





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
川西北下二叠统栖霞组微生物丘的发现及地质意义*
[全子婷1, 2, 3  , 谭秀成
, 谭秀成1, 2, 3 , 张本健4 , 唐浩1, 2, 3 , 罗冰4, 5 , 杨迅6 , 张亚4 , 肖笛1, 2, 3 , 汤艳玲1, 2, 3 ]
, 谭秀成, 张本健|
|

通讯作者简介 谭秀成,男,1970年生,教授,博士生导师,主要从事沉积学与储层地质学研究工作。E-mail:tanxiucheng70@163.com。
第一作者简介 全子婷,女,1994年生,硕士研究生,主要从事沉积学研究工作。E-mail:qzt_0904@qq.com。
通过野外露头与钻井剖面的室内外分析,确认在四川盆地西北部(川西北)下二叠统栖霞组中发育有微生物丘,它们主要由凝块灰(云)岩、叠层灰(云)岩和微生物粘结颗粒灰(云)岩等组成。这些微生物碳酸盐岩发育较为典型的凝块、叠层、窗格、粘结等组构。微生物丘大小不一,实测高度一般为几十厘米至几米,宽度通常变化于几米至几十米之间,具有底平顶凸的典型丘形外貌,以发育向上变浅的沉积序列为特征,一般由丘基、丘核、丘盖 3个微相组成,也可与颗粒滩共同构成微生物丘滩复合体。基于区域古地理背景和微生物丘特征的剖析,认为川西北地区栖霞期沉积环境总体受限,推测为半局限—局限台地环境,水深较浅,能量普遍不高;海平面频繁的相对升降变化和微生物丘的侧向迁移叠置,导致发育于缓坡背景下的碳酸盐岩台地极易受限,引起早期沉积物发生与丘滩发育密切相关的准同生期白云石化作用。因此,微生物丘滩复合体是栖霞组白云岩储集层发育的物质基础,台缘坡折带、台内缓坡折带和高地是栖霞组白云岩储集层发育的有利区带,这对寻找规模性层位不稳定的带状白云岩储集层具有重要的指导作用,并将大大拓展栖霞组白云岩储集层的勘探领域。
, Tan Xiu-Cheng, Zhang Ben-JianAbout the corresponding author Tan Xiu-Cheng,born in 1970,professor and Ph.D. advisor,is mainly engaged in the research on sedimentology and reservoir geology. E-mail:tanxiucheng70@163.com.
About the first author Quan Zi-Ting,born in 1994,is a master degree candidate of Southwest Petroleum University. She is mainly engaged in the reasearch on carbonate sedimentology. E-mail:qzt_0904@qq.com.
Through analysis of outcrops and drilling profile data in the field and laboratory,microbial mounds were found in the Lower Permian Qixia Formation in the northwestern Sichuan Basin. The microbial mound is mainly composed of thrombolites,stromatolites,and microbial bindstones. These microbial carbonates are characterized by development of typical clots,stromatolitic,fenestral,and microbial bonding fabrics. Microbial mounds of the Qixia Formation are typical mound-shaped with flat bottom and convex-up top,and various in size. They range from tens of centimeters to several meters in height and from several meters to tens of meters in width. The mounds are characterized by shallowing-upward sequences,which are generally composed of mound base,core and cap,and also may form a mound-bank complex with carbonate shoals. Analysis of regional paleogeographic background and the characteristics of microbial mounds suggests that the Qixia period in northwestern Sichuan was deposited in a semi-restricted platform,with shallow-water and the relatively low-energy conditions. The high-frequency sea-level change,lateral migration,and superposition of microbial mounds lead to the restricted environment on carbonate platform with ramp background,which results in penecontemporaneous dolomitization in earlier sediments. Microbial mound-bank complexes provide basis for development of the dolomite reservoirs in the Qixia Formation. The slope-break belt of platform margin,gentle slope break belt,and local highland inside carbonate platform are favorable zones for the development of the dolomite reservoirs in the Qixia Formation. This understanding plays an important guiding role in finding banded dolomite reservoirs with unstable scale horizons,and will greatly expand the exploration field of dolomite reservoirs in the Qixia Formation.
开放科学(资源服务)标识码(OSID) 
微生物岩是一种由微生物群落通过捕获与粘结碎屑沉积物, 再通过物理作用或化学作用沉淀而在原地形成的沉积岩(Riding, 1991)。近年来, 有关微生物岩和微生物丘的研究方兴未艾(王永标等, 2005; 梅冥相, 2007; 王红梅等, 2013; 郝雁等, 2018; 吴亚生等, 2018, 2021; 胡安平等, 2021), 但主要集中在前寒武纪— 寒武纪、泥盆纪、石炭纪及三叠纪等。对于早二叠世的微生物岩, 目前罕有报道, 仅Nakazawa等(2015)报道了早二叠世泛古洋中的秋叶环礁存在由自养和异养微生物群落共同建造的微生物礁, 刘治成等(2015)报道了四川盆地华蓥山溪口剖面栖霞组存在以灰泥为支撑结构、由多种菌类微生物参与建造而成的微生物灰泥丘。有限的研究成果表明, 早二叠世具有微生物群落繁荣的古地理背景。
已有的研究表明, 栖霞组地层时代为早二叠世空谷期中晚期(沈树忠等, 2019; 刘正元等, 2020), 华南栖霞组以广泛发育由似球粒和包粒组成的中低能滩相沉积为特征(马志鑫等, 2011), 典型特征是亮晶和泥晶胶结以一定比例共存, 明显有别于华南地区其他地质时期典型的由高能亮晶颗粒岩构成的滩相沉积。马志鑫等(2011)通过对广元上寺剖面栖霞组的研究, 认为川西北地区栖霞组似球粒包括粪球粒、巴哈马似球粒、微生物似球粒3种类型, 且以泥晶化成因的巴哈马似球粒发育为特征, 并将其解释为浅水高能生物滩沉积或浅缓坡沉积。然而, 更多的学者将其解释为开阔环境沉积(黄先平等, 2004; 胡明毅等, 2010; 杨光等, 2015; 周进高等, 2016; 厚刚福等, 2017), 但马永生等(2009)提出了川西地区为局限台地相、川西北地区为开阔台地相这一观点, 而李明隆等(2020)和芦飞凡等(2020)等通过对川西北地区栖霞组白云岩宏微观特征及地球化学特征的研究, 认为该地区存在局限重卤水渗透回流白云石化, 亦暗示区内栖霞期古环境可能受限。
近期研究发现, 在川西北地区下二叠统栖霞组发现叠层(灰)云岩、凝块灰(云)岩、微生物粘结砂屑灰(云)岩等微生物岩, 且存在与微生物丘向上变浅序列有关的丘滩相白云岩, 暗示区内栖霞期与微生物丘叠置迁移相关的古环境可能受限。鉴于此, 文中以四川盆地西北部下二叠统栖霞组为研究对象, 通过露头和钻井岩心的宏观与微观研究, 发现了研究区栖霞组确实发育有微生物丘, 并着重从岩石类型、沉积特征和沉积序列组合的角度探讨早二叠世微生物丘特征及古环境。该研究结果不仅对重新认识早二叠世栖霞期古环境有借鉴意义, 也因明确了白云岩与微生物丘的关系, 从而对于层位不稳定的带状白云岩储集层勘探具有重要的指导作用。
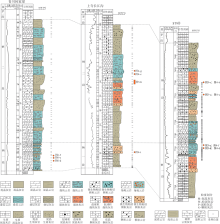
研究区位于四川盆地西北部川西山前凹陷带、龙门山断褶带以及米仓山隆起带的过渡区(宋文海和庞家黎, 1994; 陈洪德等, 2011), 包括广元、剑阁、旺苍、苍溪等地, 总面积约5000km2(图 1-a)。
 | 图 1 川西北区域地质略图及下二叠统栖霞阶地层柱状图 a— 研究区地理位置和构造位置图(断裂发育情况据杨光等, 2015; 杨跃明等, 2018), 以及本次研究所利用的露头剖面与钻井剖面位置; b— 川西北下二叠统栖霞阶地层柱状剖面示意图(修改自马永生等, 2009)Fig.1 Geological sketch of northwestern Sichuan Basin and stratigraphic column of the Lower Permian Qixia Stage |
石炭纪末期, 云南运动导致研究区地层普遍抬升暴露, 遭受差异剥蚀, 这造成二叠系沉积前的古地理格局转变为局部地貌隆坳起伏的缓坡环境(宋文海, 1987)。随后, 栖霞阶初期广泛的海侵使二叠系覆盖在石炭系、泥盆系、志留系等不同时代地层的不整合面之上(陈宗清, 2007)。中、下二叠统自下而上依次发育梁山组、栖霞组和茅口组(图 1-b)。梁山组为一套海侵初期滨岸— 沼泽相砂泥岩夹煤层沉积(四川省地质矿产局, 1991; 童崇光, 1992; 郭正吾等, 1996; 辜学达等, 1997; 魏国齐等, 2011; 赵宗举等, 2012; 黄涵宇等, 2017)。受后期持续海侵的影响, 栖霞组演变为碳酸盐岩台地沉积, 下部主要发育泥晶灰岩、生物泥晶灰岩, 局地可见斑状、纹层状云质灰岩。已有的研究成果表明, 栖霞组沉积中晚期为早二叠世最重要的成滩期, 以发育似球粒颗粒灰(云)岩、亮晶颗粒灰岩等相对高能沉积物为特征(田景春等, 2014; 娄雪, 2017; 关新等, 2018; 胡安平等, 2018; 白晓亮等, 2019)。据最新的钻井岩心和露头剖面观察, 栖霞组还发育有凝块灰(云)岩和叠层灰(云)岩等微生物岩类(图 2)。栖霞组沉积末期, 相对海平面下降, 导致栖二段颗粒滩大面积暴露, 遭受大气淡水淋滤溶蚀(王海真等, 2013; Xiao et al., 2019)。茅口组沉积初期发生持续海侵, 导致较为低能的灰岩与泥灰岩覆盖在下伏栖霞组之上(苏成鹏, 2017; 罗进雄等, 2019; Su et al., 2020)。
 | 图 2 川西北栖霞组岩性结构柱状剖面及横向对比Fig.2 Lithologic structural columns of the Qixia Formation and their lateral correlation in northwestern Sichuan Basin |
笔者采用野外宏观观察与室内显微镜下精准鉴定相结合的方法, 鉴别组成微生物丘的主要岩石类型及其岩石学特征, 而对于由白云石化作用形成的、但保存有原岩组构幻影的晶粒白云岩, 则采用单偏光显微镜薄片下加垫70g 的A4纸进行原岩组构恢复。在川西北栖霞组内, 笔者首次识别出叠层灰(云)岩、凝块灰(云)岩、微生物粘结颗粒灰(云)岩等典型的微生物岩类型。
叠层灰(云)岩是以蓝菌(蓝藻)为主的微生物在生长和新陈代谢活动过程中, 粘附和沉淀矿物质或捕获矿物颗粒而形成的叠层石(Awramik and Sprinkle, 1999)。叠层灰岩在岩心上呈深灰色块状, 未见叠层构造(图 3-a); 而叠层云岩由暗紫红色和浅灰色纹层构成缓丘状, 中间夹浅灰色角砾状凝块(图 3-b)。显微镜下, 叠层灰岩呈波状或丘状, 由丝状的钙质微生物粘结和捕获灰泥形成的微亮层与微暗层交互组成(图 4-a), 而叠层云岩因白云石化作用导致原岩组构破坏。叠层灰岩反映一种相对低能的沉积环境, 通常出现在变浅序列顶部。该类岩石中缺少生物碎屑颗粒且具有纹层与凝块状结构, 其中缺乏生物碎屑表明水体能量低且相对局限。可与Flü gel(2010)提出的标准微相SMF20— 纹层状叠层石粘结灰岩相类比, 常见于潮间带。研究区内的叠层灰岩、叠层云岩分别见于ST9井和葛底坝露头剖面。
 | 图 3 川西北栖霞组微生物岩宏观特征 a— 叠层灰(云)岩(DOL表示云化斑块, LM表示灰岩基岩), ST9井, 7709.68m, 栖二段; b— 叠层云岩, 大木垭剖面, 栖二段; c— 凝块灰岩, ST9井, 7701.09m, 栖二段; d— 凝块云岩, ST9井, 7750.55m, 栖一段; e— 凝块云岩, ST9井, 7748.00m, 栖一段。部分照片位置见图2Fig.3 Macrophotos showing microbialite characteristics of the Qixia Formation in northwestern Sichuan Basin |
 | 图 4 川西北栖霞组微生物岩微观特征 a— 图 3-a对应的显微镜下特征, 左侧为中晶云岩斑块, 右侧见丝状钙质微生物构成的叠层灰岩, ST9井, 7709.68m, 栖二段, 单偏光; b— 图 3-c对应的显微镜下特征, 凝块灰岩, 发育不规则窗格组构, ST9井, 7701.09m, 栖二段, 单偏光; c— 凝块灰岩, 见囊状菌体, 其壳为泥晶, 内充填他形亮晶方解石, 长江沟剖面, 单偏光; d— 图 3-d对应的显微镜下特征, 凝块云岩, ST9井, 7750.55m, 单偏光; e— 凝块云岩, 可见不规则状窗格孔, ST9井, 7748.00m, 栖一段, 单偏光; f— 图 4-e对应的显微镜下特征, 薄片下加垫70g 的A4纸用以恢复原岩组构, 凝块云岩, 可见窗格组构, ST9井, 7748.00m, 栖一段, 单偏光; g— 微生物粘结球粒云岩, 发育粒间溶孔, 长江沟剖面, 蓝色铸体薄片, 单偏光; h— 图 4-g位置薄片下加垫70g 的A4纸用以恢复原岩组构, 长江沟剖面, 蓝色铸体薄片, 单偏光; i— 微生物粘结似球粒灰岩, 他形亮晶方解石胶结, 长江沟剖面, 单偏光。部分照片位置见图2Fig.4 Microphotos showing microbialite characteristics of the Qixia Formation in northwestern Sichuan Basin |
凝块灰(云)岩在研究区露头和钻井剖面中均有发现, 宏观上为浅灰色— 褐灰色块状(图 3-c, 3-d, 3-e)。这类岩石是由微生物粘结、捕获灰泥和少量颗粒而成的具有典型凝块组构特征的凝块石, 显微镜下明显可见微生物粘结团块和格架, 有时可见薄壁微管状的微生物实体, 以发育微生物粘结格架形成的或者微生物死亡腐烂后留下的、大小不一的窗格孔洞和鸟眼孔为特征(图 4-b, 4-c, 4-d)。窗格孔和鸟眼孔往往被亮晶胶结物半充填— 全充填, 分布具有较强的非均质性, 从而区别于颗粒支撑的、大小和分布较均匀的粒间孔。白云石化后, 原岩组构或许遭到破坏而表现为晶粒云岩, 但具有残余原岩组构幻影的晶粒云岩, 仍可通过原岩组构恢复, 识别出粘结结构、凝块组构和窗格孔等微生物岩的典型标志(图 4-e, 4-f)。
凝块灰(云)岩中微生物粘结作用繁盛, 窗格构造明显, 是微生物丘的重要组分, 常出现在丘核部位。大致可与Flü gel(2010)提出的标准微相SMF21— 窗格泥粒灰岩和粘结灰岩相类比, 指示潮间带至潮下带的相对低能至扰动的低能水体。
微生物粘结颗粒灰(云)岩也称为微生物粘结岩或隐藻粘结岩, 主要由微生物捕获、粘结诸如球粒、似球粒、包粒、生物碎屑、砂屑等碎屑颗粒以及灰泥而成, 并以共生发育碎屑结构和微生物粘结结构为典型特征(图 4-g, 4-i)。粒间孔常被亮晶胶结物充填, 分布具有一定的非均质变化。依据颗粒成分, 可以进一步细分为微生物粘结球粒灰(云)岩(图 4-g, 4-h)、微生物粘结似球粒灰(云)岩(图 4-i)、微生物粘结砂屑灰(云)岩、微生物粘结生物碎屑灰(云)岩等类型。
微生物粘结颗粒灰(云)岩是微生物丘中重要的岩石类型之一, 垂向上往往发育于微生物丘旋回的下部或上部, 构成微生物丘的丘基或丘核。大致可与Flü gel(2010)提出的标准微相SMF16— 纹层似球粒粘结灰岩相类比, 指示具有中等循环的局限浅海环境。
微生物丘是主要由原地钙化微生物或非钙化微生物作用形成的碳酸盐建隆(Riding., 2002; Webby, 2002; 张廷山等, 2008), 在生长发育过程中由微生物菌落粘结、捕获大量沉积物, 建造速率高于相邻的其他微环境, 沉积厚度也大于同期沉积物, 从而呈正向微地貌凸起。研究区栖霞组微生物丘的发育规模大小不一, 在露头剖面上单个微生物丘的实测高度一般为几十厘米至几米不等, 宽度通常变化于几米至几十米之间, 具有底平顶凸的典型丘形外貌, 明显可见随后的海侵沉积物超覆沉积于丘体之上, 并且以发育向上变浅的沉积序列为特征(图 5)。笔者以不同地区的2个微生物丘为代表阐述其宏观特征。
 | 图 5 川西北典型露头剖面上的栖霞组微生物丘宏观特征 a— 大木垭剖面微生物丘宏观特征, 丘体高约1m, 可见明显的丘形外貌, 丘顶部见原地角砾, 红色圆点及编号为薄片取样位置; b— 长江沟剖面微生物丘宏观特征, 丘体高约6.15m, 宽约23m, 可见丘形外貌, 上覆地层超覆沉积, 位置见图2; c— 何家梁剖面微生物丘宏观特征, 丘体高79cm, 可见丘形外貌, 上覆地层超覆沉积在丘体上, 红色圆点及编号为薄片取样位置Fig.5 Macroscopic characteristics of microbial mounds of the Qixia Formation on typical section in northwestern Sichuan Basin |
在广元大木垭露头剖面中栖霞组近顶部发育1个微生物丘, 高约1m, 宽度大于5m, 呈典型的丘形地貌特征(图 5-a)。该丘体整体上呈浅灰色— 灰色块状, 全部由微生物粘结生物碎屑灰岩构成, 自丘底至丘顶, 微生物菌落与生物碎屑、灰泥形成的粘结团块逐渐呈小→ 大→ 小变化, 亮晶充填的孔隙形状及大小变化大(图 6-a, 6-b, 6-c)。受高频海平面相对下降的影响, 这个微生物丘顶部遭受准同生期暴露岩溶作用的改造, 形成原地角砾岩(图 5-a)。丘体上覆中厚层生物碎屑泥晶灰岩, 为下一期海侵沉积物。由此可见, 这个微生物丘整体上以发育微生物粘结生物碎屑灰岩和向上变浅的沉积序列为典型特征。
 | 图 6 川西北大木垭和何家梁露头剖面栖霞组微生物岩微观特征 a, b, c— 微生物粘结生物碎屑灰岩, 生屑普遍泥晶化, 局部亮晶方解石胶结, 大木垭剖面, 单偏光, 取样位置见图 5-a; d— 泥粉晶云岩, 中部可见已充填的微裂缝, 何家梁剖面, 单偏光; e— 凝块云岩, 见晶洞构造, 何家梁剖面, 单偏光; f— 凝块云岩, 见凝块组构, 何家梁剖面, 单偏光。d, e, f 取样位置见图2, 图 5-c, 图7-Ⅰ aFig.6 Microphotos showing microbialite characteristics of the Qixia Formation at Damuya and Hejialiang sections in northwestern Sichuan Basin |
在青川县何家梁剖面栖霞组下部发现了另一个微生物丘, 丘体高79cm, 总体具丘形地貌特征(图 5-c), 主体由凝块云岩构成。丘体下伏地层为泥粉晶云岩(图 6-d), 推测沉积于平均浪基面以下的局限潟湖环境; 向上过渡为凝块云岩, 具有先期未被充满的窗格构造和后期遭扩溶形成的孔洞(图 6-e); 丘体上部由凝块云岩组成, 其中可见大量白云石化的凝块组构(图 6-f)。受高频海平面相对下降的影响, 丘体顶部露出海面, 形成暴露面, 出现沉积间断并遭受大气淡水淋滤改造, 发育近地表假角砾, 暴露面上可见丘间细粒沉积物超覆沉积在微生物丘体之上(图 5-c), 结束了这个微生物丘的发育。由此可见, 何家梁微生物丘整体上以发育凝块云岩和向上变浅的沉积序列为典型特征。
与经典生物礁组合类似, 栖霞组微生物丘也发育有较为明显的丘组合。基于对露头剖面和钻井岩心剖面中不同岩石类型与沉积特征垂向叠置关系的分析, 在栖霞组发育完整的微生物丘组合中通常可以识别出丘基、丘核和丘盖3种微相。
如图 7-I所示, 何家梁剖面栖霞组微生物丘组合由多种白云岩组成, 从下到上发育丘基、丘核、丘盖3种微相。
 | 图 7 川西北栖霞组微生物丘组合的典型微相序列 Ⅰ — 何家梁剖面栖霞组微生物丘的微相序列; Ⅱ — ST9井栖霞组微生物丘滩组合的微相序列; 微观照片, 取样位置见该图中岩心照片, ST9井, 单偏光: a— 晶粒云岩, 粒径0.15~0.5 mm; b— 白云质凝块灰岩, 见窗格孔构造; c— 凝块白云质灰岩; d— 凝块白云质灰岩, 含似球粒; e— 凝块灰岩, 左下部可见1个较大的孔洞被渗流粉砂充填; f— 微生物粘结似球粒灰岩, 含包粒和生物碎屑。DOL为白云岩, LIM为灰岩Fig.7 Typical microfacies sequence of microbial mounds of the Qixia Formation in northwestern Sichuan Basin |
丘基微相位于丘组合的底部, 是微生物丘发育的基础, 主要由低能丘间海环境沉积的泥粉晶云岩组成(图 6-d)。丘核微相处于丘组合的中部, 是微生物丘组合的核心和主体, 为造丘微生物发育繁盛阶段的产物, 以发育凝块云岩为特征(图 6-e, 6-f)。由下而上, 凝块组构增多, 颗粒减少, 反映沉积水体向上逐渐变浅。伴随微生物丘的不断向上建造, 高频海平面相对下降, 最终导致微生物丘出露海面停止生长, 并遭受大气淡水淋滤改造, 在丘顶部形成原地角砾(图 7-Ia)。丘盖微相位于丘组合的顶部, 覆盖在微生物丘上, 主要由低能丘间海环境沉积的泥粉晶云岩组成, 反映高频海平面相对上升。
如图 7-Ⅱ 所示, ST9井7709.6~7711.5 m栖霞组岩心以发育微生物丘滩组合为特征, 微生物丘与颗粒滩沉积呈薄互层, 即一层颗粒滩沉积可以成为某个微生物丘的丘基, 同时又可作为下伏一个微生物丘的丘盖, 即两者互为“ 基” 与“ 盖” 。ST9井丘滩组合从下到上发育生屑滩(丘基)、微生物丘(丘核)、生屑滩(丘盖)3种微相。
生屑滩(丘基)微相位于丘滩组合的底部, 由灰色晶粒云岩组成, 推测为生屑滩沉积的白云石化产物。
微生物丘(丘核)微相位于丘组合的中部, 厚约130cm, 全部由微生物灰岩组成。底部1层厚约10cm, 岩性主要为相对高能的亮— 泥晶微生物粘结似球粒灰岩和凝块灰岩(图 7-Ⅱ e, 7-Ⅱ f), 指示高频海侵初期的相对高能环境, 并可见渗流粉砂(图 7-Ⅱ e)。下部为微生物粘结灰泥和少量的似球粒组成的白云质凝块灰岩(图 7-Ⅱ d); 向上包粒含量增加, 指示能量进一步增强, 发育窗格孔(图 7-Ⅱ b, 7-Ⅱ c)。上部为微生物粘结灰泥构成的白云质凝块灰岩, 云化部分总体呈水平状分布(图 7-Ⅱ ③)。
生屑滩(丘盖)微相位于丘组合的上部, 厚约48cm, 推测是微生物丘的建造导致海水变浅, 使其处于浪基面之上的相对高能地带而发育生屑滩沉积。因后期的白云石化和重结晶作用破坏原岩组构, 现为中、细晶云岩, 晶粒间充填渗流白云石粉屑(图 7-Ⅱ a)。宏观上, 下部为浅灰色块状晶粒云岩, 见溶蚀孔洞和叠层构造(图 7-Ⅱ ②); 上部受高频暴露岩溶作用改造, 形成近地表角砾化; 顶部为不平整的岩性突变面(图 7-Ⅱ ①), 其上为下一次高频海侵形成的微生物丘。
这一类微生物丘滩微相组合在栖霞期发育频率高, 指示随着微生物丘向上建造导致海水逐渐变浅而发生暴露岩溶; 而白云岩发育于向上变浅序列的上部, 则指示白云石化与海平面升降密切相关, 与微生物岩的发育位置关系并非密切。该类微生物丘滩微相组合可能受向上变浅和丘滩体叠置迁移导致的环境进一步受限和局部蒸发控制。
如前所述, 本次研究在四川盆地西北部野外露头和钻井剖面的栖霞组中发现了多个微生物丘, 造丘的岩石类型主要为叠层灰(云)岩、凝块灰(云)岩、微生物粘结颗粒灰(云)岩等经典的微生物岩类型。进一步研究发现, 发育于海相碳酸盐岩台地上的这些微生物岩, 均指示栖霞期沉积水体能量相对较低、循环不畅且相对受限。其次, 栖霞组微生物丘绝大多数以发育向上变浅的沉积序列为特征, 反映了高频海平面逐渐相对下降, 沉积水体逐步收缩, 微生物丘发生侧向迁移叠置, 导致沉积环境进一步受限。当环境进一步受限时, 蒸发浓缩作用会使海水的Mg/Ca值升高, 从而导致局部发生准同生期白云石化。图 7-Ⅱ 指示的白云岩发育与向上变浅序列的封隔受限密切相关也证实了这一认识。另一方面, 微生物与后生动物存在互为消长关系, 凝块灰(云)岩或叠层灰(云)中少见宏观生物化石(图 4), 即使有后生动物化石产出, 其属种不仅单一, 而且个体也较小, 表明当时的沉积环境可能不正常。在野外露头的某些微生物丘剖面上, 例如广元西北乡车家坝剖面, 下部见有个体较小的有孔虫、海百合和介形虫等化石, 向上后生动物化石减少, 仅见少量介形虫碎片, 且出现凝絮状微生物粘结生物碎屑和灰泥的现象。这种凝絮状微生物粘结组构一般指示海水盐度不正常、电解质失衡。
实际上, 已有越来越多的研究成果揭示了包括川西北在内的四川盆地早二叠世栖霞期的古海洋环境总体可能受限。如马永生等(2009)指出栖霞期上扬子台地西部为局限环境, 其余地区为开阔环境; 刘治成等(2015)基于华蓥山地区栖霞组灰泥丘的研究, 认为川中地区该时期可能存在环境受限; 刘正元等(2020)通过对川西北地区上寺猫儿塘二叠系剖面的古生物研究发现, 栖霞初期的部分类化石缺失可能与水体环境较为闭塞有关; 李明隆等(2020)和芦飞凡等(2020)通过对白云岩宏微观特征及地球化学研究, 也得出了川西北地区栖霞期古环境受限的结论。综上所述, 可以认为川西北地区栖霞期沉积环境总体受限, 推测为半局限— 局限台地环境, 海域水深不大, 能量普遍不高, 从而出现了优势的凝絮状的微生物粘结球粒、似球粒或生物碎屑等微生物灰岩构建的所谓“ 中低能滩” , 实际上应该属于微生物丘滩组合范畴。
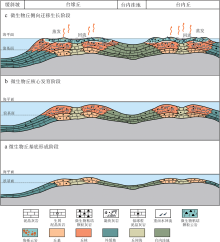
造成川西北地区栖霞期沉积环境受限的另一个重要因素与古构造和古地理背景相关。四川盆地海西期以来经历了多期构造运动(罗志立, 2009; 王鼐等, 2016), 石炭纪末发生的云南运动造成地壳抬升, 地层广泛遭受剥蚀, 川西一带普遍存在地层削蚀和准平原化。伴随二叠纪广泛海侵, 原有的古地理格局一定程度上转变为局部地貌隆坳起伏的缓坡格局(宋文海, 1987; 刘春平和朱国华, 2006; 刘小洪等, 2017)。早二叠世栖霞期, 海侵使上扬子地区演变为辽阔的浅水碳酸盐岩台地环境, 并且在四川盆地西北缘的广元— 江油一带形成了有镶边的台地边缘, 其西侧推测为水体较深的缓斜坡, 东接平缓起伏的碳酸盐岩台地。但从川西北地区栖霞组频繁的高频暴露特征来看(李明隆等, 2020), 海水总体较浅, 能量不高, 且高频海平面相对升降变化频繁, 导致发育于缓坡背景下的碳酸盐岩台地极易受限, 台地边缘也未发现亮晶颗粒岩和/或生物礁等组成的高能相带, 而是以发育微生物丘滩组合为代表的中低能弱镶边体系为特征(图 8)。
 | 图 8 川西北栖霞组微生物丘滩沉积与白云石化模式Fig.8 Depositional model of microbial mound-shoal complexes and correspondent dolomitization of the Qixia Formation in northwestern Sichuan Basin |
如图 8所示, 在高频初始海侵期, 在台地边缘和台内水下微地貌高地上开始沉积微生物粘结似球粒— 包粒灰岩, 构成微生物丘的生长基础; 在台内洼地等相对地貌低地, 则以发育泥晶灰岩和似球粒或包粒泥晶灰岩为特征(图 8-a)。随着基底稳定沉降或持续海侵的进行, 微地貌高地上连续沉积凝块岩和微生物粘结的生物碎屑灰岩或似球粒灰岩, 构成微生物丘的核心; 由于微生物丘的建造速率较快, 强化了丘体与丘间的地貌差异, 导致微生物丘的生长空间受限(图 8-b)。当丘体建造至平均海平面附近时, 因可容空间变小, 微生物丘发生侧向迁移生长, 使局部环境进一步受限, 在较强的蒸发作用影响下, 海水逐渐浓缩, 重卤水下沉并向低部位回流, 交代早期沉积物, 发生与丘滩发育密切相关的准同生期白云石化作用(图 8-c); 丘体顶部或因出露海平面而遭受岩溶改造, 形成近地表角砾岩。据上所述, 研究区栖霞期沉积环境的进一步受限也可能与微生物丘的侧向迁移叠置有关, 白云岩的形成或许与沉积环境进一步受限更为密切, 而非微生物诱导白云石化。推测栖霞组与微生物丘相关的白云岩, 纵向分布大概与多级海平面升降变化有关, 横向上可能与丘滩体发育和叠置迁移位置相关, 总体上可能具有分布层位不稳定的特征。但从总体起伏不大的古地理背景推断, 最容易导致进一步受限的古地理位置应为台缘坡折带和台内缓坡折带。
微生物岩具有良好的储集性能, 其中微生物结构和白云石化是微生物白云岩储集层形成的有利因素(李凌等, 2013; 赵文智等, 2014; 李朋威等, 2015)。但就川西北地区而言, 现今微生物岩结构保存完整的灰岩和云岩中, 与微生物组构相关的原生孔隙往往较小, 且难以见到格架孔洞, 多因胶结而丧失(图 4)。能见到明显储集空间的多是具有微生物粘结组构的球粒或生物碎屑白云岩, 以发育溶扩粒间孔为特征(图 4-g), 这表明微生物岩的总体储集性能较差, 推测可能与这一时期微生物岩形成时的水体能量较低有关。
台内和台缘的勘探实践表明, 与微生物丘相关的白云岩储集性能良好, 是主要的油气勘探对象。李明隆等(2020)认为, 栖霞组白云岩优质储集层的发育主要与白云岩高频暴露岩溶引起的不同程度改造有关, 而白云石化为准同生期回流渗透白云石化叠加同期海水循环热液成因。前已述及, 造成局部环境进一步受限和白云石化的原因为微生物丘滩的叠置迁移, 且储集层主要发育于微生物建造相关的丘滩组合中。据此认为, 微生物丘滩复合体是栖霞组白云岩储集层发育的物质基础, 台缘坡折带、台内缓坡折带和高地是栖霞组白云岩储集层发育的有利区带。
1)在四川盆地西北部(川西北)下二叠统栖霞组中新发现的微生物丘, 主要由凝块灰(云)岩、叠层灰(云)岩和微生物粘结颗粒灰(云)岩等组成。
2)川西北栖霞组微生物丘大小不一, 实测高度一般为几十厘米至几米, 宽度通常变化于几米至几十米之间, 具有底平顶凸的典型丘形外貌, 以发育向上变浅的沉积序列为特征, 一般由丘基、丘核、丘盖3个微相组成, 也可与颗粒滩共同构成微生物丘滩复合体。
3)川西北地区栖霞期沉积环境总体受限, 推测为半局限— 局限台地环境, 水深总体较浅, 能量普遍不高; 海平面频繁的相对升降变化和微生物丘的侧向迁移叠置, 导致发育于缓坡背景下的碳酸盐岩台地极易受限, 引起早期沉积物发生与丘滩发育密切相关的准同生期白云石化作用。
4)微生物丘滩复合体是栖霞组白云岩储集层发育的物质基础, 台缘坡折带、台内缓坡折带和高地是栖霞组白云岩储集层发育的有利区带。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|