与Wan(2012)的结论一致。
任昊佳(2005)此前已对徐家冲组第57旋回至第79旋回的镰蕨属进行过植物稳定碳同位素研究。在此基础上, 本次研究对徐家冲组的镰蕨属采集层位进行了补充, 增加了第33、64、65、76、84旋回的数据, 并增加了坡松冲组的数据, 其中第57、66、68、70、73与79旋回样品是2次研究都测试的, 而第69旋回样品是任昊佳(2005)报道但本次研究未测试的(图 2)。测试结果表明, 在第57旋回至第79旋回间, 笔者所测结果除中间波动略有区别外, 总体趋势与任昊佳(2005)相同, 均为上升。这进一步表明, 徐家冲组总体环境变化趋势是确定的, 但中间的变化过程多解, 需要进一步研究。
相对于徐家冲组, 坡松冲组时代跨度小, 更能表现出碳同位素值短时间内频繁的变化。研究区坡松冲组植物化石δ 13C值自下而上虽然经历下降、上升、小幅度上升和再下降的过程(图 3), 但总体并未有明显的变化趋势, 表明坡松冲组沉积时的环境相对稳定。由于坡松冲组植物化石的δ 13C值变化趋势与徐家冲组第64-68旋回的变化相似, 且两者所处的时代范围都为布拉格期晚期, 故推测该时期2个地区的环境变化较为接近。
植物的碳同位素组成是生理过程对大气进行“ 加工” 的结果, 植物是大气CO2浓度变化最灵敏的指示剂(冯净等, 2014)。低浓度的CO2是光合作用的限制因子, 植物细胞因CO2供应不足而来不及分馏重碳同位素, 从而使叶片δ 13C值偏正(O'Leary, 1981)。研究表明, 随着大气CO2浓度的上升, 植物δ 13C值逐渐偏负, 并呈显著负相关(Wagoner de Water et al., 1994; Feng and Epstein, 1995; Chen et al., 2006)。研究剖面中徐家冲组自第3大层至第7大层, 植物化石δ 13C值正偏达到5‰ 左右, 表明在这段时期大气CO2分压(pCO2)下降, 并可能导致了早泥盆世布拉格期晚期气候的变冷。至埃姆斯期, δ 13C值开始呈下降趋势(第8-9大层), 表明埃姆斯期早期可能有过短时间的气候变暖。
Farquhar等(1982, 1989)发现, C3植物叶片稳定碳同位素组成(δ 13C)与叶片细胞间CO2浓度和大气CO2浓度比值(CI/Ca)相关, 而CI/Ca值与大气二氧化碳到叶片细胞间的扩散以及CO2被光合羧化酶羧化的程度有关, 反映了净同化速率与气孔导度的相对量。
对于C3植物而言, CI/Ca值的计算公式(Farquhar et al., 1982)如下:
公式中,
依据测试所得的镰蕨属δ 13C值, 根据上述公式计算出大气的
| 表 2 云南省曲靖市徐家冲剖面下泥盆统徐家冲组镰蕨属的 |
Foster等(2017)对420 Ma以来的大气CO2浓度进行了分析, 发现泥盆纪早期的大气CO2浓度约为工业革命前的7倍, 大气CO2浓度从志留纪晚期开始呈现下降趋势, 在泥盆纪早期下降趋势相对平缓。徐家冲组镰蕨属的Ci/Ca值变化范围为0.57~0.59, 平均值为0.58, 相对稳定。因此, 这段时期内Ca的降低同时意味着Ci的降低, 表明镰蕨属在这段时间内的光合速率有所降低。同时也说明, 随着温度的降低, 镰蕨属的光合速率随之下降, 这从生理方面解释了镰蕨属Ci/Ca值保持恒定的机制。而恒定的Ci/Ca值则指示了镰蕨属对CO2变化比较敏感。坡松冲组的计算结果(表 3)与徐家冲组相似。
| 表 3 云南省文山州纸厂剖面下泥盆统坡松冲组工蕨属和始叶蕨属的 |
Chakraborty等(2011)研究印度白垩纪植物化石Ci/Ca值的变化时发现, 在合适的条件下, 植物往往会保持一个恒定或近似恒定的Ci/Ca值, 从而使它们的同化作用速率达到最优。当Ci/Ca值较高时(> 0.7), 反映了植物采用的是非节约型水分利用方式。在一些热带雨林物种中, 其Ci/Ca值均较高, 缺乏水分节约策略。本次分析的镰蕨属Ci/Ca平均值略低于0.7, 说明镰蕨属具有一定的水分节约策略, 从稳定碳同位素方面指示了曲靖市当时的气候类型不是热带雨林气候, 或者说湿润程度相对于热带雨林没有那么充足, 使得镰蕨属在利用水分时有一定的节约策略。根据柯本气候分类法(朱耿睿和李育, 2015)中的平均气温与降水量标准, 曲靖市当时的气候可能相当于现代的热带疏林草原气候或热带季风气候。
大气中13C进入植物体内并合成物质需要经过一系列物理和生物化学反应, 导致植物稳定碳同位素组成有所差异, 综合反映了植物叶片的光合作用和蒸腾作用(Korol et al., 1999)。因此, 植物叶片δ 13C值能反映与植物光合作用、蒸腾强度相关联的水分利用效率。水分利用效率(fWUE)是用以描述植物产量与消耗水量之间关系的, 对植物叶片来说: fWUE=光合作用A/蒸腾作用E。植物叶片δ 13C值已被许多研究验证为一种估计植物长期水分利用效率的可靠指标, 叶片的δ 13C值越偏正, 植物水分利用效率越高(Korol et al., 1999; Diefendorf et al., 2010; Kohn, 2010)。
C3植物碳同位素分馏公式的提出, 使得估算植物水分利用效率有了理论基础(Farquhar et al., 1982, 1989; 解三平等, 2006; Yan, 2009)。其中, Ci/Ca值是估算水分利用效率的重要数据, 数学关系式为:
Ci、Ca分别表示细胞间和大气中的CO2浓度, 1.6为水蒸气的气孔传导度与CO2的比值, fWUE的单位是μ molCO2/molH2O。
研究区早泥盆世镰蕨属、工蕨属、始叶蕨属的Ci/Ca平均值分别为0.58、0.57、0.57, 大气CO2浓度(Ca)采用Foster等(2017)提出的2000 μ mol/mol, 代入fWUE计算公式中, 得到fWUE分别为527.3 μ molCO2/molH2O、533.1 μ molCO2/molH2O、535.3 μ molCO2/molH2O。结果表明, 镰蕨属、工蕨属、始叶蕨属三者水分利用效率相近。
1)云南早泥盆世植物化石镰蕨属、工蕨属、始叶蕨属的δ 13C平均值分别为-24.5‰ 、-23.2‰ 、-22.7‰ , 均属于C3植物。
2)镰蕨属叶片的δ 13C平均值为-23.7‰ , 茎轴的δ 13C平均值为-23.6‰ , 叶片比茎轴偏负, 说明叶片比茎轴更能富集12C。镰蕨属δ 13C值变化范围在-29.0‰ ~-22.8‰ 之间, 并依据镰蕨属δ 13C值获得早泥盆世大气δ 13C值为-5.3‰ 。
3)依据云南下泥盆统徐家冲组与坡松冲组植物化石的δ 13C值纵向变化, 推测早泥盆世布拉格期晚期大气CO2浓度下降, 气候变冷, 而到了埃姆斯期早期, 大气CO2浓度上升, 气候变暖。
4)云南早泥盆世镰蕨属、工蕨属的Ci/Ca值在不同层位中保持相对稳定, 推测两者对大气CO2浓度的变化比较敏感。镰蕨属的Ci/Ca平均值略低于0.7, 指示了曲靖市当时的气候类型可能相当于现代的热带疏林草原气候或热带季风气候。
5)云南早泥盆世植物镰蕨属、工蕨属、始叶蕨属的水分利用效率相近。
致谢 感谢熊聪慧博士、刘乐博士、张迎迎博士等在碳同位素样品挑样中给予的支持, 感谢中国石油勘探开发研究院石油地质实验研究中心、核工业北京地质研究院在碳同位素检测方面给予的帮助。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|