





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
埃迪卡拉系微生物碳酸盐岩沉积特征及其地质意义:以川中磨溪8井区灯影组四段为例*
[罗垚1, 2  , 谭秀成
, 谭秀成1, 2 , 赵东方1, 2 , 罗文军3 , 刘耘3 , 肖笛1, 2 , 乔占峰4 , 曾伟1, 2 ]
, 谭秀成, 赵东方|
|

通讯作者简介 谭秀成,男,1970年生,教授,博士生导师,主要从事碳酸盐岩沉积学与储层地质学研究。E-mail: tanxiucheng70@163.com。
第一作者简介 罗垚,女,1996年生,西南石油大学硕士研究生,地质学专业。E-mail: landy0330@foxmail.com。
四川盆地高石梯—磨溪地区埃迪卡拉系灯影组广泛发育一套微生物碳酸盐岩。以磨溪 8井区灯影组四段(灯四段)为例,通过精细解析高频向上变浅序列的岩石学特征,发现灯四段微生物云岩及序列建造具有如下特征: ( 1)向上变浅序列主要由凝块石云岩、叠层石云岩和(含微生物)泥晶云岩 3类构成,伴以少量微生物粘结颗粒云岩;( 2)完整的典型向上变浅序列自下而上由(含微生物)泥晶云岩( A段)、平直状叠层石云岩( B段)、小型丘状叠层石云岩( C段)、分散状凝块石云岩( D段)、格架状凝块石云岩( E段)、微生物粘结颗粒云岩( F段)等 6个岩性单元构成,但大量的序列由 A- C- E、 A- C- D、 B- D、 B- C- D- E等不完整的岩性单元构成,且序列顶底皆以凝块石云岩 /(含微生物)泥晶云岩、叠层石云岩 /(含微生物)泥晶云岩等不平整的岩性突变面或暴露面区分;( 3)埃迪卡拉系微生物碳酸盐岩发育于浪基面之上至平均海平面附近的相对高能区域,且凝块石云岩较叠层石云岩形成的环境能量高。该研究结果不仅揭示出较高能的浅水环境控制了规模性微生物岩分布这一规律,而且对区域储集层预测具有指导意义,同时也因建立了有别于现代微生物的环境分布模式而具有重要的沉积学意义。
, Tan Xiu-Cheng, Zhao Dong-FangAbout the corresponding author Tan Xiu-Cheng,born in 1970,is a professor and Ph.D. advisor of Southwest Petroleum University. He is mainly engaged in researches on carbonate sedimentology and reservoir geology. E-mail: tanxiucheng70@163.com。
About the first author Luo Yao,born in 1996,is a master degree candidate of Southwest Petroleum University. She majors in geology. E-mail: landy0330@foxmail.com。
The Ediacaran microbial carbonate rocks are widely developed in the Dengying Formation in the Gaoshiti-Moxi area of the Sichuan Basin. This paper takes the Member 4 of the Dengying Formation in Wellblock MX8 as an example. By analyzing the macroscopic and microscopic petrological characteristics of the high-frequency upward shallowing sequence,it is found that the microbial dolomite and sequence construction in the Member 4 of Dengying Formation have the following characteristics: (1)The upward shallowing sequence is mainly composed of thrombolites,stromatolites,microbial dolomicrite and a small amount of microbial bonded granular dolomite. (2)The complete typical upward shallowing sequence is composed of six lithologic units from bottom to top,including microbial dolomicrite(A),flat stromatolite(B),small hummocky stromatolite(C),dispersed thrombolite(D),latticed thrombolite(E),microbial bonded granular dolomite(F). A large number of sequences are composed of incomplete lithologic units such as A-C-E,A-C-D,B-D,B-C-D-E,and the top and bottom of the sequence are distinguished by uneven lithologic mutation surfaces or exposure surfaces such as thrombolites-microbial dolomicrite,stromatolites-microbial dolomicrite. (3)The Ediacaran microbial carbonates were developed in relatively high energy areas above the wave base to near the average sea level,and the environmental energy of the thrombolites are much higher than that of the stromatolites. The results reveal that the high-energy shallow water environment not only controls the distribution of large-scale microbial rocks,but also provides guidance for regional reservoir prediction. Meanwhile,the results are of great sedimentological significance in the establishment of an environmental distribution model that is different from modern microorganisms.
开放科学(资源服务)标志码(OSID) 
微生物岩广泛发育于地质历史的各个时期(Riding, 2002; Riding and Liang, 2005), 其成分多样, 包括钙质、铁锰质、磷质、硅质等(Centeno et al., 2012), 又以微生物碳酸盐岩最为普遍(杨孝群和李忠, 2018)。作为沉积环境和生物化学过程相互作用的产物, 微生物岩记录了古环境演化的关键信息(戴永定等, 1996; 吴亚生等, 2007)。从“ 微生物岩” (microbiolites)这一术语被正式提出以来(Burne and Moore, 1987), 地质工作者针对其类型划分(梅冥相, 2007; 吴亚生等, 2018)、形成过程(Riding, 2006; 韩作振等, 2009)、控制因素(Andres and Reid, 2006; Woo et al., 2008)、古海水化学特征(Webb and Kamber, 2000; Kamber et al., 2014)、环境与生物的协同演化(Allwood et al., 2007)等做了大量的工作。此外, 大量的研究实例表明, 微生物碳酸盐岩具有良好的油气勘探前景(李凌等, 2013; 刘树根等, 2016), 其生烃潜力(杨浩等, 2007; 佘敏等, 2019)和油气储集意义(罗平等, 2013; 余浩元等, 2018)也受到广泛关注。虽然针对微生物岩国内外学者已做了大量研究(Knoll et al., 2013; 罗冰等, 2015; 陈娅娜等, 2017), 但截至目前, 在微生物岩的相序特征及其沉积演化模式等方面依然存在争议(罗平等, 2013)。四川盆地埃迪卡拉系灯影组内微生物碳酸盐岩分布广泛、类型众多(方少仙等, 2003; 李凌等, 2013), 且高石梯— 磨溪地区灯影组钻井取心丰富, 这为微生物碳酸盐岩研究提供了良好的材料。鉴于此, 文章以川中磨溪8井区灯影组四段(灯四段)微生物碳酸盐岩为研究对象, 开展细致的岩石学宏、微观特征研究, 明确微生物碳酸盐岩类型及序列特征, 构建其发育模式, 以期为进一步丰富和完善不同时代的微生物岩沉积模式提供基础材料和研究实例。
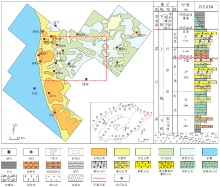
在埃迪卡拉纪(震旦纪), 扬子区构造背景主要为被动大陆边缘环境(Jiang et al., 2003)。南沱冰期后, 随着陡山沱期气温的回升, 冰川大面积融化, 相对海平面迅速上升, 使得上扬子区发生大规模海侵(柳永清等, 2003; 余谦等, 2011; 张亚冠等, 2015), 以残余古陆、滨岸— 潮坪、陆棚和盆地沉积为主, 岩性主要为白云岩、云质灰岩和泥页岩, 厚100~200 m。至灯影期, 沉积基准面总体上西高东低, 坡度极缓, 自南西向北东水体逐渐加深(李英强等, 2013; 刘静江等, 2015; 赵东方等, 2018), 四川盆地及周缘以发育大规模碳酸盐岩台地建造为特色, 但受桐湾运动影响, 该套地层遭受了不同程度的剥蚀, 厚度变化较大, 薄至50 m, 厚可高达1400 m。灯影组自下而上可分为4个岩性段: 灯一段以泥— 粉晶云岩为主, 夹少量微生物云岩; 灯二段和灯四段主要为微生物云岩和泥— 粉晶云岩, 夹颗粒云岩(Zhao et al., 2020); 灯三段则多呈现混积特征, 主要发育泥质粉砂岩、泥质云岩, 局部发育砂岩或砂质砾岩(图 1)。本次主要研究对象为磨溪地区灯影组四段, 区域构造上位于川中古隆中斜平缓带, 以发育碳酸盐岩台地微生物丘为主。
 | 图 1 川中高石梯— 磨溪地区埃迪卡拉纪灯影组四段沉积时期岩相古地理及灯影组岩性特征Fig.1 Lithofacies palaeogeography during the depositional stage of Member 4 of Ediacaran Dengying Formation and its lithologic characteristics in Gaoshiti-Moxi area of central Sichuan Basin |
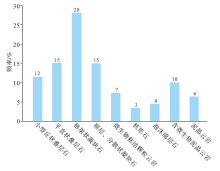
对于微生物碳酸盐岩分类, 学者们已提出了多种分类方案(Burne and Moore 1987; Shapiro, 2000; Riding, 2002; 梅冥相, 2007), 但由于分类依据不同, 造成微生物岩的分类方案分歧较大。在川中磨溪地区灯影组已识别出多种构成微生物丘建造的岩石类型, 笔者以岩心宏观结构和镜下微观组构的特征为分类依据, 将其分为5大类10小类, 主要为叠层石云岩类、凝块石云岩类、(含微生物)泥晶云岩类以及微生物粘结颗粒云岩类, 还含有少量的核形石及泡沫绵层石(表 1, 图 2)。
| 表 1 川中磨溪地区埃迪卡拉系灯影组岩石类型 Table1 Rock types of the Ediacaran Dengying Formation in Moxi area of central Sichuan Basin |
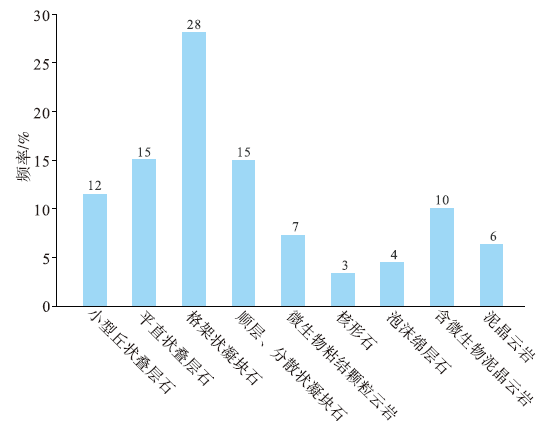 | 图 2 川中磨溪地区埃迪卡拉系灯影组岩石类型发育频率Fig.2 Development frequency of rock types of the Ediacaran Dengying Formation in Moxi area of central Sichuan Basin |
川中磨溪地区灯影组叠层石云岩按其纹层形态可分为平直状和小型丘状, 因此, 笔者以岩心上宏观组构的形态对其进行分类和命名。鉴于岩心尺度有限, 平直状叠层石可能在野外宏观上为丘状叠层石。
平直状叠层石云岩具纹层结构。岩心上纹层大多整体光滑, 呈平直状— 微波状(图 3-a), 单个纹层厚度多变, 介于1~10 mm之间, 纹层横向延续性好, 可大致追踪。显微镜下, 明暗层间的界线多呈低幅度起伏, 局部呈渐变的特征(图 3-b, 3-c)。暗层为富有机质层, 主要由暗色微生物组构和泥晶白云石构成。亮层为基质层, 贫有机质, 晶粒稍粗, 基本为泥—粉晶白云石。
 | 图 3 川中磨溪8井区埃迪卡拉系灯影组四段叠层石云岩类型 a— 平直状叠层石云岩, 岩心照片, 磨溪21井, 5064.45~5064.51 m; b— 平直状叠层石云岩, 岩心照片, 磨溪123井, 5478.31 m; c— 平直状叠层石云岩, 单偏光, 磨溪105井, 5363.40 m; d— 丘状叠层石云岩, 岩心照片, 磨溪21井, 5079.42~5079.51 m; e— 丘状叠层石云岩, 单偏光, 磨溪108井, 5300.13 m; f— 丘状叠层石云岩, 单偏光, 磨溪123井, 5479.21 mFig.3 Types of stromatolite dolomite in the Member 4 of Ediacaran Dengying Formation in Wellblock MX8 of central Sichuan Basin |
小型丘状叠层石云岩在岩心及镜下均可见明暗相间的纹层结构, 纹层形态呈明显的丘状起伏(图 3-d)。单个丘的高度虽多变, 但总体规模不大, 多为分米级, 整体呈半球状或宽缓状产出。单个丘内纹层厚度具明显的差异, 薄仅1 mm, 厚者近于1 cm。相较于平直状叠层石, 小型丘状叠层石纹层的横向连续性较差。微观下, 小型丘状叠层石的纹层多具有明显的局部形变特征, 呈揉皱起伏状(图 3-e, 3-f), 并在纹层内部形成类似小型格架状的原生孔洞, 孔洞多被粉— 细晶白云石胶结充填。
叠层石云岩的形态结构与沉积环境的水动力条件密切相关(Logan et al., 1964; 钱迈平, 1991; 朱士兴, 1993; 钱迈平等, 2002; 范正秀等, 2018)。平直状叠层石中纹层呈平直状分布, 指示相对静水的低能环境; 小型丘状叠层石中纹层形态呈揉皱起伏状, 规模不大, 为中等水动力条件下的产物。
基于凝块组构形态多变, 故将川中磨溪地区灯影组凝块石云岩按其组构形态分为3种, 分别为格架状、顺层状和分散状凝块石云岩。
分散状凝块石云岩具凝块结构, 凝块组构富有机质, 色暗。单个凝块普遍较小, 多呈不规则的小型斑点状(图 4-a), 个别凝块稍大, 以微型团块集合体产出(图 4-b)。凝块含量整体不高, 但在基质中的分布频率却有较大差异, 局部可见分散凝块富集产出、横向延续性极差, 具明显的非均质性。凝块多呈孤立状随机漂浮于泥—粉晶白云石基质中, 构成明显的基质支撑结构。凝块与基质间的界限相对模糊, 呈现由凝块到基质渐变浸染特征。
 | 图 4 川中磨溪8井区埃迪卡拉系灯影组四段凝块石云岩类型及特征 a— 分散状凝块石云岩, 岩心照片, 磨溪123井, 5484.60~5493.89 m; b— 分散状凝块石云岩, 单偏光, 磨溪105井, 5484.60 m; c— 顺层状凝块石云岩, 岩心照片, 磨溪17井, 5067.61 m; d— 顺层状凝块石云岩, 单偏光, 磨溪123井, 5477.4 m; e— 格架状凝块石云岩, 格架孔发育且后期充填多期次白云石胶结物, 单偏光, 磨溪125井, 5317.91 m; f— 格架状凝块石云岩, 岩心照片, 磨溪125井, 5320.12~5320.18 m; g— 格架状凝块石云岩, 单偏光, 磨溪51井, 5383.27 m; h— 格架状凝块石云岩, 凝块内部可见粘结的颗粒发亮红光, 阴极发光, 磨溪51井, 5383.27 mFig.4 Types and characteristics of thrombolite in the Member 4 of Ediacaran Dengying Formation in Wellblock MX8 of central Sichuan Basin |
顺层状凝块石云岩具不规则条带状结构。与叠层石云岩相似, 都发育“ 明暗条带” , 但叠层石云岩“ 条带” 厚度均匀, 横向连续性好, 而顺层状凝块石云岩中的条带形态很不规则, 即使是同一条带, 其厚度变化也存在较大差异, 横向可追踪性差(图 4-c)。单个条带是由多个凝块组成的集合体, 呈明显的定向性(图 4-d), 内部形成多个大小不一的似层状原生孔洞。单个凝块呈狭长的不规则暗斑产出, 局部相互连接缠绕形成小规模格架状团块。
格架状凝块石云岩具凝块— 格架状结构。凝块含量高, 宏观上可见浅色基质呈点状或带状分布于深灰色凝块集合体中(图 4-f), 镜下可见凝块相互聚集呈团块状集合体, 部分团块内部可见生物粘附作用形成的颗粒(图 4-g)。这些微生物团块进一步粘结缠绕生长、相互连接, 在三维空间中构成网状格架(图 4-e), 内部形成格架状孔洞, 并被多期胶结物充填。阴极发光下, 格架状结构明显可见, 凝块格架富有机质, 发光性较强, 呈相对均匀的红色, 基质发暗红色光或不发光。特别是微生物粘结颗粒在阴极发光下非常明显(图 4-h), 这些颗粒大小不一, 发光性相较于凝块格架更为显著, 呈亮红色光。
微生物岩的形貌特征与沉积相带的水动力条件有较好有相关性(Mettraux et al., 2014)。对于凝块组构而言, 凝块呈分散状在基质中生长表明其水动力并不强, 顺层状则表明其需要一定的水动力条件使其呈顺层状排列, 而格架状则以凝块间相互连接构成具有一定抗浪能力的格架体系, 且凝块组构内部可见粘结颗粒结构(Riding, 2002), 表明其是在较强的水动力条件下形成的。
岩心上, 纯净的颗粒云岩与微生物粘结颗粒云岩有时难以区分, 但相较于前者, 微生物粘结颗粒云岩有机质含量更高, 故颜色一般更暗(图 5-a)。微观分析显示, 粘结颗粒色暗, 大多颗粒并不直接接触, 接触仅局限在微生物组构内部, 整体呈漂浮状产出, 具明显的微生物包覆特征。颗粒主要为内碎屑, 多为早期形成的微生物岩被波浪打碎后再沉积而成的, 内部可见微生物组构(图 5-b, 5-c)。颗粒之间可见明显的微生物粘结痕迹, 构成颗粒支撑。颗粒间填隙物以亮晶白云石为主, 溶孔发育。
 | 图 5 川中磨溪8井区埃迪卡拉系灯影组四段其他岩石类型及特征 a— 微生物粘结颗粒云岩, 岩心照片, 磨溪17井, 5037.85 m; b— 微生物粘结颗粒云岩, 单偏光, 磨溪105井, 5323.94 m; c— 微生物粘结颗粒云岩, 单偏光, 磨溪108井, 5332.11 m; d— 泡沫绵层石云岩, 单偏光, 磨溪125井, 5317.82 m; e— 核形石云岩, 单偏光, 磨溪105井, 5303.52 m; f— 含微生物泥晶云岩, 单偏光, 磨溪125井, 5380.89 mFig.5 Other rock types and characteristics of the Member 4 of Ediacaran Dengying Formation in Wellblock MX8 of central Sichuan Basin |
研究区还发育少量泡沫绵层石和核形石等与微生物相关的微生物云岩。
泡沫绵层石具绵层结构。微观下, 微生物组构或以独立球状、椭球状、不规则状的“ 泡沫空腔” 产出, 或相互粘结聚集, 呈不规则泡棉状网络(图 5-d)。单个泡沫大小不一, 介于0.05~0.4 mm之间, 多为不规则圆形。泡沫之间为原生格架孔洞, 可见多期次白云石胶结充填。
核形石整体色暗, 具包覆颗粒结构。其大小不一, 形态多样, 呈椭球状、纺锤状(图 5-e)。核形石由内部的核心和外部的包壳组成, 内部可见微生物组构呈团块状分布。核形石之间的格架孔被多期次白云石所充填。
研究区泥晶云岩有2种, 分别为含微生物泥晶云岩和泥晶云岩。虽然含微生物泥晶云岩不属于典型的微生物白云岩类型, 但由于发育少量的微生物组构而又明显有别于纯净的泥— 粉晶云岩。岩心上, 含微生物泥晶云岩常与泥— 粉晶云岩伴生, 镜下呈致密的泥— 粉晶结构, 可见少量有机质组构呈不太明显的凝絮状或纹层状分布(图 5-f)。泥晶云岩则不具微生物痕迹, 呈纯净的泥— 粉晶结构。
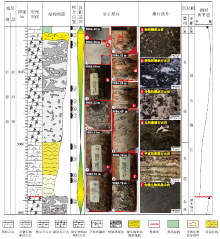
川中磨溪8井区灯影组四段可识别出多个与微生物丘密切相关的向上变浅序列, 其单个旋回厚度不大, 以米级为主, 在纵向上频繁叠置。核形石和泡沫绵层石发育频率低(图 2), 在微生物丘序列建造上不具普遍性。因此, 向上变浅序列主要由(含微生物)泥晶云岩、叠层石云岩和凝块石云岩3类构成, 伴以少量粘结颗粒云岩。完整的典型向上变浅序列自下而上由(含微生物)泥晶云岩(A段)、平直状叠层石云岩(B段)、小型丘状叠层石云岩(C段)、分散状凝块石云岩(D段)、格架状凝块石云岩(E段)、微生物粘结颗粒云岩(F段)等6个岩性单元构成(图 7)。在沉积的单个微生物丘序列中, 受相对海平面波动影响, 不一定会出现上述所有岩性组合类型, 可能是其中几类的纵向叠置, 如A-C-E(图 6-a中Ⅰ , 6-c)、A-C-D(图 6-a中Ⅱ )、B-D(图 6-a中Ⅲ )、B-C-D-E(图 6-b)、A-C(图 6-d中Ⅰ )、A-B-C(图 6-d中Ⅱ )和A-E(图 6-d中Ⅲ )等不完整的岩性单元构成。序列顶底皆以凝块石云岩/(含微生物)泥晶云岩、叠层石云岩/(含微生物)泥晶云岩等不平整的岩性突变面或暴露面区分。
 | 图 6 川中磨溪21井5084.4~5087.0 m段沉积序列综合柱状图Fig.6 Comprehensive histogram of 5084.4~5087.0 m sedimentary sequence from Well MX21 in central Sichuan Basin |
高石梯— 磨溪地区灯影组四段沉积期古地理背景以浅水碳酸盐岩台地为主(图 1)(陈娅娜等, 2017; 周进高等, 2017), 其微生物岩建造的组合类型及规模主要为在相对海平面升降引起的水体能量变化下的响应。
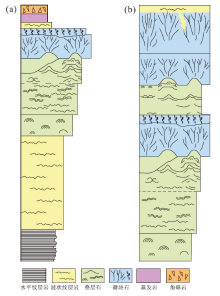
上述已识别出的灯影组四段单个沉积旋回的岩石组合类型、厚度以及纵向上多个沉积旋回的频繁叠置(图 6), 不仅与相对海平面的频繁升降有关, 还与相对海平面波动下微生物丘建造的叠置迁移密切相关(李凌等, 2013)。根据微生物丘垂向发育特征, 总结归纳出研究区灯影组四段纵向上完整的向上变浅序列(图 7)和理想的微生物丘沉积模式(图 8)。
 | 图 7 川中磨溪8井区埃迪卡拉系灯影组四段不同岩性组合沉积旋回频繁叠置Fig.7 Sedimentary cycles of different lithologic assemblages frequently superimposed in the Member 4 of Ediacaran Dengying Formation in Wellblock MX8 of central Sichuan Basin |
 | 图 8 川中磨溪8井区埃迪卡拉系灯影组四段微生物丘沉积模式Fig.8 Microbial mound depositional model of the Member 4 of Ediacaran Dengying Formation in Wellblock MX8 of central Sichuan Basin |
微生物丘纵向上可分为丘基、丘核、丘盖微相: 丘基微相位于微生物丘底部, 是微生物发展的初期和拓殖阶段, 微生物较缺乏, 以发育泥晶云岩、含微生物泥晶云岩和叠层石云岩为主, 是初始海平面扰动背景下的产物; 丘核微相形成于浪基面之上至平均海平面附近的相对高能区域, 以垂向加积为主, 微生物生长开始繁盛, 进入微生物发展的泛殖阶段, 此时水体能量较高, 沉积速率较快, 微生物粘结、缠绕生长, 形成具有一定抗浪能力的格架系统, 发育凝块石云岩和微生物粘结颗粒云岩; 丘盖微相位于微生物丘的顶部, 是微生物发展的衰亡阶段, 此时受海侵影响沉积水体加深, 以发育贫微生物的泥晶云岩为主。由于研究区相对海平面波动频繁, 往往不会出现丘盖微相, 或下一期微生物丘的丘基微相为上一期的丘盖微相。
针对微生物碳酸盐岩的发育模式, 国内外学者做了大量工作(朱士兴, 1993; Jahnert and Collins, 2012), 但截至目前, 大多数学者都把叠层石(钱迈平, 1991; Guhey et al., 2011; 范正秀等, 2018)和凝块石(Mancini et al., 2004)分开讨论。由于对前寒武纪微生物岩(尤其是叠层石和凝块石)的沉积模式缺乏系统的研究, 学者们常通过借鉴现代微生物岩实例来讨论古老深层微生物碳酸盐岩的沉积模式(黄擎宇, 2016; 王文之, 2016)。但需要注意的是, 现代微生物岩沉积环境与元古代差别巨大(Lyons et al., 2009, 2014; Zhang et al., 2016), 因此是否能用“ 将今论古” 的思维将现代沉积模式沿用于前寒武纪还有待进一步证实。
笔者通过大量岩心及镜下观察发现, 在研究区发育的向上变浅序列中, 叠层石皆发育于凝块石下部(图 6, 图 7)。国内外学者报道的中新元古代实例也能证实这一点, 如梅冥相(2014)将天津蓟县剖面雾迷山组划分为若干个米级旋回, 但自下而上都是以泥晶白云岩/泥质白云岩— 叠层石生物层— 凝块石生物丘构成; 从阿曼Qarn Alam地区新元古代野外露头剖面的多个相序组合中可以看到, 单个序列都是先沉积叠层石再发育凝块石(Mettraux et al., 2014)(图 9)。
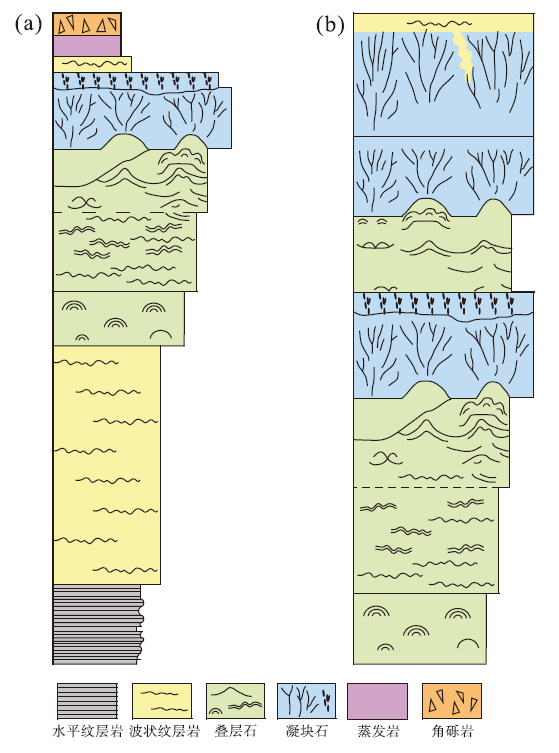 | 图 9 阿曼Qarn Alam地区新元古代微生物岩野外露头相序组合类型及特征(据Mettraux et al., 2014; 有修改) a— 向上变浅序, 岩性由下至上依次为: 水平纹层岩— 波状纹层岩— 叠层石— 凝块石— 纹层岩— 蒸发岩— 角砾岩; b— 由2套向上变浅序列组合而成, 岩性由下至上依次为: 叠层石— 凝块石— 波状纹层岩— 叠层石— 凝块石— 波状纹层石Fig.9 Types and characteristics of outcrop facies sequence assemblages of the Neoproterozoic microbialites in Qarn Alam area, Oman(modified from Mettraux et al., 2014) |
而现代沉积实例证实, 凝块石主要形成于水体较深的潮下带环境, 例如, 对巴哈马和鲨鱼湾地区微生物岩的研究表明, 凝块石多发育于相对开阔的浅潮下带环境(Burne and Moore, 1987; Jahnert and Collins, 2012)。鉴于此, 笔者认为导致埃迪卡拉纪叠层石发育在凝块石下部的主要原因可能是: (1)有研究表明, 中元古代丝状蓝细菌多生长在球状蓝细菌的下部(Knoll et al., 2013), 并且凝块结构与球形菌占主导的微生物群落的同期生长和钙化作用相关(Aitken et al., 1989), 而在叠层石中丝状蓝细菌占主导地位(Awramik, 1971); (2)前寒武纪和显生宙沉积底质的巨大差异, 可能暗示微生物岩发育模式与现今并不一致, 前寒武纪为微生物席底(Seilacher, 1999), 无后生动物扰动, 底质具黏性, 微生物席与水流相互作用形成大量缠绕、粘结的包覆结构, 而现代为混合底质, 后生动物的扰动强, 沉积物层容易被水流打散; (3)前寒武纪海水的化学性质与显生宙存在明显差异(Sandberg, 1983; Lowenstein et al., 2003; Dickson, 2004), 可能也是导致微生物岩发育模式不同于显生宙的原因。由于缺乏直接的微生物证据, 并要经历了强烈的成岩作用, 因此上述推论还有待今后进一步研究才能得到合理的阐释。
综上所述, 依据现代观察建立的微生物碳酸盐岩发育模式并不适用于前寒武纪, 埃迪卡拉纪微生物碳酸盐岩沉积发育模式还需进一步厘定。此外, 在叠层石和凝块石共存的沉积环境中, 两者发育的水体深度并无定论, 应具体情况具体分析。
微生物碳酸盐岩作为一类重要的储集层, 随着国内外油气勘探实践的深入已得到证实, 如美国墨西哥湾侏罗系(Mancini et al., 2004)、阿曼盐盆上震旦统— 下寒武统“ Ara群” (Hudec and Jackson, 2007)、东西伯利亚的中上元古界至下寒武统(Tull, 1997)和中国渤海湾盆地中元古界雾迷山组(费宝生和汪建红, 2005)等, 都发现了以微生物岩为储集层的大型油气田。
四川盆地灯影组储集层中微生物岩是重要的储集岩类型, 其储集意义已得到广泛认可(李凌等, 2013; 赵文智等, 2015; 刘树根等, 2016)。磨溪8井区灯影组四段的原生孔洞大多被破坏, 储集空间类型以次生为主, 包括凝块间扩溶格架溶孔(图 10-c)、微生物粘结颗粒残余粒间孔(图 10-b)、纹层间残余溶孔(图 10-e, 10-f)以及具微生物粘结组构间的顺层残余溶孔(图 10-a, 10-d), 表明与微生物岩相关的储集空间类型多表现为组构间的选择性溶蚀(图 10)。
 | 图 10 川中磨溪8井区埃迪卡拉系灯影组四段微生物碳酸盐岩储集空间类型 a— 格架状凝块石云岩, 凝块间发育顺层状残余溶蚀孔(洞), 岩心照片, 磨溪105井, 5372.16~5372.24 m; b— 微生物粘结颗粒云岩, 发育粒间溶孔且被沥青半充填, 单偏光, 磨溪105井, 5325.68 m; c— 格架状凝块石云岩, 凝块间发育残余格架溶孔, 单偏光, 磨溪51井, 5341.2 m; d— 叠层石云岩, 顺层状溶蚀孔洞, 岩心照片, 磨溪125井, 5378.10~5378.32 m; e— 丘状叠层石云岩, 纹层间发育残余溶孔, 单偏光, 磨溪125井, 5378.40 m; f— 平直状叠层石云岩, 纹层间发育残余溶孔, 单偏光, 磨溪105井, 5320.07 mFig.10 Microbial carbonates reservoir space types in the Member 4 of Ediacaran Dengying Formation in Wellblock MX8 of central Sichuan Basin |
灯影组优质储集层的发育主要与微生物丘高频暴露及岩溶作用造成的不同程度改造有关。现今保存的优质储集层主要集中于单个向上变浅序列的中上部, 虽单个旋回厚度不大, 以米级旋回为主, 但纵向上的频繁叠置迁移呈现出“ 薄层多套” 的特征, 累计厚度可观, 具良好的相控性。据此认为, 微生物丘是灯影组储集层发育的物质基础, 与微生物丘相关的沉积微地貌高地是灯影组储集层发育的有利区带。
1)四川盆地高石梯— 磨溪地区埃迪卡拉系灯影组四段发育的与微生物丘建造相关的岩类可分为5大类10小类, 分别是: (1)平直状叠层石; (2)小型丘状叠层石; (3)分散状凝块石; (4)顺层状凝块石; (5)格架状凝块石; (6)微生物粘结颗粒云岩; (7)泡沫绵层石; (8)核形石; (9)含微生物泥晶云岩; (10)泥晶云岩。研究区的向上变浅序列均由以上岩类有序组合构成, 且单个旋回厚度不大, 以米级为主。
2)磨溪8井区灯影组四段微生物丘发育于碳酸盐岩台地相区, 不同类型微生物岩的形成与水动力条件强弱有良好的相关性, 且在相应的沉积环境中大致呈分带性分布。在此基础上, 建立了该时期的微生物碳酸盐岩沉积演化模式: 丘基以发育泥晶云岩、含微生物泥晶云岩和平直状叠层石云岩为主; 丘核以发育小型丘状叠层石、凝块石云岩和微生物粘结颗粒云岩为主。
3)埃迪卡拉纪微生物碳酸盐岩沉积模式有待进一步厘定。微生物碳酸盐岩的相序结构是沉积环境有序变化的结果, 不同时代的微生物群落对应的岩相相序有其共性, 更有其特点。对不同时代的微生物岩类型的演化规律进行研究, 并建立其对应的沉积模式, 将有助于沉积微相的厘定及岩相古地理的精细刻画。
4)磨溪8井区灯影组四段储集层类型以孔洞型为主, 储集层分布具有明显的相控性和旋回性, 优质储集层主要发育于向上变浅序列旋回的中上部, 纵向上频繁叠置, 薄层多套, 累积厚度大。据此, 微生物丘可作为灯影组储集层发育的物质基础, 寻找与微生物丘相关的微地貌高地对灯影组储集层有利区带的预测有一定指导意义。
(责任编辑 张西娟; 英文审校 刘贺娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|