





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
鄂尔多斯盆地奥陶系马家沟组马五5亚段微生物岩盆地尺度分布特征及主控因素
[杨梦颖1, 2  , 谭秀成
, 谭秀成1, 2 , 杨鸣一3 , 许玉青4 , 晏巍5 , 苏文杰6 , 钟寿康1, 2 , 熊鹰1, 2 , 郭木石7 ]
, 谭秀成, 杨鸣一|
|

通讯作者简介 谭秀成,男,1970年生,博士,教授,主要从事沉积学与储层地质学研究工作。E-mail:tanxiucheng70@163.com。
第一作者简介 杨梦颖,女,1997年生,西南石油大学研究生,主要从事沉积学研究工作。E-mail:ymying22@163.com。
基于岩心、薄片和测录井资料分析发现,鄂尔多斯盆地奥陶系马家沟组马五 5亚段下部发育 1套盆地尺度稳定分布的微生物碳酸盐岩组合,其特征为: ( 1)岩石类型主要由泥质灰(云)岩、泥晶灰(云)岩、凝块灰(云)岩、叠层灰(云)岩、粘结砂屑灰(云)岩及岩溶角砾灰(云)岩等构成,凝块石主要呈层状、瘤状和格架状,叠层石可识别出层纹状、波状和柱状 3种类型;( 2)垂向上可识别出 4种与微生物岩密切相关的、向上变深复变浅的典型沉积序列;( 3)高频旋回小层划分与对比分析发现,微生物岩在横向上稳定连片分布,具有盆地尺度的可对比性和等时性;( 4)基于岩心刻度测井方法建立测井相模板,依据该模板对全盆地 404口井进行逐一识别和解释,发现微生物碳酸盐岩的累计厚度一般在 3~7m之间,整体呈向东减薄的变化趋势,连续分布面积超过 2.5× 105km2,揭示了较短时间内盆地尺度微生物岩规模发育的独特沉积现象。沉积序列分析表明,马五 5亚段沉积初期存在单旋回缓慢变深复变浅、多旋回组合振荡式持续缓慢海侵的海平面变化趋势,且相对平静的古构造环境与早期蒸发岩填平补齐作用的耦合提供了相对平坦的古地貌条件,因此在马五 5亚段沉积时期总体持续缓慢海侵背景下,环境同质化的底形条件是规模性浅水微生物碳酸盐岩发育的根本原因。系统报道马五 5亚段盆地尺度微生物岩的发育与分布特征,既丰富了鄂尔多斯盆地奥陶系马家沟组微生物岩的研究,也有助于丰富完善对碳酸盐岩台地微生物岩规模性繁育机制的认识。
, Tan Xiu-Cheng, Yang Ming-YiAbout the corresponding author Tan Xiu-Cheng,born in 1970,is a professorat in Southwest Petroleum University. He is engaged in researches of sedimentology and reservoir geology. E-mail:tanxiucheng70@163.com.
About the first author Yang Meng-Ying,born in 1997,is an graduate studentat in Southwest Petroleum University. Her studies focus on sedimentology. E-mail:ymying22@163.com.
Based on the analysis of core,thin section and logging data,basin-scale microbial carbonate with stable distribution developed in the lower part of the Ma55 sub-member of the Ordovician Majiagou Formation in Ordos Basin. The rock types include argillaceous limestone,limestone,stromatolite,thrombolite,bound sand-clastic limestone,karst breccias. It contains various shaped stromatolite and thrombolite,such as laminar,wavy and columnar stromatolite,laminar,nodular and arborescent thrombolite. Vertically,four typical sedimentary sequences closely related to microbialites are identified,indicating relative sea-level fluctuations. The division and comparative analysis of the small layers with high-frequency cycles suggest that the microbialites are stably distributed laterally,and have basin-scale distribution. Based on the logging template established by core-log calibration,the lithology of 404 wells in the whole basin were interpreted.The result shows that cumulative thickness of microbialites range from 3 to 7m,and the thickness shows a decreasing trend from west to east.Furthermore,the continuous distribution area of microbialites can exceed 2.5×105km2in area,demonstrating that the basin-scale microbialites developed in a relatively shorter time.Analysis of sedimentary sequence in the early depositional stage of Ma55 sub-member indicates that the relative sea level first slowly rises and then falls in a single cycle,while it shows an oscillating and slowly rising trend in a multi-cycle combination.Further research suggests that the relatively stable paleo-tectonic environment coupled with the filling of early evaporite provides relatively flat paleo-geomorphic conditions.Under the background of continuous slow transgression in the depositional stage of Ma55 sub-member,the bottom shape condition of environmental homogeneity is the fundamental reason for the development of large-scale microbial carbonate in shallow water.This paper reports the development and distribution of basin-scale microbialites in the depositional stage of Ma55 sub-member,which not only enriches the research of microbialites in the Majiagou Formation,but also helps improve the understanding of development mechanism of large-scale microbialites in carbonate platforms.
开放科学(资源服务)标识码(OSID) 
作为微生物— 环境相互作用的沉积产物, 微生物碳酸盐岩发育在温暖、清澈以及较浅的水体环境中(Riding, 1991; Browne et al., 2000; Rezende et al., 2013), 结构形态和分布模式易受控于海平面变化、水动力条件、水体盐度(Lindsay et al., 2017)、古构造与古地貌等外部环境因素(Reid et al., 2000; 何起祥, 2003; Woo et al., 2008; 韩作振等, 2009; 杨仁超等, 2013)。较为敏感的发育条件, 导致微生物碳酸盐岩集中发育在台地边缘、台内局部高地以及潮下沉积环境(Kennard and James, 1986; Rezende et al., 2013; 刘静江等, 2016; 徐欣等, 2018), 常呈孤立点丘状或带状展布(几十至上百平方千米), 平面分布规模较为有限, 例如四川盆地震旦系灯影组发育裂陷边缘丘滩, 呈U形分布, 宽5~40km、长约500km(Shapiro, 2000; Webby, 2002; 李凌等, 2013; 周进高等, 2017)。
受沉积底形、水动能条件以及微生物对适宜环境的优选等因素影响(曹红霞等, 2021; 胡安平等, 2021), 鄂尔多斯盆地马家沟组报道的微生物岩以零星分布的微生物点丘或丘滩复合体形式存在, 且多集中于海退背景的马五6、马五1+2等亚段(熊鹰等, 2016; 席胜利等, 2017; 乔亚斌等, 2020)。近期, 基于鄂尔多斯盆地系统取心井的精细岩心观察发现, 马五5亚段底部可能稳定发育2期微生物岩, 这一发现在马家沟组诸多微生物岩研究中尚属首次。此外, 微生物岩可作为环境信息的载体(Whalen et al., 2002), 对其结构形态及纵向叠置关系的研究, 有利于揭示马五5亚段沉积初期水动力条件和演变规律, 且平面分布特征对沉积期地貌形态、沉积格局的厘定至关重要。
因此, 文中尝试通过系统的岩石学和测录井资料, 理清研究区马五5亚段微生物岩的沉积特征和空间分布规律, 进而探讨该套规模性微生物岩的发育主控因素。该研究成果可丰富微生物岩研究案例, 同时为马五5亚段沉积初期鄂尔多斯盆地水动力条件与古地貌恢复等沉积学研究提供重要的证据。
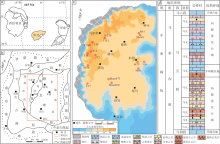
鄂尔多斯盆地位于华北台地西缘, 面积约为3.7× 1km2。中寒武世至中奥陶世, 华北地块南、北两侧海槽洋底扩张, 因均衡作用于鄂尔多斯盆地中部偏西地区形成“ L” 形大型隆起带(图 1-b; 侯方浩等, 2002)。隆起东侧(研究区)以浅水碳酸盐岩台地沉积为主(Wang et al., 2016)。中奥陶世马家沟组沉积时期研究区经历了3次海侵— 海退旋回式沉积, 其中马一段、马三段和马五段为高位体系域沉积, 马二段、马四段和马六段为海侵体系域沉积(史基安等, 2009; 张永生等, 2015)。
 | 图 1 鄂尔多斯盆地中奥陶世区域地质背景 a— 中奥陶世(465Ma)华北板块位置(据Wang et al., 2016; 有修改); b— 中奥陶世鄂尔多斯盆地内部构造单元(侯方浩等, 2002)及研究区位置; c— 鄂尔多斯盆地中东部马家沟组马五5亚段地层等厚图及取心井位置; d— 奥陶系马家沟组五段地层柱状剖面示意图Fig.1 Geological setting of the Middle Ordovician in Ordos Basin |
马五段发育海退沉积, 自下而上分为马五10— 马五1共10个亚段(黄正良等, 2015; 席胜利等, 2017)。中部马五5亚段为次一级短周期海侵沉积, 海水侵入范围最大, 沉积水体较深, 主要发育深灰色泥晶灰岩和泥粉晶云岩(图 1-d), 地层厚度平均为25m, 在盆地中东部分布稳定(图 1-c)。马五5亚段沉积前, 盆地内广泛发育碳酸盐岩与膏盐岩的韵律共生沉积, 尤其在马五6亚段沉积了最大规模的巨厚蒸发岩(图 1-d)(胡彬等, 2014; 樊馥等2015)。
根据大量岩心和薄片的精细观察, 结合前人的分类方案(吴亚生等, 2018), 在马五5亚段下部梳理出叠层石、凝块石以及与微生物相关的颗粒岩共3大类微生物碳酸盐岩。
叠层石是由于周期性沉积作用和捕获细粒沉积物形成的纹层状底栖微生物沉积物, 岩心上为明暗相间的毫米级纹层韵律构成的叠层构造(图 2-b, 2-c), 局部纹层揉皱变形, 可分为层纹状、波状和柱状3种亚类。
 | 图 2 鄂尔多斯盆地奥陶系马家沟组马五5亚段微生物碳酸盐岩宏观特征 a— 层纹状叠层灰岩, T112井, 3314.31m; b— 波状叠层云岩, Mt1井, 2418.10m; c— 柱状叠层云岩, Mt1井, 2419.32m; d— 层状凝块云岩, JT1井, 3550.69m; e— 瘤状凝块灰岩, 暗色部分泥晶方解石含量较高, T112井, 3316.25m; f— 格架状凝块云岩, T18井, 3702.75m; g— 格架状凝块灰岩, T36井, 3338.26m; h— 粘结砂屑云岩, 桃36井, 3338.38m; i— 微生物团块灰岩, T36井, 3334.30m; 红色箭头指示凝块结构Fig.2 Macrofeature of microbial carbonates of the Ma55 sub-member of Ordovician Majiagou Formation in Ordos Basin |
层纹状叠层石的纹层为近水平— 微波状, 单纹层厚度不均一, 边缘具小型凹凸起伏, 横向上纹层延续性较好, 常见生物扰动构造截断纹层(图 2-a)。与层纹状叠层石相比, 波状叠层石的纹层波状起伏更大, 单纹层厚度较均一, 边缘光滑, 横向延续性更好(图 2-b)。柱状叠层石的纹层为半球状, 具有变化的宽度或固定的直径(多为厘米级), 横向延续性较差, 呈间断性丘状起伏并堆积成柱体, 且这些柱体被沉积物分隔开或相互联结, 整体具有向上生长的趋势(图 2-c)。
微观下, 叠层石暗层较薄, 为富有机质层, 由暗色微生物组构和泥晶沉积物组成(图 3-a), 局部可见被捕获的球粒(图 3-b); 亮层稍厚, 为贫有机质层, 泥晶沉积物增多, 粘结球粒分散于泥晶基质中; 两者之间界线模糊, 呈渐变过渡。纹层间发育不规则状窗格孔, 内部充填亮晶方解石/白云石胶结物(图 3-a, 3-b, 3-c)。叠层石多发育在强蒸发环境的潮间带和潮下带(Kennard and James, 1986), 常见板条状石膏与叠层石伴生(图 3-c), 以及发育与早期大气淡水淋溶有关的溶孔和基质溶蚀微孔(图 3-b)。
 | 图 3 鄂尔多斯盆地奥陶系马家沟组马五5亚段微生物碳酸盐岩微观特征 a— 层纹状叠层灰岩, T112井, 3314.31m; b— 波状叠层云岩, Mt1井, 2418.10m, 蓝色铸体薄片, 发育膏摸孔; c— 柱状叠层云岩, Mt1井, 2419.32m, 黄色箭头指示石膏; d— 层状凝块云岩, JT1井, 3550.69m; e— 瘤状凝块灰岩, 顶部泥晶方解石含量较高, T112井, 3316.25m; f— 格架状凝块云岩, 左侧泥晶、粉晶白云石含量较高, T18井, 3702.75m, 蓝色铸体薄片, 发育晶间溶孔; g— 格架状凝块灰岩, 白色箭头指示格架生长方向, T36井, 3338.26m; h— 粘结砂屑云岩, 桃36井, 3338.38m; i— 微生物团块灰岩, T36井, 3334.30m。红色箭头指示微生物组构, 橙色箭头指示亮晶方解石/白云石胶结物Fig.3 Microfabric characteristics of microbial carbonates of the Ma55 sub-member of Ordovician Majiagou Formation in Ordos Basin |
凝块石为底栖微生物群落形成的微生物岩, 岩心上呈浅灰色— 深灰色或者浅灰色— 灰褐色, 以发育凝块结构为典型特征。根据凝块形态和连接方式的差异, 可分为3种亚类: 层状、瘤状和格架状。层状凝块石具明暗相间揉皱条带状结构, 与层纹状叠层石相比, 单纹层厚度稍大且层内厚度不均匀, 边缘凹凸起伏明显; 横向上, 单条带延展性较弱, 出现纹层分叉或合并现象(图 2-d)。瘤状凝块石的凝块呈似扰动的瘤状, 边缘稍圆滑, 颜色偏浅, 大小不等, 大者可达5cm, 凝块间接触紧密, 以堆积集合体形式产出(图 2-e)。格架状凝块石中凝块相互连接, 搭建成横向和垂向延伸交汇的网络状, 边缘不规则, 白云岩化后颜色偏暗(图 2-f, 2-g)。
微观下, 凝块石的暗色凝块由近圆形或椭圆状的球粒相互连接构成, 球粒内部为均质的暗色泥晶方解石, 边缘模糊。层状凝块结构为多个凝块顺层状搭连汇聚形成的、侧向延伸的暗色条带状集合体(图 3-d); 瘤状和格架状凝块结构为多个凝块不定向缠绕聚集而成, 单个凝块内前者球粒较分散, 泥晶方解石增多(图 3-e), 后者生物格架具向上生长特征(图 3-g)。白云石化凝块的内部无球粒结构, 发育暗色不规则泥晶白云石斑块, 存在局部重结晶现象(图 3-e, 3-f)。此外, 格架间常为粘结微生物生长的空腔或残余孔隙, 后期被填隙物或胶结物(方解石或白云石)充填/半充填(图 3-f)。凝块结构与凝块间界面较模糊, 以凝块沉积过渡为泥晶方解石沉积为主, 见少量微生物组构和球粒分布。凝块石在研究区发育频率高, 主要沉积于水体能量相对较低的潮下带(Rezende et al., 2013)。
1)粘结砂屑灰(云)岩。岩心上呈灰褐色, 自然断面粗糙, 颗粒结构明显, 顺层分布(图 2-h)。微观下, 砂屑颗粒色暗, 被微生物组构包裹粘结, 呈悬浮状孤立分布, 不具定向排列(图 3-h)。砂屑颗粒含量大于50%, 呈球状或扁条状, 分选差, 磨圆中等, 粒径0.2~2mm, 内部为均质的泥晶沉积物, 边缘不光滑, 砂屑间填充亮晶方解石胶结物。粘结砂屑岩发育于水动力相对较强的沉积环境, 仅在局部井区可见。
2)微生物团块灰岩。岩心上呈灰褐色, 颗粒结构明显(图 2-i)。微观下, 暗色微生物团块含量大于50%, 多具扁条状和不规则边缘, 分选差, 粒径大于2mm, 内部为暗色凝块, 可见球粒和微生物粘结特征(图 3-i)。与凝块石相比, 微生物团块灰岩多为孤立分布, 团块直径较大。团块间沉积褐色泥晶方解石和暗色不溶残余物, 见切割团块组构现象, 指示水体能量较强。
通过岩心及薄片观察发现, 鄂尔多斯盆地马五5亚段下部发育与微生物岩相关的米级沉积旋回, 笔者系统梳理并总结出了4种典型的沉积序列。
1)序列Ⅰ : 凝块灰(云)岩— 泥晶灰岩— 凝块灰(云)岩— 微生物团块灰岩。该序列以JT1井3551.51~3547.35m井段为代表(图 4-a), 典型特征为发育层状凝块灰岩。序列底部沉积纹层状泥云岩, 与下伏岩溶角砾岩呈不整合接触。其上发育凝块石(图 2-d), 且自下而上以层状、格架状、瘤状的不同中型构造类型逐渐过渡, 其中瘤状凝块灰岩呈厚层产出, 向上瘤状团块逐渐变小, 随后转为沉积纹层状泥晶灰岩, 表明水体能量逐渐减小。再向上瘤状灰岩过渡为微生物团块灰岩, 顶部无暴露, 表明水体能量逐渐增大。上覆下一旋回底部的层状凝块灰岩。该序列常发育于研究区西部, 厚3~4m。
 | 图 4 鄂尔多斯盆地奥陶系马家沟组马五5亚段微生物碳酸盐岩典型垂向沉积序列Fig.4 Typical vertical sedimentary sequence of microbial carbonates of the Ma55 sub-member of Ordovician Majiagou Formation in Ordos Basin |
2)序列Ⅱ : 粘结砂屑灰(云)岩— 凝块灰(云)岩— 叠层灰(云)岩— 泥晶灰岩— 凝块灰(云)岩— 微生物团块灰岩。该序列以T36井3338.60~3333.98m井段为代表(图 4-b), 位于盆地中部, 主要由凝块灰岩和微生物团块灰岩构成。序列开始于暴露面之上, 下伏岩溶角砾岩, 呈不整合接触。底部为海侵初期的泥灰质沉积, 之上依次发育泥晶灰岩和薄层粘结砂屑云岩(图 2-h)。向上微生物岩呈直立生长, 发育格架状凝块灰岩(图 2-g), 凝块格架规模减小转变为瘤状凝块, 随后瘤状凝块减小过渡为叠层灰岩, 直至微生物岩停止生长, 沉积纹层状泥晶灰岩。上述这种微生物岩中型构造类型的演变过程表明水体能量先增大再持续减小。顶部沉积厚层微生物团块灰岩(图 2-i), 无暴露, 表明水体能量持续增强。上覆下一旋回底部的格架状凝块灰岩。该序列结构发育频率较低, 厚3~4.5 m。
3)序列Ⅲ : 凝块灰岩— 泥晶灰岩— 生物扰动泥粉晶云岩。该序列以微生物岩仅发育凝块石为典型特征(图 4-d)。格架状凝块灰岩发育于低能的泥晶灰岩基底上, 伴随凝块格架的减少和离散并过渡为瘤状凝块灰岩, 向上沉积均质泥晶灰岩。顶部以泥粉晶云岩中发育生物扰动构造结束, 未暴露。该序列结构在研究区内普遍发育, 厚1.5~2m。
4)序列Ⅳ : 叠层灰(云)岩— 角砾灰(云)岩。该序列以M60井2424.40~2422.30m井段为代表(图 4-d), 典型特征为微生物岩仅发育叠层云岩。序列底部表现为柱状叠层石沉积于岩溶角砾之上, 呈不整合接触, 向上叠层石纹层表现为柱状— 波状— 层纹状逐渐演变的特征, 顶部具暴露溶蚀。研究区内亦可见序列结构中缺少波状— 层纹状叠层石沉积, 以Mt1井2421.85~2418.90m井段为代表(图 4-c), 旋回中柱状叠层石单柱体垂向规模由大变小(图 2-c), 顶部以岩溶角砾岩结束。该类型序列常发育于研究区东部, 厚1.5~2m。
总体上, 研究区以凝块石沉积序列占主导、水体能量具向上持续减弱再增强的特征, 上升半旋回多表现为层状— 格架状— 瘤状凝块石的微生物岩组合模式, 而瘤状凝块灰岩— 微生物团块灰岩组合模式普遍发育于下降半旋回。此外, 以叠层石为唯一微生物岩类型的序列组合, 常在顶部见岩溶角砾岩。
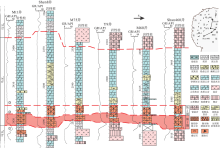
由于沉积水体咸化和海侵— 海退等旋回特征可作为多旋回沉积地层对比划分的可靠依据, 并能达到各层沉积体接近于等时对比(谭秀成等, 2008), 故文中以五级旋回顶面作为对比基准面, 建立了近东西向和南北向的取心井连井剖面(图 5, 图 6), 以揭示微生物岩在横向上的分布特征。
 | 图 5 鄂尔多斯盆地奥陶系马家沟组马五5亚段微生物碳酸盐岩取心井南北向连井对比Fig.5 North-south cross section of six cored wells showing distribution of different microbial carbonates of the Ma55 sub-member of Ordovician Majiagou Formation in Ordos Basin |
 | 图 6 鄂尔多斯盆地奥陶系马家沟组马五5亚段微生物碳酸盐岩取心井东西向连井对比(图例同图 5)Fig.6 East-west cross section of six cored wells showing distribution of different microbial carbonates of the Ma55 sub-member of Ordovician Majiagou Formation in Ordos Basin(the legend is the same as Fig.5) |
依据旋回地层分析发现: (1)测井曲线在横向上相似, 具可对比性, 揭示出马五5亚段下部稳定发育2期等时的微生物岩沉积体, 其由2个五级海侵— 海退旋回构成, 分别位于GR高值①、②(第1期)和②、③(第2期)内(图5)。(2)研究区旋回类型为上升半旋回厚度大于下降半旋回的不完全对称型, 上升半旋回由泥灰(云)岩、泥晶灰(云)岩、凝块灰(云)岩和叠层灰(云)岩组成, 下降半旋回主要由瘤状凝块灰岩和微生物团块灰岩组成。(3)横向上, 微生物岩累计厚3~6m, 呈向东减薄的趋势。第1期微生物岩在研究区东部主要发育柱状叠层石, 中西部主要沉积格架状/瘤状凝块石, 累计厚度分别为0.5~3m和1~3m, 且自西向东由1个五级旋回沉积演变为2个六级旋回沉积。第2期微生物岩厚度小于第1期, 微生物岩普遍发育凝块石, 累计厚度0.5~3m, 呈西厚东薄的变化趋势。(4)近南北向连井剖面中, 第1期微生物岩在研究区北部发育序列Ⅳ , 南部发育序列Ⅱ (图 5), 由北向南表现为叠层石向凝块石演变的特征; 近东西向连井剖面中, 第1期微生物岩在研究区西部发育序列Ⅰ 、Ⅱ , 东部发育序列Ⅳ , 由西向东呈凝块石向叠层石演变的特征(图 6)。横向上第2期微生物岩则主要发育序列Ⅲ , 稳定沉积凝块石(图 5, 图 6)。整体来看, 纵向上呈由叠层石和凝块石沉积向瘤状凝块石沉积占主导的转变, 横向上则表现为西部以凝块石沉积序列和连续沉积为主, 而东部以叠层沉积序列和间断沉积为特征。
由于马五5亚段底部微生物岩发育段的测井曲线具有一定相似性, 结合取心井岩电匹配关系, 总结了4类岩电响应模型。类型一以T18井为代表, 底部GR高值尖为“ 中高两低型” , 向上曲线平缓(图 7-a), 发育薄— 中层叠层石和凝块石。类型二以M60井为代表, 底部GR高值尖为“ 一高一低型” , 第1期沉积叠层石, 第2期沉积凝块石(图 7-b)。类型三以T36井为代表, 底部GR高值尖为“ 两高型” (图 7-c), 向上低幅度波动, 发育粘结砂屑岩。类型四以Y9井为代表, 底部GR高值尖为“ 单高型” , 柱状叠层石发育于蒸发岩之上(图 7-d)。
 | 图 7 鄂尔多斯盆地奥陶系马家沟组马五5亚段微生物碳酸盐岩伽马测井相模板(所有曲线上的紫色代表第1期微生物岩, 蓝色指示第2期微生物岩)Fig.7 Gamma logging template of microbial carbonates of the Ma55 sub-member of Ordovician Majiagou Formation in Ordos Basin(purple represents phase I microbialites and blue indicates phase II microbialites) |
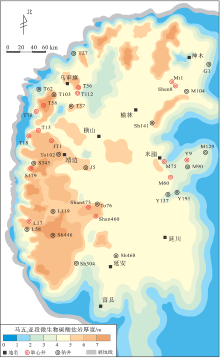
应用建立的测井相模板对研究区404口钻井的微生物碳酸盐岩发育段厚度进行识别, 并绘制了马五5亚段底部微生物碳酸盐岩厚度平面分布图(图 8)。微生物岩在研究区西部厚度最大, 呈向东减薄的变化趋势: (1)微生物碳酸盐岩累计厚度大于7m的区域集中在T58井— S379井— L17井区一带, 而研究区北部仅在神木西侧的2处区域出现, 呈孤立状分布; (2)微生物碳酸盐岩累计厚度在4~7m范围的区域集中分布于乌审旗— 横山— 靖边以西地区, 呈南北向条带状; (3)微生物碳酸盐岩累计厚度在3~4m范围的区域展布范围最广, 占研究区总面积的80%以上。
 | 图 8 鄂尔多斯盆地中东部奥陶系马家沟组马五5亚段微生物碳酸盐岩厚度平面分布Fig.8 Thickness distribution of microbial carbonates of the Ma55 sub-member of Ordovician Majiagou Formation in central and eastern Ordos Basin |
受奥陶纪末期加里东运动影响, 研究区以西马五5亚段地层剥蚀严重, 但中东部地区马五5亚段底部微生物碳酸盐岩序列保存良好, 横向上可对比、追踪性强, 估算平面展布面积可达2.5× 105km2, 约占现今盆地面积的67%, 是一套盆地尺度稳定分布的等时微生物岩体。
海侵— 海退旋回通过改变水体深浅、动能及营养含量等环境因素, 进而影响微生物岩发育的适宜环境、宏观形态以及空间分布(Tucker, 1977; Whalen et al., 2002; Adams et al., 2005)。叠层石可发育于不同水深条件下, 主要在潮间及潮下带(Kennard and James, 1986), 形态易受水体扰动程度控制(Dupraz et al., 2006), 孤立的柱状叠层石主要发育在水体较深的潮下带, 具分支的柱状叠层石以及穹形叠层石多沉积于潮间带, 纹层状和波状叠层石多见于潮上带(钱迈平, 1991; Guhey et al., 2011)。凝块石的沉积水体深度普遍大于叠层石, 主要发育于潮下带(Burne and Moore, 1987; Feldmann and McKenzie, 1998; Ezaki et al., 2003; Mancini et al., 2004; Rezende et al., 2013)。已有报道中, 微生物碳酸盐岩普遍为快速海侵背景下高位体系域的沉积产物, 且具向上变浅的序列结构(Ezaki et al., 2003), 表现为叠层石和微生物粘结砂屑岩覆盖于凝块石或颗粒滩之上, 构成丘滩复合体(李凌等, 2013; 熊鹰等, 2016)。因此, 微生物岩产状、组合形式的纵向演变, 是研究相对海平面升降的有效材料。
鄂尔多斯盆地在马五段沉积时期为典型的局限— 蒸发海台地环境(史基安等, 2009; 张永生等, 2015), 表现为总体的海退背景下存在多个多周期的次级海平面振荡, 其中马五5亚段沉积时期为海侵环境。结合岩性、单旋回结构、多旋回组合形式等方面的垂向演变发现, 在上升半旋回, 微生物岩附着于初始海泛的低能沉积物之上, 随后由下至上依次出现层状凝块石/粘结砂屑→ 格架状凝块石→ 瘤状凝块石(图 4-a, 4-b); 产状的变化表现为层状向格架状的演变, 这表明沉积可容空间逐渐增大, 而凝块间泥晶充填物自格架状向瘤状凝块间增多, 则表明水动力条件的减弱, 即水体逐渐加深的过程。下降半旋回中, 瘤状凝块向上变为微生物团块(图 4-a, 4-b), 推测水动力条件增强, 并出现微生物组构搅碎现象, 指示水体逐渐变浅。整体来看, 研究区微生物岩的单旋回结构样式的变化, 呈现出沉积水体变深再变浅的演化趋势。此外, 2套旋回的组合形式中, 尤其在东部可见下部单旋回以发育叠层石为主, 上部单旋回则沉积瘤状凝块石(图 4)。多旋回组合形式的改变与单旋回中结构样式的演变一致, 不仅体现了不同级次旋回的自相似性, 而且表现出多旋回向上缓慢持续海侵的过程。
综上所述, 单旋回变深复变浅、多旋回持续缓慢海侵的相对海平面变化趋势, 是以瘤状凝块为特色的微生物岩垂向组合模式的重要驱动因素; 同时由单序列和多序列组合中微生物岩形态演变推导出的相对海平面演化趋势, 证实了马五5亚段较大规模的海侵沉积背景(黄正良等, 2015)。文中将此过程进一步细化, 定义为振荡式持续缓慢海侵。
1)下伏蒸发岩填平补齐的沉积前古地貌是微生物岩盆地尺度大规模发育的根本原因。受营养供给、光照条件等生命活动所需生态条件的限制(Rezende et al., 2013), 微生物岩常分布在台地边缘、台内点滩等水下高地或盐度异常、间接暴露的极浅水潮间带等环境(Kennard and James, 1986; Burne and Moore, 1987; 刘静江等, 2016; 徐欣等, 2018), 因此其平面分布规模较为有限(Shapiro, 2000; Webby, 2002; 周进高等, 2017)。相较之下, 研究区马五5亚段下部发育的2期微生物碳酸盐岩具盆地尺度的等时分布规模(图 6, 图 7, 图 8), 这在地质历史时期众多微生物岩的研究实例中较为罕见。微生物对繁殖环境的优选性及盆地尺度分布的现象, 表明全盆范围均为适宜微生物发育的场所, 暗示沉积前的地貌在全盆范围内差异不大。马家沟组沉积时期, 鄂尔多斯盆地继承了中央古隆起、吕梁水下隆起、靖边凸起等一系列寒武纪古构造单元(邵东波等, 2019; 周进高等, 2020; 魏柳斌等, 2021), 表现为隆坳相间的不平坦古地貌格局; 另一方面, 马五5亚段沉积前, 盆地内广泛沉积了若干套平面分布广、厚度规模大的蒸发岩(胡安平等, 2019; 左洺滔等, 2021), 由于蒸发盐具极高的沉积速率, 填平补齐的沉积作用在规模性蒸发岩发育层位普遍可见(谭秀成等, 2014; 王天云等, 2020), 因此推测在马五5亚段沉积前, 盆地被巨厚膏盐岩填平补齐, 沉积中心与沉降中心重合, 盆内原隆坳相间的古地形逐渐趋于平缓, 故下伏蒸发岩填平补齐的沉积前古地貌是控制马五5亚段下部微生物碳酸盐岩盆地尺度发育的根本原因(图 9-a)。
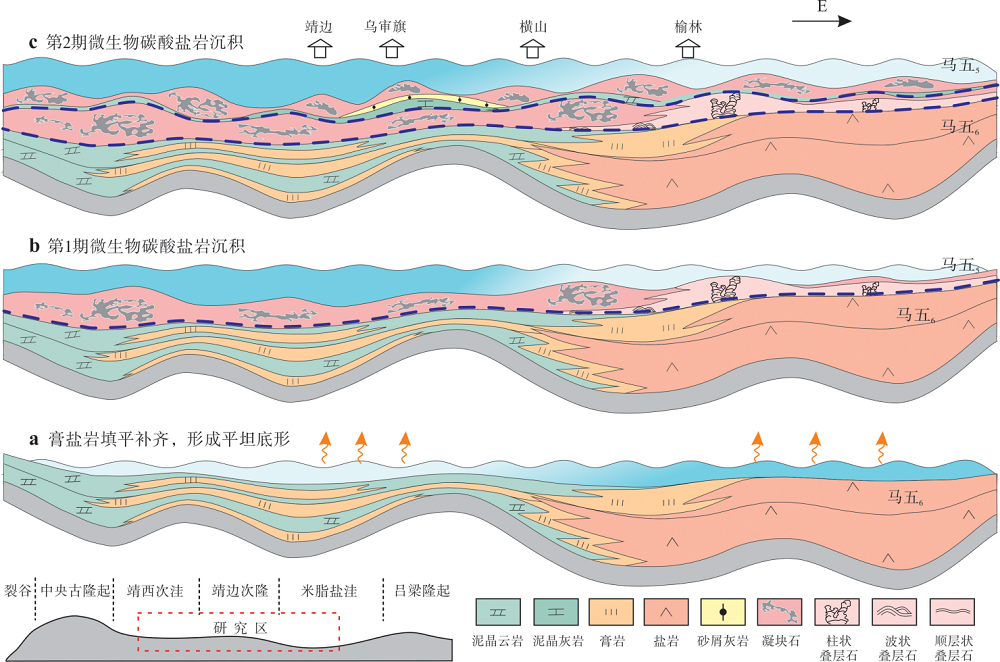 | 图 9 鄂尔多斯盆地中东部奥陶系马家沟组马五5亚段盆地尺度微生物碳酸盐岩沉积演化Fig.9 Depositional model of basin-scale microbial carbonates of the Ma55 sub-member of Ordovician Majiagou Formation in central and eastern Ordos Basin |
2)沉积期平静的古构造背景是产生微生物碳酸盐岩大面积均一化现象的必要条件。研究区内马五5亚段地层厚度稳定(图 1-c), 推测沉积期无剧烈的构造活动, 因而微生物碳酸盐岩沉积初期, 盆地构造活动相对平静, 古地理格局稳定, 有助于微生物岩持续连片发育, 这也是陆表海台地发生沉积填平补齐作用的先决条件。若此时构造活跃, 将引起地貌隆坳差异增大, 在持续海侵期沉积相和环境易产生分异, 仅会在局部水下高地分散发育微生物点丘, 不利于形成大面积分布的微生物碳酸盐岩。由此可见, 海侵背景下环境均一化的平坦沉积底形是规模性浅水微生物碳酸盐岩形成的根本条件。
3)东西向微生物碳酸盐岩发育产状与组合形式的差异暗示马五5亚段沉积期具西低东高的沉积期古地貌。盆地尺度分布的微生物碳酸盐岩是下伏蒸发岩填平补齐作用和相对平静的构造运动耦合的结果, 但微生物碳酸盐岩发育的产状和组合形式进一步反映了差异性沉积地貌。具体表现为: (1)西部以凝块沉积序列和连续沉积为主(图 4-a, 4-b), 向东演变为叠层石沉积序列和间断沉积(图 4-c, 4-d), 揭示出海水向东变浅(Ezaki et al., 2003; Guhey et al., 2011); (2)微生物碳酸盐岩旋回组合在西部由五级旋回构成, 而东部表现出六级旋回特征, 并且西部单旋回厚度大于东部(图 6); (3)马五5亚段地层厚度分布稳定, 呈西略厚、东略薄的特征(图 1-c)。结合马五5亚段下部等时地质体呈西厚东薄的变化特征(图 8), 表明盆地尺度下呈西略低、东略高的沉积期地貌差异, 从侧面揭示了该构造古地貌最晚始于马五5亚段沉积时期。
综上所述, 在其他发育条件都相同的情况下, 蒸发岩填平补齐作用和相对平静的构造运动耦合控制了稳定平坦底形(图 9-a)。持续缓慢海侵期, 底形沉积环境同质化, 微生物群落固定于低能的或较硬的沉积物上(温志峰等, 2004)并向四周衍生, 发育规模性展布的微生物碳酸盐岩(图 9-b)。微生物碳酸盐岩建造具水下正地貌特征, 导致沉积地貌产生微弱差异分化, 同时缓慢海侵期可容空间较小, 促使第2期微生物碳酸盐岩向局部高地的坡折处叠置迁移(图 9-c), 最终于马五5亚段下部形成盆地尺度的微生物碳酸盐沉积体。
1)鄂尔多斯盆地马家沟组马五5亚段下部发育一套盆地尺度的微生物碳酸盐岩组合, 包括叠层灰(云)岩、凝块灰(云)岩、粘结砂屑灰(云)岩及微生物团块灰岩等, 其中瘤状凝块灰岩发育频率最高。
2)垂向上, 发育4种与微生物碳酸盐岩密切相关的、向上变深再变浅的典型沉积序列, 微生物碳酸盐岩发育段单旋回具变深复变浅、多旋回持续缓慢海侵的特征。
3)横向上, 2期微生物碳酸盐岩稳定连片分布, 具盆地尺度的可对比性和等时性。微生物碳酸盐岩的累积厚度一般在3~7m, 整体呈向东减薄的趋势, 盆内连续分布面积可超过2.5× 105km2。
4)缓慢海侵背景下, 下伏蒸发岩填平补齐作用以及稳定构造环境形成了相对平坦古地貌, 环境同质化的底形条件造就了马五5亚段下部浅水微生物碳酸盐岩盆地尺度发育的独特沉积现象, 为碳酸盐岩台地微生物岩规模性发育机制的研究提供了实际案例。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|