


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
东秦岭陕西聂家沟剖面二叠纪—三叠纪之交沉积特征及其古环境意义*
[李飞洋1, 2  , 张立军
, 张立军1, 2 , 李泰然1 , 杨琦琦1 , 牛永斌1, 2 , 宋慧波1, 2 ]
, 张立军, 李泰然|
|

通讯作者简介 张立军,男, 1982年生,河南理工大学资源环境学院副教授,主要从事沉积学、遗迹学等研究。 E-mail: Ljzhanghpu@gmail.com。
第一作者简介 李飞洋,男, 1993年生,河南理工大学资源环境学院硕士研究生,从事沉积学研究。 E-mail: 1169006628@qq.com。
东秦岭陕西镇安聂家沟地区二叠纪—三叠纪之交发育一套海相碳酸盐岩沉积序列,较完整地记录了二叠纪末生物大灭绝事件前后浅海碳酸盐岩台地相生物群落演替及环境变化等信息,是研究二叠纪末生物大灭绝事件前后生态环境变化与沉积响应的理想区域。通过碳酸盐岩微相分析方法,对陕西镇安聂家沟剖面二叠系—三叠系界线附近的碳酸盐岩生物组合和微相类型进行了详细分析,共识别出 11个微相类型: 斑点状凝块石、叠层石灰岩、鲕粒—纹层状叠层石灰岩、有孔虫颗粒灰岩、藻—海百合泥粒灰岩、含有被包壳和被磨蚀骨屑颗粒的粒泥灰岩、鲕粒颗粒灰岩、集合颗粒灰岩、含鲕粒的粒泥灰岩、泥晶灰岩和泥岩。根据碳酸盐岩微相特征及沉积相标志,在二叠系—三叠系界限附近划分出台地边缘、开阔台地和局限台地 3种沉积相,其反映了二叠纪—三叠纪之交频繁的沉积相带变更的特点。该剖面碳酸盐岩微相反映的古生态和生物群落演替特征与中国华南同时期其他剖面具有较好的一致性,即二叠纪末生物大灭绝事件之后,早三叠世正常浅水碳酸盐岩台地生物类型和丰度极低,仅含有少量的双壳类、海百合等,灭绝事件界线附近以微生物碳酸盐岩等特殊微生物沉积构造占主导,之后微生物岩等消失,又出现了以双壳类为首的生物碎屑石灰岩。研究结果为了解二叠纪末生物大灭绝事件中生物和环境的协同演化提供了新的材料和认识。
About the corresponding author ZHANG Lijun,born in 1982,an associate professor of Henan Polytechnic University,is engaged in ichnology and sedimentology. E-mail: Ljzhanghpu@gmail.com.
About the first author LI Feiyang,born in 1993,is a master degree candidate. He is engaged in sedimentology. E-mail: 1169006628@qq.com.
The marine carbonate strata of the Permian-Triassic transition were found in the Niejiagou section of Zhen'an,Shaanxi. It records the succession of biota and environmental changes of shallow-marine carbonate platform in the Permian-Triassic transition,and is a perfect area for studying the depositional response and palaeoenvironmental changes before and after the end-Permian mass extinction. Based on the detailed analysis of carbonate microfacies and biology combination in the Permian-Triassic transition of the Neijiagou section,11 microfacies types are identified,including patched agglomerates,stromatolites,oolitic-layered stromatolites,foraminiferal grainstone,algal-crinoid grainstone,wackestone containing coated and eroded bone debris particles,oolitic grainstone,aggregate grainstone,oolitic wackestone,mudstone and micrite. According to the characteristics of carbonate microfacies and sedimentary indicators,marginal shoal facies,open platform facies and restricted platform facies are divided in the Permian-Triassic transition. The microfacies characteristics reflect frequent sedimentary microfacies changes in the Permian-Triassic transition. The biological succession and stratigraphic sequence of the Permian-Triassic transition in the Niejiagou section have good uniformity correlation with other sections in South China. After the end-Permian mass extinction,low biodiversity and biological abundance,e.g.,small bivalves and crinoid,occur in the shallow-marine carbonate platform in the Early Triassic. The special microbial sedimentary structures dominated around the mass extinction event boundary. Afterwards,the microbialites disappeared and the bioclastic limestone reoccupied. This research can provide new understanding for the coevolution of organisms and the environment caused by the end-Permian mass extinction.
开放科学(资源服务)标志码(OSID) 
二叠纪— 三叠纪之交发生了显生宙以来规模最大的一次生物大灭绝事件, 导致了约81%海洋生物灭绝(Stanley, 2016; Fan et al., 2020)。古生代型生态系统在此时走到了尽头, 而新的生态系统尚未形成(殷鸿福和宋海军, 2013; 戎嘉余和黄冰, 2014; 宋海军和童金南, 2016)。二叠纪— 三叠纪之交作为研究生物与环境协同演化的关键转折期, 长期受到人们的关注, 已在生物灭绝的模式、过程以及机理上取得了一系列共识(Erwin, 1994; Kiessling and Simpson, 2011; Brand et al., 2012; Yin et al., 2012; Shen et al., 2019), 如认为温室效应叠加缺氧事件是导致二叠纪末生物大灭绝事件的主要原因(Isozaki, 1997; Brennecka et al., 2011; Meyer et al., 2011; Sun et al., 2012)。相较于众多的二叠纪— 三叠纪之交生物多样性和地球化学研究(Raup, 1979; Baud et al., 1989; 曹长群等, 2002; 王永标等, 2005; 吴亚生等, 2007; 宋海军等, 2012; Jin et al., 2014; Song et al., 2018; Liu et al., 2020), 该时期传统的高分辨率沉积学研究较为薄弱(赵俊兴等, 2003), 这也限制了从不同角度深入了解此次灭绝事件对于不同沉积相区的影响程度。通过对聂家沟剖面二叠纪— 三叠纪之交高分辨率沉积学的详细研究, 可以查明二叠纪末生物大灭绝事件在不同古地理区域的影响特征, 为了解二叠纪末生物大灭绝事件对于生物和环境的协同演化提供新的材料和认识。
“ 微相” 一词最早是由Brown(1943)定义的, 早期仅是在岩石薄片的鉴定中用来描述岩石学和古生物特征的术语。现今“ 微相” 一词已变成综合性术语, 指在岩石薄片、岩石揭片、剖光片中描述岩石沉积特征和古生物特征。由于碳酸盐岩成因和结构的特殊性, 使碳酸盐岩微相分析方法在沉积学中的作用尤为突出。结合碳酸盐岩颗粒类型、沉积结构和岩石基质等各方面特征, 目前已建立了完善的碳酸盐岩分类体系和对应的沉积相模式(Folk, 1959; Dunham, 1962; 余素玉, 1989; Flü gel, 2010)。碳酸盐岩微相分析方法在地质历史时期高分辨率地层沉积学方面具有明显的优势(Jin et al., 2000; 曹长群和郑全峰, 2009), 结合高分辨率地质年代学可更好地诠释重大地质突变期前后古环境和古生态的精细变化特征。
东秦岭陕西镇安地区二叠系— 三叠系的地层出露连续、厚度巨大, 几乎全为碳酸盐岩沉积序列。对于该地区P-T界线附近的地层已做过大量基础性研究, 如白斌等(2002)将该区二叠系— 三叠系划分为4个层序单位, 认为4次大幅度海平面变化及区域构造沉降作用是形成4个Ⅱ 级不整合面的根本原因; Zhang等(2020)将牙形石Hindeous parvus的首现位置确定为聂家沟剖面二叠系与三叠系之间的界线(位于金鸡岭组最下部鲕粒灰岩底部向上2 m的位置)。文中基于该区域P-T之交高分辨率碳酸盐岩微相的研究, 试图从古生态和古环境角度深入了解二叠纪末生物大灭绝事件前后该区域生物与环境的协同演化关系。
研究剖面位于陕西省镇安县聂家沟附近(图1), 处于秦岭褶皱系南秦岭褶皱带的风县— 金鸡岭褶皱束东段(白斌等, 2002), 北接秦岭造山带。在二叠纪— 三叠纪之交, 南秦岭微板块拼接于华北板块最南缘, 与华南板块隔勉略洋相望(Zhang et al., 2020)。秦岭造山带是在长时间的多期构造运动中形成的, 特征复杂(张国伟等, 1996; 李三忠等, 2002)。中三叠世, 华北、华南和秦岭微板块完成了最终拼合, 结束了中国“ 南海北陆” 的地理格局(裴放和王凌云, 2006; 陈衍景, 2010; Dong et al., 2015; 张国伟等, 2019)。
 | 图1 二叠纪— 三叠纪之交全球古地理图及陕西镇安聂家沟剖面位置 A— 二叠纪— 三叠纪之交全球古地理图(据Ziegler, 1988; 有修改); B— 交通位置; C— 大地构造位置(据张国伟等, 2019; 有修改)Fig.1 Global palaeogeography map of the Permian-Triassic transition and location of Niejiagou section in Zhen'an, Shaanxi |
聂家沟剖面二叠纪— 三叠纪之交发育龙洞川组和金鸡岭组, 两者呈整合接触关系。乐平世海侵旋回基本上呈西浅东深的特点, 西部地层以亮晶颗粒灰岩为主, 东部地层以颗粒泥粒灰岩、颗粒粒泥灰岩及泥晶灰岩为主。乐平统龙洞川组由灰色— 浅灰色厚层亮晶颗粒灰岩组成, 生物化石种类丰富, 包括腕足类、有孔虫类、钙质藻类、棘皮类和珊瑚等。早三叠世海侵事件后, 全区再次变为浅水碳酸盐沉积环境, 金鸡岭组主要发育薄层— 中层泥晶灰岩, 生屑颗粒丰度及分异度极低, 仅见少量双壳类、小型腹足类和小型有孔虫等化石(白斌等, 2002; 程成等, 2017)。
微相分析的主要目的是分析“ 微” 环境, 这就决定了它的研究内容是分析能够反映沉积环境信息的岩石微观特征(包洪平和杨承运, 1999)。微相分析方法由野外工作和室内工作两部分组成, 野外工作主要是对研究区域进行实地考察, 包括岩性描述、地层测量、记录化石类型及采集有价值的化石等, 并根据沉积特征和生物组合对沉积环境进行初步的分析和划分, 而室内工作是利用偏光显微镜对岩石薄片进行观察, 根据岩石颗粒类型和丰度、沉积构造、岩石基质和组构等特征对岩石薄片进行分类和命名, 结合野外宏观特征对微相类型进行系统地划分, 分析古环境和古生态意义。
文中以陕西省镇安县西口回族镇聂家沟剖面二叠纪— 三叠纪之交地层为研究对象(图2)。在实地勘测的基础上, 以逐层采样的方式自下而上系统采集了54块岩石样品。乐平统龙洞川组从上至下采集了3块样品, 未见底; 金鸡岭组从下至上采集了51块样品, 未见顶。采样间距3~64 cm, 平均间距16 cm。岩石样品及岩石薄片均保存在河南理工大学生物遗迹与成矿过程重点实验室。室内分析时, 采用Axio Imager M2研究级数字透反射偏光显微镜对薄片进行观察和照相。碳酸盐岩微相分类参照Dunham(1962)及Flü gel(2010)的分类体系, 并根据实际情况稍做修改。
 | 图2 陕西镇安聂家沟剖面二叠纪— 三叠纪之交的地层宏观特征 A— 二叠系— 三叠系层序, a和d为研究剖面顶、底分界线, b为龙洞川组和金鸡岭组的分界线, c为PTB的界线; B— 金鸡岭组叠层石; C— 金鸡岭组斑状凝块岩(黑色箭头)和泥岩(白色箭头)Fig.2 Outcrop characterisitics of the Permian-Triassic transition strata at Niejiagou section in Zhen'an, Shaanxi |
碳酸盐岩颗粒类型及其沉积特征是划分微相类型、分析沉积环境及海平面变化的重要依据(赵俊兴等, 2003; Flü gel, 2010)。陕西镇安聂家沟剖面乐平统龙洞川组和下三叠统金鸡岭组中常见的颗粒类型有生物碎屑、包壳型颗粒、似球粒、叠层石、囊泡和包粒(图3)。
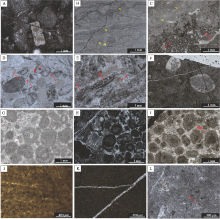
 | 图3 陕西镇安聂家沟剖面下三叠统金鸡岭组下部典型生物颗粒 A— 金鸡岭组, 第41层, 个别层位见少量似球粒颗粒(Pl), 单偏光; B— 金鸡岭组, 第9层, 双壳类生物颗粒(Bi), 单偏光; C— 金鸡岭组, 第26层, 见(f)厄尔兰德虫(Earlandia), 单偏光; D— 金鸡岭组, 第27层, 腕足化石碎屑(Br), 壳体呈层纤状结构, 单偏光; E— 金鸡岭组, 第28层, 见完整腹足类生物化石, 单偏光; F— 金鸡岭组, 第42层, 见个别介形虫颗粒(o), 单偏光; G— 金鸡岭组, 可见有孔虫(f)球旋虫(Glomospira), 第34层, 单偏光; H— 金鸡岭组, 见有孔虫(f)节房虫(Nodosaria), 第34层, 单偏光; I— 金鸡岭 组, 第24层, 可见大小混杂的核形石(On)、鲕粒和包粒, 单偏光Fig.3 Microscopic photos of biological particles in lower part of the Lower Triassic Jinjiling Formation at Niejiagou section in Zhen'an, Shaanxi |
1)生物碎屑。纵向上分布范围广, 分布比例不均衡。龙洞川组生物碎屑丰度及分异度高, 生物碎屑有钙藻、有孔虫、䗴、棘皮类和苔藓虫等(图4-D, 4-E)。金鸡岭组生物碎屑具有种类少、丰度低及体积小等特点, 包括双壳类、海百合茎、有孔虫、腕足类、腹足类及介形虫等(图3-B至3-H)。
 | 图4 陕西镇安聂家沟剖面二叠纪— 三叠纪之交的沉积微相特征 A— MF1, 斑状凝块岩, 第4层, 可见大量形状不规则的囊状微生物化石, 化石内部由块状方解石和微亮晶组成, 单偏光; B— MF2, 叠层石灰岩, 第25层, 叠层石明暗交替, 形态从上至下依次可见缓波状、平行状及穹顶状, 单偏光; C— MF3, 鲕粒— 纹层状叠层石灰岩, 第26层, 右上部为纹层状叠层石(箭头位置), 左下部为鲕粒颗粒, 可见双壳类(b)和海百合茎(e), 单偏光; D— MF4, 有孔虫颗粒灰岩, 第3层, 可见大量䗴和有孔虫(f)化石、海百合茎(e)和钙藻(d), 颗粒间为亮晶方解石填充, 单偏光; E— MF5, 藻— 海百合泥粒灰岩, 第2层, 可见个别有孔虫(f)、粗枝藻(d)、海百合茎(e), 颗粒间主要为亮晶方解石填充, 单偏光; F— MF6, 第38层, 含有被包壳和被磨蚀骨屑颗粒的粒泥灰岩, 可见体腔内被亮晶方解石填充的双壳类; G— MF6, 鲕粒颗粒灰岩, 第1层, 可见大量鲕粒颗粒, 颗粒分选中等, 粒间为亮晶方解石填充, 单偏光; H— MF7, 集合颗粒灰岩, 第9层, 可见大量鲕粒、团粒和集合颗粒, 颗粒主要分布在亮晶方解石或微亮晶内, 单偏光; I— MF8, 鲕粒粒泥灰岩, 第28层, 见变形破碎的鲕粒(Oo), 颗粒间为亮晶方解石胶结, 单偏光; J, K— MF10, 泥岩, 无生物碎屑颗粒, 见大量黑色有机质, 单偏光; L— MF11, 泥晶灰岩, 生物碎屑极少, 见介形虫(o)Fig.4 Sedmentary microfacies of the Permian-Triassic transition strata at Niejiagou section in Zhen'an, Shaanxi |
2)包壳型颗粒。鲕粒分布于金鸡岭组下部和中部, 以圆形或椭圆形的单皮鲕为主, 集中分布, 粒径约在500~1500 μ m之间, 分选中等(图5-G)。纵向上, 最下部地层中鲕粒的圈层结构较多, 核心多为亮晶方解石, 偶见有孔虫和腹足类碎屑。各层位的鲕粒均出现不同程度的重结晶(图4-G)。核形石粒径为毫米级到厘米级, 分布于金鸡岭组中部, 多与鲕粒伴生出现(图3-I)。核形石核心已被完全泥晶化, 内部原生结构无法辨认, 外层见泥晶套。
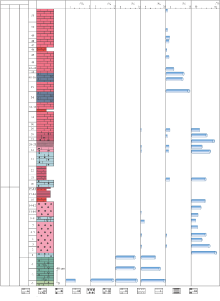
Zhang et al., 2020)'> | 图5 陕西镇安聂家沟剖面二叠纪— 三叠纪之交的综合地层柱状图(生物地层及碳同位素数据引用自Zhang et al., 2020)Fig.5 Stratigraphic column of the Permian-Triassic transition strata at Niejiagou section in Zhen'an, Shaanxi (biostratigraphic strata and carbon isotope data from Zhang et al., 2020) |
3)似球粒。分布于金鸡岭组中部(图3-A)。似球粒表面混浊, 内部均质, 外部呈次圆状、棒状或无明显形状。优势颗粒粒径小于500 μ m。
4)叠层石。分布于金鸡岭组中部, 纹层组构较为发育, 侧向连续性好(图4-B)。垂向上可见缓波状、水平状及穹顶状纹层交替出现, 生物丰度及分异度极低。见极少量有孔虫和藻类颗粒。个别层位见叠层石与鲕粒交替出现。
5)囊泡。分布于金鸡岭组下部斑状凝块石内(图4-A), 由微晶外壁和内部填充亮晶方解石组成, 优势颗粒粒径为500~1000 μ m。囊泡形状不规则, 大部分呈球形、椭球形及不规则球形, 外围轮廓平滑, 无棱角。
6)包粒。分布于金鸡岭组中部, 与鲕粒伴生出现(图3-I)。包粒外部为无纹理泥晶镶边, 核心为亮晶方解石(Flü gel, 2010)。研究区包壳的粒径在200~500 μ m之间, 呈不规则圆状, 分选性较好。
根据沉积结构、颗粒类型及生物组合等特征, 参照Dunham(1962)和Flü gel(2010)的分类体系, 将陕西镇安聂家沟剖面二叠纪— 三叠纪之交地层划分为11种微相类型(图4, 表1)。
| 表1 陕西镇安聂家沟剖面二叠纪— 三叠纪之交的沉积微相类型及特征 Table 1 Types and characteristics of sedmentary microfacies of the Permian-Triassic transition strata at Niejiagou section in Zhen'an, Shaanxi |
乐平统龙洞川组主要发育浅灰色厚层生物碎屑灰岩, 碳酸盐岩微相类型有有孔虫颗粒灰岩和藻— 海百合泥粒灰岩。颗粒类型以生物碎屑为主, 包括有孔虫、䗴、海百合茎及钙藻等。
金鸡岭组岩性以灰褐色中层凝块石灰岩、灰色厚层泥晶灰岩及灰褐色薄层叠层石灰岩为主。微相类型包括鲕粒颗粒灰岩、斑点状凝块石、集合颗粒灰岩、叠层石灰岩、鲕粒— 纹层状叠层石灰岩、含有被包壳和被磨蚀骨屑颗粒的粒泥灰岩、含鲕粒的粒泥灰岩、泥岩和泥晶灰岩。碳酸盐岩颗粒类型有生物碎屑、鲕粒、似球粒、包粒及核形石等, 生物碎屑丰度及分异度极低, 有双壳类、腹足类、腕足类、海百合茎、有孔虫及介形虫等。垂向上, 生物颗粒含量整体呈递增趋势, 下部地层中的生物颗粒以大量的微生物为主, 上部地层中双壳类生物含量有所增加, 但只集中分布于少数层位。
陕西镇安聂家沟剖面二叠纪— 三叠纪之交的碳酸盐岩微相特征反映出频繁的相带变化(图5)。龙洞川组和金鸡岭组自下而上依次发育有台地边缘、局限台地及开阔台地沉积, 垂向序列上3种沉积环境更替出现。金鸡岭组微生物岩有叠层石和凝块石, 两者形成于不同的水深环境, 其中前者主要形成于潮间带— 浅潮下高能带, 而后者形成于深潮下低能带(刘建波等, 2007)。在叠层石灰岩中出现了水平状、缓波状及穹顶状等不同形状的纹层(图4-B), 亦代表了不同水深的沉积环境(Ezaki et al., 2004)。另外, 鲕粒灰岩中形态、大小不同的鲕粒、核形石和包粒等颗粒混杂出现(图3-I), 也体现出沉积环境频繁地波动。
聂家沟剖面乐平统龙洞川组上部(-3--1层)岩性为浅灰色— 灰色厚层亮晶颗粒灰岩, 颗粒磨圆度高, 分选性好, 生物丰度和分异度高, 多为窄盐性海洋生物, 如钙藻、海百合及有孔虫等, 指示了水体环境动荡的台地边缘浅滩环境。金鸡岭组(1-51层)沉积微相特征显示海平面经历了多期波动, 整体上指示1个海侵序列(图5): 金鸡岭组下部地层(1-15层)以斑状凝块石灰岩为主, 生物丰度高但分异度低, 见大量钙质微生物和极少量的双壳类和海百合茎生屑, 指示水体环境稳定的开阔台地沉积环境; 金鸡岭组中部地层(15-28层)整体呈退积— 进积— 退积的层序特征, 纵向上依次交替, 发育局限台地环境的泥岩、叠层石灰岩和鲕粒— 叠层石灰岩与浅滩环境的鲕粒颗粒灰岩以及开阔台地环境的鲕粒粒泥灰岩; 金鸡岭组上部地层(29-51层)沉积时相对海平面变化整体呈进积序列特征, 伴多期海平面小幅度波动, 岩石类型以泥晶灰岩为主, 中间夹泥岩和含有被包壳和被磨蚀骨屑颗粒的粒泥灰岩, 生物含量较低, 有双壳类、介形虫和小型有孔虫等, 指示水体环境稳定的开阔台地环境。
综合聂家沟剖面二叠纪— 三叠纪之交后生生物碎屑(双壳类、腕足类、有孔虫和海百合等)的分布特征(图6), 表明二叠纪末生物大灭绝事件前后浅海碳酸盐岩台地古生态特征存在着明显的差异。
 | 图6 陕西镇安聂家沟剖面二叠纪— 三叠纪之交的生物碎屑及微生物岩分布示意图Fig.6 Distribution of bioclastic and microbialites of the Permian-Triassic transition strata at Niejiagou section in Zhen'an, Shaanxi |
在二叠纪末生物大灭绝事件之前, 乐平统龙洞川组的主要岩性为亮晶颗粒灰岩, 富产结构复杂的䗴、有孔虫、棘皮类和钙藻等生物化石。依据统计数据结果(图6)可知, 后生生物分异度和丰度在鲕粒颗粒灰岩处(金鸡岭组第1层)大幅度降低, 但䗴和有孔虫生物灭绝时间要早于棘皮类和钙藻。具体表现为: 棘皮类和钙藻的含量随时间变化的趋势平缓, 且钙藻表现尤为明显; 䗴和有孔虫含量在藻— 海百合泥粒灰岩处(龙洞川组第-2层)急剧下降, 镜下薄片中仅见极个别完整的䗴和有孔虫化石(图4-E)。后生生物的演化特征表明, 䗴和有孔虫对于环境变化的敏感度和响应速度快于海百合茎和钙藻, 更容易受到灾变事件的影响(Song et al., 2015), 也从侧面进一步证实了受生理特征影响, 生物对于大灭绝事件具有选择性和不等时性的特点(Shen et al., 2009; Song et al., 2013; Hull, 2015)。
二叠纪末生物大灭绝事件后, 浅海碳酸盐岩台地生态系统遭到严重打击, 䗴和钙藻从生态系统中彻底消失, 有孔虫、棘皮类、腕足类及双壳类等也遭受不同程度的重创。因此, 下三叠统金鸡岭组后生生物的丰度及分异度极低, 生物群落以微生物为主, 含有极少量的小型有孔虫、双壳类、介形虫和海百合茎等(图6)。
综上所述, 受二叠纪末生物大灭绝事件的影响, 乐平世生物丰富的浅海碳酸盐岩台地生态环境被早三叠世以微生物为主并伴有小型腹足类、有孔虫及双壳类等广栖生物的简单生态系统所代替。这一生态演替特点与中国华南同时期其他浅海碳酸盐岩台地一致(Song et al., 2013; Tian et al., 2019)。
下三叠统金鸡岭组中产出的生物多为双壳类、海百合、腹足类以及有孔虫, 腕足类产出较少, 主要原因在于双壳类、海百合、腹足类及有孔虫等对环境波动(如盐度、氧气含量、浑浊度及湍流等)的适应能力强(Clapham and Bottjer, 2007; Song et al., 2013; Chen et al., 2019), 而对海洋环境变化敏感、生物缓冲差的腕足类成为二叠纪末大灭绝事件中最大的受害者之一(Clapham and Bottjer, 2007)。同时, 金鸡岭组中的有孔虫、腕足类及双壳类等生物出现了小型化效应(“ Lilliput effect” )(Twitchett, 2007; Bing et al., 2010), 其中有孔虫以管状和盘状为主, 个体远小于灭绝前的水平(图3-C, 3-G, 3-H), 腕足类和双壳类生物也出现形态变小、壳体厚度变小的变化趋势。这种小型化现象, 目前主要归因于缺氧、海洋酸化及全球变暖等恶劣的环境条件(Kaiho, 1998; Hunt and Roy, 2006; Kuroyanagi et al., 2009; Song et al., 2011; Kershaw et al., 2018)或是降低同类竞争水平(Chen et al., 2019)。
1)系统的碳酸盐岩微相分析表明, 陕西镇安聂家沟剖面二叠纪— 三叠纪之交的沉积可划分为11种碳酸盐岩微相类型, 其中乐平统龙洞川组顶部为有孔虫颗粒灰岩和藻— 海百合泥粒灰岩微相, 金鸡岭组下部为鲕粒颗粒灰岩、斑点状凝块石、集合颗粒灰岩、叠层石灰岩、鲕粒— 纹层状叠层石灰岩、含有被包壳和被磨蚀骨屑颗粒的粒泥灰岩、含鲕粒的粒泥灰岩、泥岩和泥晶灰岩微相。根据碳酸盐岩微相特征及沉积相标志, 将该段沉积划归为3种沉积相类型, 分别是台地边缘相、开阔台地相和局限台地相, 指示聂家沟剖面在二叠纪— 三叠纪之交发生了多期次的沉积相带转化。
2)陕西镇安聂家沟剖面二叠纪— 三叠纪之交的碳酸盐岩微相所反映的古生态学特征显示, 二叠纪末的生物灭绝事件沉重打击了浅海碳酸盐岩台地生态系统, 早三叠世初期变为以微生物碳酸盐岩为主的简单生态系统。
(责任编辑 张西娟; 英文审校 刘贺娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|