






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
埃迪卡拉纪—寒武纪之交微生物岩特征对比及古海洋学意义: 以汉南—米仓山地区为例*
[邓嘉婷1  , 李飞
, 李飞1, 2 , 龚峤林1 , 李红1 , 易楚恒1 , 连承波1 ]
, 李飞, 龚峤林|
|

第一作者简介 邓嘉婷,女,1994年生,硕士研究生,主要从事碳酸盐沉积学方面研究。E-mail: dengjiating125@163.com。
寒武纪初期不仅发生了宏体生物大爆发,而且也出现了地质历史时期少见的蓝细菌鞘体大规模钙化事件。埃迪卡拉纪—寒武纪之交海水化学性质的转变对真核生物的演化起到了重要作用,但是这种转变对微生物岩发育特征以及蓝细菌钙化事件的产生有无影响,目前尚不明确。鉴于此,对华南上扬子北缘汉南—米仓山地区上埃迪卡拉统—寒武系第二统多个典型微生物岩发育剖面进行了系统野外调查和室内岩石学分析,结果表明:上埃迪卡拉统灯影组叠层石广泛发育,以平铺状、缓波状特征为主,而凝块石既可以呈补丁状分布于叠层石纹层间,又可以呈细小的凝絮状、粘结状特征构成厚层岩层;寒武系第二统仙女洞组叠层石丰度显著降低,以高大、坚硬的丘状隆起为特点,包括单独的凝块石丘,以及微生物与古杯的联合建丘。虽然寒武系第二统微生物岩的层状结构和凝块结构与埃迪卡拉系相比并无太大差异,但是寒武系微生物岩内部保存有大量的钙化微生物化石,已识别出附枝菌(Epiphyton)、肾形菌(Renalcis)和葛万菌(Girvanella)等多种类型。在收集、整理前人有关微生物岩特征和发育资料的基础上,本次研究初步整理出华南寒武系第二统微生物岩的时空分布特点,发现寒武纪第二世第三期是钙化微生物大量发育的一个时期,在随后的第四期达到一个小的高峰。对于此次蓝细菌钙化作用幕的启动机制,除前人提出的海水高钙离子浓度和蓝细菌体内二氧化碳浓缩机制等认识外,寒武纪早期海水性质的转变(方解石质原生矿物受成岩改造程度较低)、适度的陆源碎屑输入(黏土组分保护作用)也有利于钙化微生物结构的保存,应引起重视。
About the first author Deng Jia-Ting, born in 1994, is a master degree candidate of the Southwest Petroleum University. She is engaged in research of carbonate sedimentology. E-mail: dengjiating125@163.com.
Early Cambrian not only witnessed the explosion of animals,but also experienced a large-scale cyanobacterial calcification event that was rare in Precambrian. The transition of seawater chemistry from Ediacaran to Cambrian played a crucial role in the evolution of eukaryotes,but it is still unclear whether this transition also had impacts on the development and compositions of microbialites and on the start of cyanobacterial calcification event. This study conducted a systematic field investigation and petrological examination on the microbialite structures from the Upper Ediacaran to Cambrian Series 2 in the Hannan-Micangshan area that is located in the northern part of the Upper Yangtze area of the South China Block. The stromatolites of the Dengying Formation show regularly and irregularly laminated structures; the thrombolites are generally distributed between stromatolite layers or shown as individual,thick-bedded rocks,with fine-grained clotted and lump structures. In the the Xiannüdong Formation of Cambrian Series 2,the abundance of stromatolites is significantly reduced in the study area. The composition of microbialites is characterized by rigid,mound-like structures,including individual microbial mounds,and calcimicrobe-archaeocyathan bioherms. Although the laminated and clotted structures of microbialites of Cambiran Series 2 are similar to those of the Ediacaran,the Cambrian forms contain a large number of sheath-calcified microbial microfossils in microstructures,including Epiphyton, Renalcis, Girvanella, etc. Based on the literature review results of the characteristics and occurrences of the microbialites of Cambiran Series 2,this study has preliminarily sorted out the temporal and spatial distributions of the microbialites in the Cambrian Series 2 of South China. This study agrees with previous understandings that the high Ca2+ concentration in seawater and the reactive of CO2 concentration mechanism in cyanobacteria may have played a key role in the initiation of Cambrian cyanobacterial calcification event. In addition,the seawater chemical compositions in the Cambrian Series 2 that were changed from “aragonite-dolomite sea” to “calcite sea”,as well as the exceptional burial mechanism of clay coating within calcified microstructures may also be seen as two favorable conditions for the preservation of early Cambrian calcified microorganisms,which should be paid attention to.
开放科学(资源服务)标识码(OSID) 
微生物岩是指由底栖微生物群落通过粘结与捕获细粒沉积物以及通过与新陈代谢活动有关的诱导矿化过程在原地产生的碳酸盐, 经固结成岩作用后形成的生物沉积岩(Burne and Moore, 1987)。针对微生物岩前人开展了大量研究工作, 主要关注与微生物活动密切相关的沉积样式, 如叠层石(具纹层状结构)和凝块石(具花斑状凝块/团块结构)等(Riding, 2011a)。微生物岩在指示微生物群落构成、沉积环境、古海水化学性质等方面具有重要作用, 是研究古生态和古环境的重要载体(Riding, 2000; Webb and Kamber, 2000; Dupraz et al., 2009)。此外, 微生物岩的结构构造与孔隙发育程度关系密切, 其中窗格孔(洞)与格架孔(洞)可作为主要的储集空间, 并在四川盆地和塔里木盆地相关层位已取得重大油气发现(罗平等, 2013; 刘树根等, 2016; 沈安江等, 2016)。
微生物岩发育广泛, 从时间上可以追溯到早太古代, 并以中、新元古代及寒武纪和奥陶纪初期最为发育(Grotzinger and Knoll, 1999; Riding, 2000; Lee et al., 2015)。之后, 由于海水饱和度的显著降低和真核生物的兴盛, 微生物岩的发育丰度及规模大大降低, 其常依附于大型造礁生物, 或间歇性地出现在生物大灭绝事件之后, 例如晚泥盆世弗拉斯期—法门期(Webb, 1996; 龚一鸣和李保华, 2001)、二叠纪—三叠纪之交(王永标等, 2005)。微生物岩中的底栖微生物群落包括自养细菌(如蓝细菌)、异养细菌(如硫酸盐还原菌)和真菌类等, 其中能够进行光合作用的蓝细菌是一种主要的有机质生产者, 也是水体中溶解氧的重要来源之一(Nealson, 1997)。微生物并不直接参与微生物岩的形成, 在这个过程中起到关键作用的是微生物胞外聚合物(Extracellular polymeric substances, 简称EPS)(Pratt et al., 2000; Dupraz et al., 2004; Foster et al., 2009)、微生物膜(Biofilms)(Sutherland, 2001; 梅冥相等, 2019)以及微生物席(Microbial mats)(Visscher and Stolz, 2005; Baumgartner et al., 2006; Dupraz et al., 2009), 这三者在不同尺度下不仅能产生原地碳酸盐沉淀, 还能捕获、粘附外界沉积物, 构成微生物岩主体。EPS聚集于细胞外, 是由微生物制造出的一种黏性的、具保护作用的基质(Dupraz et al., 2004), 紧邻细胞外且与细胞形态贴合的管状或丛状EPS称为鞘体(Merz and Zankl, 1993; Riding, 2011b)。由于微生物自身新陈代谢活动或周围环境的变化, 可在鞘体内部或表面形成碳酸钙结晶, 其中在鞘体表面或外部形成的碳酸钙结晶是灰泥的重要来源, 而鞘体内部发生碳酸盐成核并保留下来的结构称为鞘体钙化蓝细菌(由于钙化鞘体形态与蓝细菌轮廓一致, 可简称为钙化蓝细菌), 例如葛万菌(图 1-A)(Pratt et al., 2000; Caron and Jackson, 2006)。另外, 还存在隐晶质的致密枝状和房室状微生物发生钙化的情况, 如附枝菌和肾形菌(Golubic et al., 2000; Riding, 2011a), 但由于现代蓝细菌中没有类似结构, 关于其成因和分类目前仍存在一些争议(Pratt et al., 1984; Chafetz and Guidry, 1999; Adachi et al., 2014), 故这些微生物也被称为疑似蓝细菌(Possible cyanobacteria)(Riding, 2001)。包括蓝细菌和疑似蓝细菌在内的微生物保存下来的钙质微体化石也被统称为钙化微生物(Calcified microorganisms)(Rowland and Shapiro, 2002)。
 | 图 1 蓝细菌鞘体钙化特征及机制 A—葛万菌, 样品来自南江沙滩剖面寒武系仙女洞组, 单偏光; B—蓝细菌鞘体钙化机制(参考Riding, 2011b)Fig.1 Photomicrograph showing calcified cyanobacteria and possible mechanism for formation of sheath-calcified cyanobacteria |
钙化微生物并不是普遍存在的, 例如现代海洋环境中鞘体发生钙化的蓝细菌非常少见, 这可能与当前海水中钙离子浓度较低有关(Arp et al., 2001)。地质历史时期中大规模钙化微生物出现在寒武纪、中—晚泥盆世以及中—晚三叠世等, 这种发生蓝细菌大规模钙化的现象也被称为“ 蓝细菌钙化事件” 或“ 蓝细菌钙化作用幕” (Riding, 1992; Chen and Lee, 2014)。虽然国内研究者已经认识到寒武纪存在钙化作用幕(梅冥相等, 2017), 但是对于最早的一批钙化蓝细菌的发育时间以及形成机制等, 目前研究还不太深入。另外, 蓝细菌钙化事件对微生物岩的宏观与微观特征是否会产生影响, 还有待深入探讨。基于此, 笔者以上扬子北缘汉南—米仓山地区为例, 通过对比分析上埃迪卡拉统和下寒武统微生物岩的类型、特征和发育情况, 明确了寒武纪钙化作用幕在中国的启动时间, 并对引发这次事件的原因进行了初步探讨。本次研究成果可为了解微生物岩结构对埃迪卡拉纪—寒武纪古海洋环境转变的响应提供参考, 亦可为认识钙化作用幕影响下的微生物成因储集岩类型和孔隙发育特征提供一定理论依据。
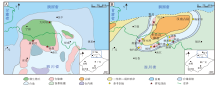
汉南—米仓山地区位于扬子板块北缘与秦岭造山带之间的过渡地带(Song et al., 2018)。该地区东邻大巴山推覆断褶带, 南接四川盆地, 西邻龙门山推覆构造带和松潘—甘孜造山带, 北为汉南推覆构造带(魏显贵等, 1997; 余谦等, 2011)。受裂陷活动和构造抬升作用影响, 新元古代晚期扬子板块北缘发育汉南古陆, 西南缘形成康滇古陆, 因而总体上扬子板块呈现出西高东低的沉积格局(魏显贵等, 1997; Zhu et al., 2003)。汉南古陆的发育受区域海平面升降影响, 范围常发生显著变化(甚至消失)。汉南古陆核心区域位于汉中—西乡一带, 米仓山地区(旺苍—南江地区)位于古陆核心区域西南方向, 为一受推覆构造作用影响的隆起区, 两者在新元古代晚期—古生代早期常共同影响区域古地理格局。除汉南古陆与米仓山隆起核心区外, 研究区内常见上埃迪卡拉统—寒武系第二统沉积记录(图 2)(魏显贵等, 1997; 曾楷等, 2020; 李红等, 2021; Ding et al., 2021)。
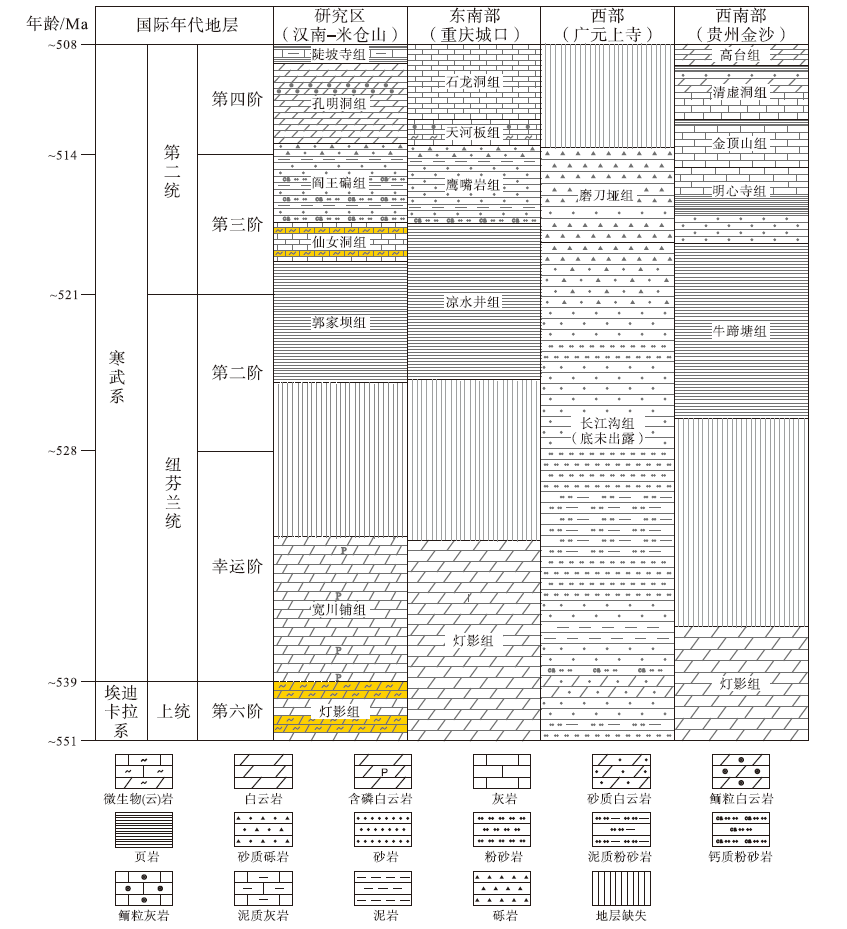 | 图 2 汉南—米仓山地区及其邻区上埃迪卡拉统至寒武系第二统地层划分与对比(据曹仁关, 1996; 张廷山等, 2008; 周传明等, 2019; 曾楷等, 2020; Ding et al., 2021; 有修改, 黄色阴影部分为本次研究主要关注的层位)Fig.2 Lithostratigraphic correlation of the Upper Ediacaran to Cambrian Series 2 in Hannan-Micangshan area and adjacent regions(modified from Cao, 1996; Zhang et al., 2008; Zhou et al., 2019; Zeng et al., 2020; Ding et al., 2021. The yellow shadow is the main focus of this study) |
研究区上埃迪卡拉统主要出露灯影组, 根据岩性特征和区域地层发育特点可进一步划分为4段, 其中一段主要为贫藻白云岩, 二段和四段为富藻白云岩(微生物白云岩), 三段主要为滨岸混积岩或碎屑岩(刘树根等, 2016)。受桐湾运动影响, 灯影组内部与顶部常发生小规模缺失, 与寒武系底部宽川铺组呈不整合接触(图 2)。受寒武纪初期(大致对应第二期)海侵作用的影响, 整个研究区大部分被海水淹没, 以粉砂质泥岩沉积(郭家坝组)为主, 之后在缓慢海退的过程中围绕汉南古陆和米仓山隆起发育浅水碳酸盐—碎屑组分混合沉积(仙女洞组), 包括砂质鲕粒滩、微生物丘和古杯丘等(龚峤林等, 2018; 曾楷等, 2020)。
本次研究的微生物岩主要采自灯影组(二段和四段)以及仙女洞组(图 2)。按照国内关于埃迪卡拉系和寒武系最新划分方案(周传明等, 2019; 朱茂炎等, 2019), 2套微生物岩分别属于埃迪卡拉系第六阶和寒武系第三阶(图 2)。本次研究野外实地调查的剖面包括勉县大河坝(灯影组和仙女洞组)、旺苍唐家河(灯影组和仙女洞组)、南江田垭(仙女洞组)、南江沙滩(仙女洞组)和南郑福成(仙女洞组), 参考剖面包括西乡杨家沟剖面(仙女洞组)(杨友运和叶俭, 1996)、南郑碑坝剖面(仙女洞组)(杨慧宁等, 2016)、南江杨坝(灯影组)(刘树根等, 2016)和柳湾剖面(仙女洞组)(刘学利, 2000; 张廷山等, 2002)、通江新潮剖面(仙女洞组)(Hicks and Rowland, 2009)以及旺苍青竹园和青木洞剖面(仙女洞组)(赵兵等, 1997)等(图 3)。
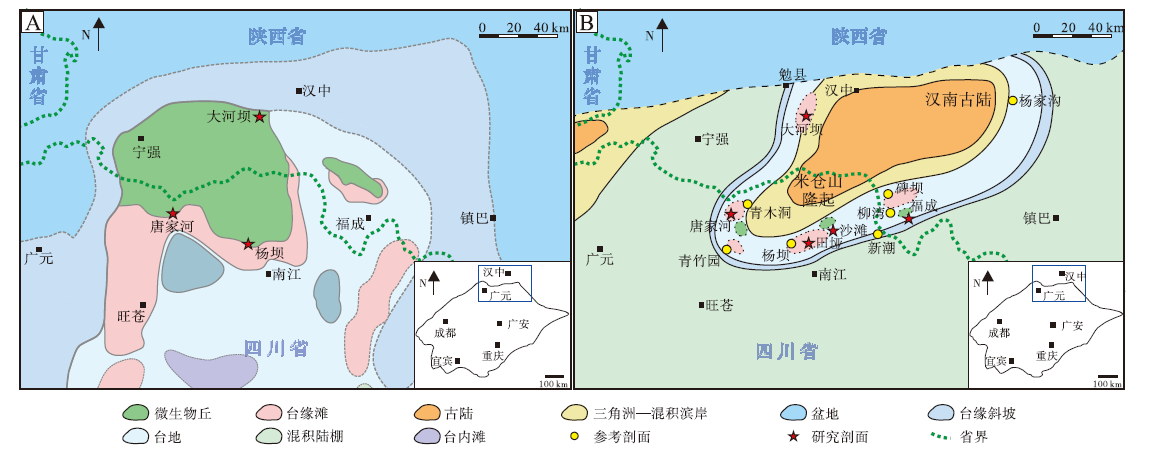 | 图 3 汉南—米仓山地区埃迪卡拉纪晚期(A)与寒武纪第二世第三期(B)古地理图及研究剖面位置(修改自李智武等, 2019; 曾楷等, 2020; 李红等, 2021)Fig.3 Location and palaeogeography of the Late Ediacaran (A) and Age 3 of Cambrian Series 2 (B) in Hannan-Micangshan area (modified from Li et al., 2019; Zeng et al., 2020; Li et al., 2021) |
本次研究通过野外详细调查, 对研究区灯影组与仙女洞组微生物岩宏观尺度和中观尺度下的特征进行系统描述和对比。室内对采集的岩石样品一部分切割抛光后利用高分辨率扫描仪成像后进行中观和微观尺度观察, 另一部分制作成薄片, 在偏光显微镜下(Lecia DM4P)进行岩石学微观尺度研究。微生物岩描述术语主要采用Shapiro和Awramik(2006)方案, 钙质微生物类型和主要特征参考Riding(1991)。另外, 本次研究还系统地收集了汉南—米仓山地区以及周缘浅水剖面寒武系第二统微生物岩发育记录的相关文献资料(图 3), 主要用于对微生物岩的类型、特征和发育情况等进行总结和对比。
4.1.1 叠层石
叠层石在研究区灯影组二段和四段均大量出露。在野外露头上, 叠层石形态较为单一, 以平波状、缓波状为主, 纹层大部分细密、平直, 少数具有波状起伏的特点(图 4-A)。未见柱状、锥状和穹隆状等典型的直立结构。叠层石发育厚度从几厘米至数米不等, 侧向延伸性好, 可达数米至数十米。中观尺度上, 叠层石具有毫米级纹层(平均厚度为2~4 mm), 呈平直状或断续状延伸, 少量纹层具有波状弯曲(图 4-B), 纹层之间常见平直的脉体和不规则形状的晶洞。
 | 图 4 汉南—米仓山地区上埃迪卡拉统灯影组叠层石形貌特征 A—叠层石宏观特征, 大河坝剖面; B—叠层石中观特征, 箭头所指为波状弯曲纹层, 唐家河剖面; C—叠层石微观特征, 其中暗色纹层占主导, 三角所指为窗格孔, 杨坝剖面, 单偏光Fig.4 Photographs showing stromatolites of the Upper Ediacaran Dengying Formation in Hannan-Micangshan area |
镜下观察发现, 纹层状叠层石内部主要为大量相互叠置的、厚度不均的暗色纹层。这类暗色纹层在微观尺度下多为深灰色—黑色隐晶质结构, 矿物类型主要为微晶白云石。大量球粒常与暗色纹层共生, 单层厚度在200~300 μm之间(图 4-C)。还存在少量浅色纹层分布于暗色纹层之间, 层厚大多在90~180 μm之间, 主要由细晶白云石组成。另外, 叠层石暗色纹层内部还可见鸟眼构造和窗格构造(图 4-C)。
4.1.2 凝块石
凝块石在研究区灯影组二段和四段发育也较为广泛, 但是发育规模总体较小(图 5)。以唐家河剖面为例, 在野外露头上凝块石主要呈厚层块状或者补丁状产出。块状凝块石厚度不等, 从数十厘米到数米均较为发育, 横向可延伸数米至数十米。补丁状凝块石主要产出于叠层石纹层之间, 以瘤状或斑状形式产出, 厚度一般不超过10 cm。在中观尺度上, 可明显观察到大量灰白色斑点状凝块结构, 形态不规则, 以椭圆状为主, 凝块直径总体较小, 以0.3~0.5 cm为主, 很少有超过1 cm的大凝块存在(图 5-A)。不同凝块断续相连, 或呈完全的分散状。凝块之间为大量皮壳状胶结物, 呈网状结构。此外, 凝块石内部可见大量被脉体充填、大小不一的孔洞(图 5-B), 该现象在灯影组二段地层中非常发育。
 | 图 5 汉南—米仓山地区上埃迪卡拉统灯影组凝块石形貌特征 A—凝块石宏观特征, 杨坝剖面; B—凝块石光面标本, 唐家河剖面; C, D—凝块石镜下照片, 可见凝絮状球粒构成的凝块结构, 唐家河剖面, C为单偏光, D为正交光Fig.5 Photographs showing characteristics of clotted structures in thrombolites of the Upper Ediacaran Dengying Formationin Hannan-Micangshan area |
偏光显微镜下观察发现, 凝块结构是由大量致密、隐晶质的深灰色—黑色凝絮状球粒集合体组成(图 5-C, 5-D), 呈不规则状分布。紧密相连的凝块结构和分散状凝块结构均较为发育。不同凝块结构可以相互串连构成格架状, 或在横向上粘结构成类似于层状的结构。单个凝块边缘较为规则, 呈港湾状或皮壳状, 内部为大量凝絮状集合粒, 少量为微亮晶和亮晶胶结物(图 5-C)。凝块结构之间的孔隙常被亮晶胶结物充填, 胶结物多具有世代性, 颗粒边缘以纤状—叶片状等厚环边胶结物为主, 内部主要为粒状、亮晶胶结物。
4.2.1 叠层石
叠层石在仙女洞组出露较差, 野外仅在南郑福成剖面有少量发现。宏观上, 叠层石以穹隆状为主, 高度在50 cm左右, 宽度在40~60 cm之间, 分布于大型凝块石丘的顶部。中观尺度上, 叠层石纹层较为细密、光滑, 呈穹顶状向上凸起, 以较大幅度的顶突特征为代表(图 6-A, 6-C)。
显微镜下观察发现, 叠层石结构多由次一级、小的叠层石紧密排列堆积成层, 不同层之间亦相互叠置, 构成大的叠层结构(图 6-B, 6-D)。单个小的叠层石具有中间凸、两边凹的穹顶特征, 主要由致密的微晶状纹层构成, 少量为微亮晶方解石构成的明色层(图 6-B)。小的叠层石之间为陆源粉砂和黏土组分(图 6-B)。暗色层内可以识别出少量丝状葛万菌(图 6-B小图), 顺纹层发育, 其他类型的钙质微生物非常少见。
 | 图 6 汉南—米仓山地区寒武系第二统仙女洞组叠层石特征和素描图 A—叠层石野外特征, 具有穹顶短柱状形态(黄色线条所示); B—叠层石镜下特征(单偏光), 具有上拱特征, 纹层内可见葛万菌(小图); C, D—分别为图A和B的素描图, 进一步展示了纹层内部结构。野外和镜下图片均来自南郑福成剖面Fig.6 Photographs and corresponding line drawings showing laminar structures of stromatolites of the Xiannü dong Formation of Cambrian Series 2 in Hannan-Micangshan area |
4.2.2 凝块石
寒武系仙女洞组微生物岩以凝块石为主, 其发育特征在福成、沙滩和唐家河等剖面较为典型。宏观上, 凝块石呈大型丘状产出, 宽度1~5 m, 高度一般在0.5~2 m之间。凝块石丘的基底一般为鲕粒岩或钙质细砂岩, 丘顶盖层常见鲕粒岩或钙质砂岩, 丘体一般侧向叠置发育, 丘和丘之间常为含砾屑砂岩或砂质鲕粒岩(图 7)。
 | 图 7 汉南—米仓山地区寒武系第二统仙女洞组钙质微生物—古杯丘宏观特征(大河坝剖面, 地质锤长度为1 m)Fig.7 Field photograph showing calcimicrobial-archaeocyathan mounds of the Xiannü dong Formation of Cambrian Series 2 in Hannan-Micangshan area (Daheba outcrop, and the hammer’ s length is 1 m) |
此外, 部分凝块石丘内部还可见古杯化石和鲕粒等。中观尺度上, 凝块直径以0.5~1 cm为主, 以不规则斑点或团块状产出(图 8), 形态多样, 产出密集或呈分散状。可见凝块结构在纵向上相连, 构成枝状结构(图 8-C), 或者相互连接形成网状结构; 凝块结构之间常见陆源碎屑(例如大河坝剖面, 图 8-B)。此外, 古杯动物常与凝块结构共生, 可见古杯化石碎屑被致密凝块结构包裹构成粗壮格架(厘米级), 格架间常为陆源粉砂和泥质组分。
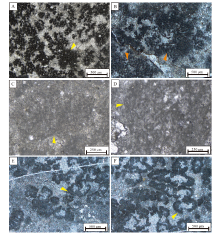
 | 图 8 汉南—米仓山地区寒武系第二统仙女洞组凝块石中观、微观特征 A, B, C—凝块结构野外特征; D—附枝菌主导的凝块, 单偏光; E—葛万菌主导的凝块结构, 单偏光, 详细特征见图 9-C和9-D; F—肾形菌主导的凝块结构, 单偏光。其中A来自福成剖面, B来自大河坝剖面, C和E来自沙滩剖面, D和F来自唐家河剖面Fig.8 Field photographs and photomicrographs of clotted structures of the Xiannü dong Formation of Cambrian Series 2 in Hannan-Micangshan area |
镜下可见凝块结构主要由附枝菌和肾形菌等钙质微生物以及相邻的微晶成分构成(图 8-D至8-F), 部分还黏结灰泥和少量细粒陆源碎屑组分, 偶见鲕粒和古杯化石碎片。凝块的形状与钙质微生物的生长方向和形态有关。
附枝菌组成的凝块(图 8-D; 图 9-A, 9-B)整体呈灌木丛状: 纵切面上可见附枝菌呈细长分枝状, 长度在130~400 μm之间, 菌体呈束状(图 9-A)或扇状(图 9-B)展开; 横截面上附枝菌呈聚集分布的圆斑点, 直径在30~50 μm之间。葛万菌组成的凝块(图 8-E; 图 9-C, 9-D)形状大小不规则, 并由葛万菌聚集数量、缠绕程度以及所粘结的泥晶含量所决定。在镜下可观察到发育良好的长体管状构造, 菌丝体呈单管弯曲, 由深灰色管壁和中间明亮的管芯组成, 长度不等, 最长可达1 mm, 管体不分叉, 直径均匀, 大约为14~20 μm; 菌丝个体相互缠绕和穿插, 具有明显的泥晶外壁。由肾形菌构成的凝块, 是由单个或串连的集合体组成(图 8-F; 图 9-E, 9-F), 其中单个肾形菌菌体由囊状或新月形房室状结构构成, 外壁由暗色泥晶组成, 内部为空腔, 直径约300 μm。
 | 图 9 汉南—米仓山地区寒武系第二统仙女洞组常见钙化微生物类型和特征 A, B—附枝菌(唐家河剖面), 分枝状特征明显, 黄色箭头指示簇状附枝菌呈圆斑状分布(横截面), 橙色箭头指示附枝菌菌群呈中空的发散状分布, 唐家河剖面, 单偏光; C, D—葛万菌, 属丝状蓝细菌类(黄色箭头所示), 沙滩剖面, 单偏光; E, F—肾形菌, 房室状特征明显(黄色箭头所示), 唐家河, 单偏光Fig.9 Photomicrographs showing microstructures of calcified microbial microfossils of the Xiannü dong Formation of Cambrian Series 2 in Hannan-Micangshan area |
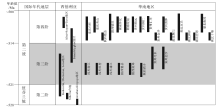
自太古宙以来, 微生物岩在沉积记录中分布非常广泛, 埃迪卡拉纪和寒武纪是微生物岩最为发育的几个时期之一(Riding, 2000)。尽管埃迪卡拉纪和寒武纪时代相邻, 但是2个时期的微生物岩在宏观面貌和微观结构上却存在一定差异(图 4至图 8)。其中上埃迪卡拉统微生物岩以叠层石为主, 在宏观尺度下多以厚层状、板状特征出现, 主要形成于开阔陆架环境, 潮坪环境下柱状和穹窿状叠层石也较为发育(Thorie et al., 2018); 凝块石相对少见, 以补丁状、层状结构为主, 常与叠层石共生(图 5)。而在寒武纪早期, 开阔陆架环境下的叠层石发育规模明显缩小, 凝块石占据主导地位(Rowland and Shapiro, 2002; Hicks and Rowland, 2009), 可见叠层石形成于大型凝块石建造的顶部, 凝块石的规模与上埃迪卡拉统相比显著增大(米级以上), 且形态上以丘状建隆占主导(图 8)。中观尺度上, 埃迪卡拉系和寒武系微生物岩最明显的区别是: 寒武系凝块石内部出现树枝状(国内外也有一些学者把这种类型称为树枝石)和扇状结构(图 8-A, 8-C), 这些特征可以指示凝块石的生长方向; 而相关特征在研究区上埃迪卡拉统灯影组并未发现, 国内其他地区报道也非常少见。微观尺度上, 虽然埃迪卡拉系和寒武系微生物岩的部分特征相似, 但是寒武系第二统保存的层状和凝块结构内部均可见大量钙化微生物化石, 而这在埃迪卡拉系微生物岩中是非常少见的, 埃迪卡拉系微生物岩中的蓝细菌更倾向于以磷酸盐化、硅化、黄铁矿化等特异埋藏形式保存(王伟等, 2013; Min et al., 2020), 而非纯粹的钙化形式保存。需要指出的是, 并不是所有的寒武系微生物岩内部结构中均可见钙化微生物的产出, 但与埃迪卡拉系相比, 在寒武系第二统钙化微生物产出明显增多(图 10)。
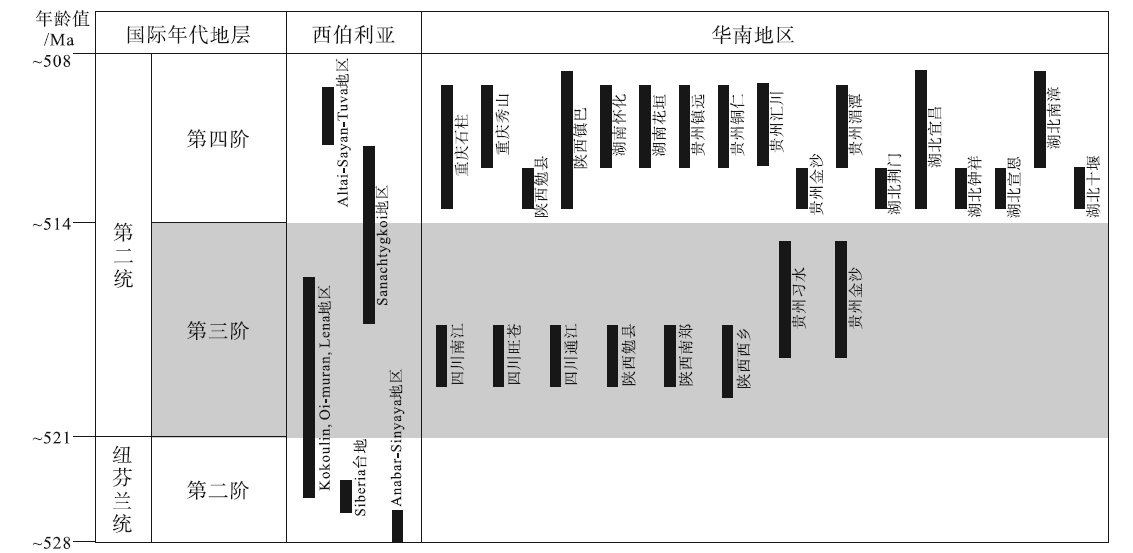 | 图 10 华南地区寒武系具钙化微生物特征的微生物岩分布简图及其与西伯利亚对比(西伯利亚资料引自Rowland和Shapiro, 2002)Fig.10 A brief chart showing spatial and temporal distributions of calcified microbes in the Cambrian microbialites of South China, and a comparison with Siberia Block (the Siberia data cited from Rowland and Shapiro, 2002) |
虽然在中国埃迪卡拉系和寒武系底部地层中有大量关于蓝细菌发育的报道(曹仁关, 1980, 1996; 侯奎和陈延成, 1990; 钱逸等, 2007), 但是这些蓝细菌绝大多数保存在磷块岩或硅质岩中, 而非保存在碳酸盐组构中, 故保存方式可能与特异性埋藏条件有关(Muscente et al., 2017)。基于统计资料来看, 蓝细菌钙化事件可能始于寒武纪纽芬兰世(图 10), 并在西伯利亚台地(Tommotian期)广泛分布的微生物岩建造中已经开始大量发育钙化微生物(Luchinina et al., 2013)。华南地区钙化微生物广泛分布在寒武纪第二世的川北—陕南、贵州、湖北、重庆、湖南等地(杨爱华, 2005; 张廷山等, 2005; 梅冥相, 2007; Hicks and Rowland, 2009; 王建坡等, 2014; Tang et al., 2019), 例如第二世第三期汉南—米仓山地区已经开始发育大量含有钙化微生物的微生物岩(图 10), 以凝块石丘和凝块石—古杯丘的建造形式出现(图 7)(刘学利, 2000; Rowland and Shapiro, 2002; 穆西南和袁训来, 2003; 张廷山等, 2005; Hicks and Rowland, 2009; 王建坡等, 2014; Zhang et al., 2017; Tang et al., 2019)。这说明华南地区寒武纪钙化微生物的首现可能晚于西伯利亚台地(图 10), 直到第二世第四期时, 其类型、丰度和分布范围才达到寒武纪以来1个小的高峰期(图 10)。另外, 在华北和塔里木地区第二世也都有钙化微生物发育的报道(宋金民等, 2014; Lee et al., 2016), 这也与全球其他板块(如西伯利亚、蒙古、澳大利亚、北美等)寒武纪早期钙化微生物岩发育趋势(Rowland and Shapiro, 2002; Adachi et al., 2014)具有较好的相似性。因此, 笔者初步认为华南寒武纪第二世第三期微生物岩中钙化微生物的广泛出现可被视为中国寒武纪“ 蓝细菌钙化作用” 的勃发期。
5.2.1 海水钙离子浓度与二氧化碳浓缩机制(CCM)
蓝细菌鞘体钙化的记录最早出现在中元古代, 在古生代特别是在早古生代, 具有钙化鞘体的微生物较为常见(图 11)。
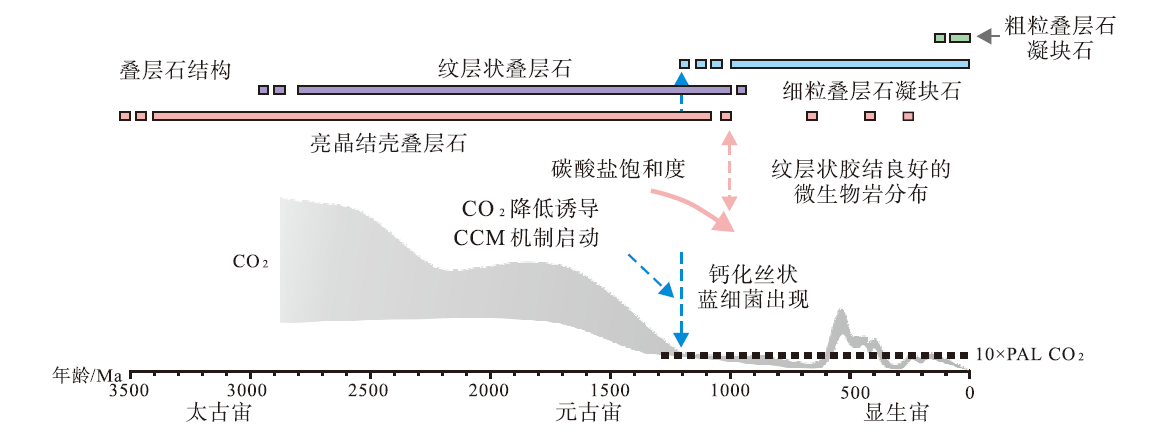 | 图 11 地质历史时期微生物岩发育特征及与二氧化碳浓度机制的关系(Riding, 2011b; PAL表示现代大气水平)Fig.11 Development of different types of microbialites in Earth’ s history and relationship between occurrences of calcified microorganisms and CO2 concentrating mechanism(after Riding, 2011b. PAL representing the modern atmospheric level) |
一些研究者认为早古生代海洋环境中钙化蓝细菌的繁盛与海水中具有较高的钙离子浓度有关, 且钙离子浓度的显著下降与钙化蓝细菌的消失具有明显的相关性(Arp et al., 2001)。Riding(2006)进一步的工作认为, 除了较高的钙离子浓度可以促进鞘体钙化外, 蓝细菌新陈代谢过程中二氧化碳浓缩机制(CCM)的运作可能也起到了重要作用。CCM机制可以调节二氧化碳与蓝细菌内部的交换效率, 进而影响微环境碱度和碳酸钙沉淀速率(图 1; 图 11)(Riding, 2006; Price et al., 2008)。当CO2浓度降低至约10倍现代大气水平时, 诱发CCM工作(Konhauser and Riding, 2012), 形成较为普遍的钙化蓝细菌化石(Riding, 2011b)。寒武纪初期大气CO2浓度显著增加, 可能超过10倍现代大气水平(图 1), 此时蓝细菌钙化鞘体的产生与CCM机制的运转之间关系可能并不密切。
5.2.2 古海洋化学性质
钙化微生物的形成和保存与它们原生矿物类型存在重要关系。碳酸盐矿物类型主要有文石、低镁方解石和高镁方解石(> 4 mol % MgCO3)3种主要类型(Morse et al., 2007; 李飞等, 2015)。蓝细菌新陈代谢过程中, 钙离子的固定主要是通过对海水来源流体中钙离子的吸附实现的, 而非调控菌体微环境的Mg/Ca值, 因而钙化蓝细菌的矿物构成很可能主要受海水Mg/Ca值的调控(Ries et al., 2008)。
由于文石是亚稳定矿物, 在成岩过程中会向更为稳定的低镁方解石矿物转变(Morse et al., 2007; 李飞等, 2015), 因而原生矿物为文石的钙质微生物组构很可能会发生溶解和重结晶, 使钙化微生物的主要识别特征(如管、壁结构等)难以判别。高镁方解石也属于亚稳定矿物, 成岩过程中同样会被转化为低镁方解石(Morse et al., 2007), 但是Mg含量低于8 mol % MgCO3的高镁方解石在热力学上相对于文石更为稳定(Berner, 1975), 因此在原始矿物结构保存程度上也优于文石。由低镁方解石构成的钙质微生物结构受成岩改造的影响较小, 因而其原始的精细结构在相对封闭的情况下更容易保存下来(Sandberg, 1983)。
寒武纪早期正处于埃迪卡拉纪“ 文石—白云石海” 向早古生代“ 方解石海” 转变的过渡期(Sandberg, 1983)。成冰纪和埃迪卡拉纪的海水以广泛的缺氧和极高的海水Mg/Ca值为特征, 被称为“ 文石—白云石海” , 并在埃迪卡拉纪末期快速转变为典型的“ 文石海” (Hood et al., 2011; Wood et al., 2017)。对流体包裹体和非骨骼碳酸盐矿物(鲕粒和海相胶结物)的研究表明(Wood and Zhuravlev, 2012; Zhuravlev et al., 2015; Wood et al., 2017), 寒武纪第三期以方解石质非骨骼矿物和方解石主导的生物矿物初现为特征(Porter, 2007), 应属于“ 方解石海” 时期。因此, 在寒武系第三阶保存情况良好的钙化蓝细菌的大量出现, 与海水性质有利于方解石质微生物结构的保存可能存在密切关系。
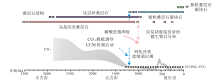
5.2.3 特异性埋藏
寒武纪早期钙质微生物的保存可能也与铝硅酸盐的保护机制存在一定关联。从埃迪卡拉系灯影组与寒武系仙女洞组微生物岩镜下特征的对比来看, 灯影组微生物岩内部的纹层状结构与凝块状结构保存程度良好, 这与当时准同生期拟晶白云石化有关(Zempolich et al., 1988)。由于白云石矿物的热力学稳定性较高, 早期白云石化对微生物原始结构的保存起到了促进作用(降低了后期成岩改造的影响程度)(Morse et al., 2007)。但是早期白云石化发生的时间稍晚于文石矿物的溶解(Berner, 1975), 因而即使存在早期白云石化作用, 原始文石质微生物组构也较难保存下来。硅化和磷酸盐化最早可以在紧邻沉积物—水界面之下的环境中发生(图 12), 因而早期磷酸盐化或硅化的蓝细菌结构能够在埃迪卡拉纪较为广泛地出现(王伟等, 2013; Muscente et al., 2015)。同时, 埃迪卡拉纪海水中较高的P离子和Si离子浓度(Tarhan et al., 2016; Lenton and Daines, 2018), 以及早期成岩过程中有机质和大量陆源碎屑组分分别释放的磷酸盐和硅酸盐, 对蓝细菌的磷酸盐化和硅化保存起到了促进作用(王伟等, 2013)。
 | 图 12 特异性埋藏路径理想模式图(修改自Muscente et al., 2017)Fig.12 A cartoon showing ideal model of exceptional taphonomic pathways in shallow-burial depositional succession (modified from Muscente et al., 2017) |
寒武纪以来, 海水中镁离子含量显著降低(Brennan et al., 2004), 形成拟晶白云石的机制不复存在, 大量钙质微生物化石的保存也与磷酸盐化和硅化没有直接关系。但是, 通过对汉南—米仓山地区钙质微生物的镜下观察, 笔者发现附枝菌、葛万菌与肾形菌的钙化结构中含有大量的自生黏土矿物, 以及较高含量的黏土级陆源碎屑组分。由于微生物组构本身具有粘结捕获周围环境细粒陆源碎屑组分的能力(Burne and Moore, 1987), 因而在近岸浑浊水环境下, 吸附的细粒陆源碎屑组分(主要为黏土)通过在蓝细菌表面形成薄膜(Martin et al., 2004; 王伟等, 2013), 或者在早期成岩过程中扩大抑制碳酸盐溶解的Fe/Mn还原带范围(Su et al., 2020; 苏成鹏等, 2021), 进而促进原始钙化微生物结构的保存。尽管如此, 特异性埋藏对于钙化微生物的大量出现可能只是起到次要作用。蓝细菌钙化鞘体的保存从依赖其他矿化方式到直接钙化, 可能主要与蓝细菌钙化能力的增强有关。
通过系统对比汉南—米仓山地区上埃迪卡拉统与寒武系第二统微生物岩宏、微观特征与内部结构, 结合华南地区同时期其他剖面资料, 发现埃迪卡拉纪晚期与寒武纪早期微生物岩相比, 在形态上出现以层状叠层石和凝块石占主导向具有明显丘状特征的凝块石占主导的转变。同时, 凝块石与叠层石从埃迪卡拉系的共生或邻近产出, 转变为寒武系的相对独立。凝块结构从大量细小凝块的网状分布转变成寒武系典型的直立枝状或扇状特征。2个时期微生物岩最大的不同体现在微观结构上, 即寒武系第二统纹层结构和凝块结构内部出现了大量钙化的微生物且类型多样。现有资料显示华南寒武系第二统第三阶开始大量出现钙化微生物记录, 以葛万菌、附枝菌和肾形菌为主, 在第四阶具钙化微生物特征的微生物岩在华南不同地区广泛出现。华南寒武纪钙化微生物的发育晚于全球范围内寒武纪最早记录, 但发展趋势与全球范围内其他板块特征基本符合。对于这次寒武纪早期蓝细菌钙化事件的产生, 目前国际上认为与当时海水钙离子浓度的增加以及蓝细菌二氧化碳浓缩机制的激活有关。本次研究认为, 寒武纪早期海水化学成分的转化可能有利于方解石质蓝细菌钙化结构的保存。另外, 研究区寒武纪第二世第三期陆源细粒物质(以黏土为主)的大量输入, 其中有相当一部分被微生物结构粘结、捕获, 这些陆源黏土物质通过形成黏土膜或碱性成岩流体环境, 减轻了原始组构被成岩改造的影响。从目前看, 寒武纪蓝细菌钙化作用幕的启动机制还有很多问题值得探讨, 有待进一步研究。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|