



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
滦河三角洲潮坪环境现代生物遗迹组合及其分布特征*
[王海邻1  , 张彬
, 张彬1 , 宋慧波1, 2 , 王长征1 , 胡斌1, 2 , 张立军1, 2 ]
, 张彬, 王长征|
|


第一作者简介 王海邻,女,1971年生,博士,副教授,主要从事环境科学、沉积学和遗迹学的教学和研究工作。E-mail: whl@hpu.edu.cn。
现代生物遗迹的特点、分布和组合特征及其与沉积环境、沉积物性质的关系,对遗迹化石及古环境研究具有重要的实证价值,但现代生物遗迹研究起步较晚,国内研究成果较少。文中论述了滦河三角洲潮坪环境中现代生物遗迹的组成、组合与分布特征,并分析了遗迹组合与沉积环境的关系。研究表明: (1)层面遗迹以节肢动物、环节动物、腹足动物的爬行迹和拖迹为主; 层内遗迹包括垂直或高角度倾斜于层面的潜穴、平行于层面的潜穴,以居住迹为主,丰度高,类型多样。(2)建立了3种生物遗迹组合: 遗迹组合I以Y形和垂直管状潜穴为特征,局部丰度较高,表层潜穴和层面拖迹不发育,造迹生物主要为沙蚕和螃蟹,沉积物底层具有砂含量较高、水动力强、有机质含量低的特点,显示其出现于三角洲潮坪砂质沉积环境;遗迹组合Ⅱ以各种形态的层内潜穴和大量的层面爬行拖迹为特征,层内潜穴包括垂直管状、J形、U形、Y形和分枝形潜穴,丰度高,造迹生物主要是沙蚕和螃蟹,层面遗迹常见爬行拖迹和足辙迹,造迹生物主要为沙蚕、螃蟹、扁玉螺、寄居蟹,沉积物底层具有高泥质含量、高含水率和高有机质含量的特点,形成于三角洲潮坪中低潮区;遗迹组合Ⅲ以与层面近似垂直和与层面近似平行的层内潜穴以及大量的层面运动迹为特征,与层面近似垂直的潜穴包括垂直管状、J形、U形、Y形和复杂分枝形潜穴,造迹生物主要是螃蟹、泥甲虫及其幼虫, 与层面平行的表层潜穴主要是蝼蛄、泥甲虫及其幼虫营造的,层面上的运动迹常见为螃蟹足辙迹和鸟类足迹,沉积物底层具泥质含量高、含水率低、有机质含量高、底质固结程度较高的特点,产生于三角洲潮坪高潮区。(3)基于对遗迹组合和分布特征及其与底层性质和沉积环境关系的分析,建立了三角洲潮坪生物遗迹组合的沉积环境分布模式。该研究成果既可作为古代三角洲潮坪沉积环境的判识依据,也可为相似遗迹化石组合的沉积环境恢复提供佐证。
About the first author WANG Hailin,born in 1971,Ph.D.,is an associate professor of Henan Polytechnic University. She is mainly engaged in teaching and researches of environmental sciences,ichnology and sedimentology. E-mail: whl@hpu.edu.cn.
The characteristics,distribution and assemblage of lebensspuren and their relationships with sedimentary environment and sediment properties are significant for the study of trace fossils and palaeoenvironment. However,the research on lebensspuren started late and few related work is found in China. The paper discussed the composition,assemblage and distribution characteristics of lebensspuren in tidal flat of Luanhe delta,and analyzed the relationship between the biological assemblage and the sedimentary environments. The results are as follows: (1)The lebensspuren at the surface of bedding mainly include the repichnia and trails made by arthropoda,annelida and gastropods,and the lebensspuren in the bedding include the burrows approximately perpendicular and parallel to bedding plane,in which the domichnia is dominant with high abundance and various types. (2)Three kinds of biological assemblages are established. Type I is characterized by Y-shaped and vertical tubular burrows with locally high abundance. The epigenic burrows and trails on the bedding plane are less,and the trace makers mainly include Nereis succinea and Brachyura. The bottom of the sediments are characterized as high sand content,high intensity hydrodynamics and low organic matter content,indicating the sandy delta tidal flat sedimentary environment. Type II is characterized as various burrows in the substrate and numerous trails on the surface of bedding. The burrows in the substrate include vertical tubular,J-shaped,U-shaped,Y-shaped and branched burrows with high abundance,and the trace makers mainly include Nereis succinea and Brachyura. The lebensspuren on the formation surface mainly include repichnia and trackways,e.g., Nereis succinea, Brachyura, Glossaulax didyma and Paguridae. The substrate sediments are characterized as high muddy content,high water content and high organic matter content,forming in the middle and lower tidal area of delta tidal flat. Type Ⅲ is characterized by the burrows approximately perpendicular and parallel to the bedding plane and motion traces on the bedding surface. The burrows in the substrate include vertical tubular,J-shaped,U-shaped,Y-shaped and complex branched burrows,which are mainly made by Brachyura,mud beetles and their larvaes. The burrows parallel to bedding plane are mainly made by Cryllotalpa,mud beetles and their larvaes. The motion traces on the bedding surface include the trackways made by Brachyura and footprint made by birds. The substrate sediments for type Ⅲ assemblage showed the characteristics of high muddy content,low water content,high organic matter content and high degree of consolidation,indicating the high tidal area of delta tidal flat. (3)Based on the analyses of lebensspuren assemblage,distribution characteristics and the relationship with the substrate sediments properties and sedimentary environment,a sedimentary environment distribution model of lebensspuren in deltaic tidal flat was established. The research results can not only acted as a basis for the identification of the sedimentary environment of the ancient delta tidal flat,but also provide evidence for the sedimentary environment restoration of similar relic fossil assemblages.
遗迹学是研究古代和现代生物在沉积物层面和层内活动时遗留下来的痕迹(吴贤涛, 1986; 胡斌等, 1997), 与生态条件、沉积物性质、沉积环境有着密切的关系。研究现代生物遗迹的特点、分布和组合特征及其与沉积环境、沉积物性质的关系, 对遗迹化石及古环境研究具有重要的实证价值。
关于现代生物遗迹的研究, 尤其是滨岸潮间带现代生物遗迹研究, 受到了国外遗迹学者的广泛关注(Seilacher, 1954, 1964, 1967; Frey, 1970; Frey and Howard, 1986, Frey et al., 1987; Frey and Basan, 1987; Seike et al., 1999, 2011; Jorge et al., 2009), 如丹麦的Richard和Bromley(1996)对潮间带造迹生物的生活习性和造迹过程开展了详细的调查和研究, 波兰的Uchman和Pervesler(2006)针对现代潮间带沉积物的性质与生物遗迹形态特征的关系开展了较详细的研究, 他们的研究成果极大地推进了遗迹学的发展。20世纪 90年代以来, 现代三角洲的新遗迹学研究开始受到关注, 对河控、浪控和混合控制三角洲的遗迹学特征开展了相关研究(Gingras et al., 1998; Gani et al., 2004)。进入21世纪以来, 学者们加深了对新遗迹形态学的描述、沉积环境的分析及其与遗迹化石的对比(Abdel-Fattah, 2019; Paz et al., 2020), 如Paz等(2020)通过描述不同物理化学因素下浪控三角洲和河控三角洲的相对优势, 发现河控三角洲中造迹生物受到的环境压力较大, 浪控的相对缓和。
国内对古遗迹学研究较多, 对现代生物遗迹的研究起步较晚, 开展工作相对较少。在潮坪沉积方面, 王珍如等(1988, 1994)对北部湾和青岛日照潮坪沉积中的现代生物遗迹开展了研究, 王英国(2000)研究了渤海湾西岸大石河河口湾的现代生物遗迹, 宋慧波等(2014)和王海邻等(2017a, 2017b)对杭州湾庵东浅滩以及青岛日照沿岸潮间带的现代生物遗迹进行了研究。关于现代三角洲的新遗迹学研究开展的更少, 仅对黄河三角洲潮坪沉积中的现代生物遗迹的组成和分布特征以及现代生物遗迹分布与物化条件的响应关系进行了研究(胡斌等, 2012, 2015; Wang and Hu, 2014; Wang et al., 2019a, 2019b; 王媛媛等, 2019)。
关于滦河三角洲的沉积学研究已开展的较为广泛和深入, 但相关的新遗迹学研究则较少进行, 目前公开报道的仅为王观忠和吴贤涛(1992)、胡斌等(2015)对滦河三角洲的现代生物遗迹进行了调查和描述。本次工作是在野外调查的基础上, 通过取样分析, 针对滦河三角洲前缘现代沉积物中的生物遗迹组成、组合和分布特征及其与沉积环境和沉积物底层性质之间的关系进行了较深入的分析, 建立了三角洲潮坪不同沉积亚环境的生物遗迹组合。该研究成果一方面能丰富中国新遗迹学的研究内容, 另一方面可为地史时期三角洲沉积环境的研究乃至古地理、古生态、古环境的重建提供佐证。
滦河是中国第二大多沙性河流, 发源于内蒙古高原, 流经燕山山脉, 从河北滦县县城北面出山后进入下游平原, 在乐亭县姜各庄镇汇入渤海, 形成了以滦县为顶点的冲积扇→河流→三角洲沉积体系。历史上滦河经历过多次变迁, 最近一次是1915年在莲花池村附近冲决滨海砂丘注入渤海, 不断淤积形成了北起塔子沟、南至网子沟的最新三角洲(刘福寿, 1993)。滦河三角洲为河控—波控三角洲, 位于中纬度季风区, 气候具有显著的大陆性特征, 潮汐属于正规和不正规的半日潮, 距离滦河口较近的秦皇岛平均潮差66 cm, 最大潮差376 cm(1980—2002)(薛春汀, 2016)。
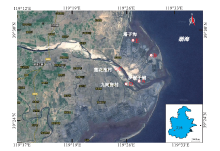
滦河三角洲潮滩现代沉积物中浮游生物和底栖生物较多, 本次野外观察时综合考虑了沉积物底质类型和人工干扰程度, 最终确定了罗锅子铺、塔子沟和莲花池村附近A、B、C 、D、E 共5个观察点(图1)进行分析。
 | 图1 滦河三角洲潮坪现代生物遗迹考察点位置(来自Google Earth(TM)) A、B、C 、D、E-野外观察点Fig.1 Location of investigated sites of ichnoassemblage in tidal flat of Luanhe delta(from Google Earth(TM)) |
滦河三角洲可分为三角洲平原、三角洲前缘和前三角洲。三角洲平原位于高潮线之上, 现代滦河河道分流点大约在莲花池村附近, 呈扇形向东南突出, 包括废弃河道、河床、天然堤、泛滥平原等次一级的地貌单元, 沉积物主要为中—细砂、细砂、粉砂及黏土质粉砂等(李从先等, 1984)。三角洲前缘位于高潮线至波浪作用基底水深(波基面或正常天气浪基面), 在现代滦河入海口处达6~7 m深, 向入海口两侧水深减至4 m左右。滦河三角洲主要受海浪和潮汐作用影响, 发育三角洲潮坪, 包括滨外沙坝、潟湖和水下岸坡3个次一级的地貌单元(高善明, 1983; 李从先等, 1984)。
三角洲潮坪沉积物以细砂和粉砂为主, 其中滨外沙坝和水下岸坡沉积由细砂组成, 潟湖沉积物相对较细, 含一定量的粉砂, 分布在现代河口外及北、南两侧(李从先等, 1984; 黎刚和殷勇, 2010)。
滦河三角洲发育多种砂泥交互沉积。因受河水、海浪、潮汐、风等多种地质营力影响, 发育多种沉积构造, 常见的有波状层理、水平层理、爬升层理和槽状交错层理等。 在潟湖、积水洼地处有大范围的泥裂现象, 潮滩上因遭受潮流和波浪作用, 浪成波痕发育, 常见的有对称和不对称的波痕、干涉波痕、尖顶的或削顶的波痕、舌形波痕等(图2)。
 | 图2 滦河三角洲潮滩的地貌特征和常见沉积构造 A—塔子沟附近E处潮间带; B—九间房附近D处潮坪上的波痕; C—罗锅子铺附近A处的风成沙丘; D—莲花池村附近C处的水平层理Fig.2 Geomorphic characteristics and common sedimentary structures in tidal flat of Luanhe delta |
本研究选取滦河三角洲潮坪的沉积物和现代生物遗迹作为野外考察对象, 对沉积物性质和生物遗迹的形态学、沉积学和遗迹学进行研究。通过手持GPS进行采样点定位, 采集离层面20 cm深的沉积物样品进行粒度和总有机碳(TOC)含量分析, 共采集沉积物样品5个。采集水样5个, 用于测定盐度。所有实验分析均在河南理工大学完成。
粒度分析测试时, 取风干样品0.2 g, 加 10 mL浓度30%的H2O2煮沸至反应平静, 冷却; 再加 10 mL浓度10%的HCl 煮沸至反应平静; 冷却后加蒸馏水, 沉淀 12 h 至溶液清亮; 倒掉清液后, 加入 5 mL 浓度为36 g/L 的NaPO3进行搅拌, 放入震荡仪使颗粒高度分散。 待完全沉淀后, 使用 Mastersize2000进行测量, 测量误差小于1%。
TOC样品制备方法如下: 在105 ℃下对样品进行24 h初始干燥; 取研磨土样2 g, 加入30 mL浓度36%的盐酸, 放入封闭的样品管, 标记液体高度; 离心机离心5 min, 静置后倒掉上部清液, 加纯水至标记高度; 放入离心机去酸化, 重复4~5遍, 至pH值为7; 处理后的土样置于烧杯中放入烘箱, 60 ℃烘干3 d, 每天10 h。样品处理完成后, 各样品称取2份5 mg, 土壤标准物质称取3 mg, 用锡杯包裹, 使用仪器Apollo 9000通过燃烧法测定沉积物中有机碳的含量。在燃烧损失分析期间, 用精确到0.001 g的电子秤测定有机物燃烧造成的重量损失。
海水盐度采用手持式盐度计 SX813测试, 分辨率 0.01。
现代生物遗迹组合与分布特征主要受控于造迹生物的组成与分布, 并受到气候和沉积环境的影响(Ekdale et al., 1984)。潮坪上的主要造迹生物为水生底栖生物, 它们生活的沉积物底层与水深、盐度、含氧量、水体能量、底层性质(粒度、固结程度等)、有机质含量和食物的供给等环境因素密切相关, 这些因素相互联系, 影响着生物群落的分布。造迹生物的种类、分异度、个体大小、内生生物的活动量和行为方式等, 直接影响着生物遗迹的形态、产状、组合和分布特征等。本次工作针对滦河三角洲潮坪不同沉积亚环境下沉积物的粒度、有机质含量、海水盐度进行了取样和测试, 对不同沉积亚环境的水动力条件、沉积底层性质进行分析, 同时观察和研究现代生物遗迹的组成、组合与分布特征以及造迹生物的种类和造迹行为方式。
研究区沉积物粒度特征见表1, 海水盐度和沉积物TOC含量、固结程度、含氧量及水动力条件见表2。由表1和表2可知, 沉积物粒度组成与沉积环境有关, 在距离潮沟、河道和河口较近处, 入海泥沙堆积。在波浪、潮汐、风力的共同作用下, 位于河口附近的A、C和D处潮滩水动力条件相对较弱, 沉积物中泥和细沙含量相对较高, 有机质含量也相对较高; 而B和E处的潮滩海浪作用较强, 沉积物中泥质含量很少、粗沙含量高、有机质含量低。
| 表1 滦河三角洲潮坪沉积物粒度 Table 1 Sediment grain size in tidal flat of Luanhe delta |
| 表2 滦河三角洲潮坪沉积物性质 Table 2 Sediment properties in tidal flat of Luanhe delta |
研究区沉积物中观察到的造迹生物常见的共有7种, 包括节肢动物门的螃蟹(Brachyura)和寄居蟹(Paguridae)、软体动物门的扁玉螺(Glossaulax didyma)、环节动物门的沙蚕(Nereis succinea)、直翅目的蝼蛄(Gryllotalpa)、鞘翅目的泥甲虫(Coleoptera)及其幼虫、鸟类等。遗迹的生态类型主要是爬行迹(拖迹、足辙迹)、居住迹、停息迹、觅食迹、鸟类足迹以及生物粪粒等(表3)。
| 表3 滦河三角洲潮坪造迹生物种类及其遗迹生态类型 Table 3 Trace-makers category and their ethological types in tidal flat of Luanhe delta |
3.3.1 塔子沟附近地区 观察点B和E处的潮滩, 位于滦河三角洲最北部。该区潮滩地貌呈弧形, 面积较小, 潮间带水动力条件较强, 沉积物较疏松。表层沉积物呈黄褐色, 颗粒较粗, 有机质含量少, 而自表层向下10 cm左右为黑色沉积物, 有机质含量较高。沉积物以细砂和中砂为主, 含少量泥, 细砂含量为41.64%~41.87%, 中砂含量为36.18%~49.27%, 粗砂含量为2.48%~14.61%, 指示海浪作用强, 受海水影响较大。海水盐度为32.06‰ , 属于潮间带咸水环境。该区观察到的造迹生物主要是沙蚕(Nereis succinea)和螃蟹(Brachyura)。
沙蚕(Nereis succinea)主要分布在潮滩上靠近海水边缘处。塔子沟附近B和E处潮间带水动力较强, 沉积物颗粒较粗, 观察到的沙蚕遗迹主要是层内居住潜穴, 多为垂直管状, 少量为Y形。潜穴直径2~5 mm, 穴道上下等粗, 深度为8~20 cm, 内表面光滑, 整体与层面基本垂直或高角度相交。主要分布在局部泥质含量较多、有机质含量较高的地方, 潜穴丰度低, 在1 m2范围内最多可观察到8~10个潜穴(图3-D)。该区沙蚕营造的层面遗迹很少, 沉积物表面未发现潜穴口、沙蚕的爬行迹和觅食迹, 也未发现沙蚕排泄的长条形粪粒, 这可能与该区域沉积物为砂质、受较强水动力的海浪和潮水冲蚀有关, 这造成沉积物表面的沙蚕潜穴口被砂粒掩盖, 层面的爬行迹、觅食迹和粪粒等也因被冲刷而消失。
 | 图3 滦河三角洲塔子沟附近潮滩砂质沉积中的现代生物遗迹 A—螃蟹营造的垂直管状潜穴; B—螃蟹潜穴开口, 周围散布砂球; C—螃蟹营造的层面潜穴开口, 周围有爪痕和掘穴砂球; D—沙蚕造的Y形和垂直管状潜穴Fig.3 Lebensspuren in the sand sediment of tidal flat near Tazigou area, Luanhe delta |
螃蟹(Brachyura)主要是圆球鼓窗蟹, 一般栖息于潮间带的高潮区, 个体较小。退潮后, 圆球鼓窗蟹在沉积物中向下挖潜, 并将掘穴过程产生的砂粒送出潜穴, 于穴口周围散布。其营造的层内潜穴一般与层面呈高角度斜交, 深15~50 cm, 直径一般1~3 cm, 穴道多不分枝, 形状多为垂直管状或略微倾斜的J型, 穴口一般与沉积物表面齐平, 穴口周围常见掘穴过程产生的砂球和搬运砂球时留下的环绕穴口分布的层面足辙迹(图3-A, 3-B, 3-C)。潜穴周围沉积物层面上还可发现螃蟹爬行时留下的足辙迹, 呈两排平行的点线形分布, 与遗迹化石Diplichnites(双趾迹)类似。
3.3.2 罗锅子铺附近潮间带 距离滦河较近的A处潮滩沉积中, 沉积物以细砂为主, 含少量泥和粉砂, 其中泥含量约为4.78%, 粉砂含量约为21.1%, 细砂含量约为60.16%, 中砂含量12.52%, 粗砂含量1.41%, TOC含量为2.72%, 盐度为31.03‰ 。受滦河河水和海水共同影响, 以海水作用为主, 盐度略低于海水标准盐度, 仍属潮间带沉积环境。该区的造迹生物主要有沙蚕(Nereis succinea)、螃蟹(Brachyura)、寄居蟹(Paguridae)和鸟类。
沙蚕(Nereis succinea)营造的遗迹包括层面遗迹和层内潜穴。层面遗迹包括爬行迹、觅食迹以及沙蚕的粪粒: 爬行迹是沙蚕在沉积物表面爬行过程中产生的, 呈任意弯曲或微弯曲的条带状, 不分枝, 中间呈浅沟状, 两侧为隆起的脊痕, 常与2个潜穴口连接(图4-G), 与遗迹化石Aulichnites(犁沟迹)类似; 沙蚕觅食时反复从潜穴口伸出, 在穴口周围形成几条5~6 cm长、呈星射状分布的表层觅食迹(图4-H); 有时在穴口还可发现呈圆锥形堆积的沙蚕粪粒, 长条形, 长0.2~0.3 mm, 粪粒大小与沙蚕大小有关。层内遗迹主要是居住潜穴, 形状多为小型的垂直管状、U形、Y形, 潜穴上下等粗, 内表面光滑, 穴壁有生物分泌的粘液, 与周围沉积物颜色不同, 直径一般为2~5 mm, 深度8~20 cm(图4-F)。垂直管状潜穴与层面垂直或高角度相交, 形态特征与遗迹化石Skolithos(石针迹)类似; U形潜穴与层面基本垂直, 无蹼状构造, 形态特征与遗迹化石Arenicolites(沙蠋迹)类似; Y形潜穴形态特征与遗迹化石Polykladichnus(多枝迹)类似。该区沙蚕营造的层内潜穴丰度极高, 主要分布在潮滩的中低潮区或者潮间带低洼积水处边缘和潮沟两侧反复被潮水淹没的沉积物中, 在100 cm2范围内可观察到80~106个潜穴, 对0~20 cm深沉积物扰动剧烈(图4-D, 4-E, 4-F)。
 | 图4 滦河三角洲罗锅子铺附近潮滩砂泥互层沉积中的现代生物遗迹 A—石膏浇铸的螃蟹U形潜穴; B—石膏浇铸的螃蟹Y形和J形潜穴; C—石膏浇铸的螃蟹复杂分枝形潜穴; D—沙蚕营造的垂直管状潜穴(斜切剖面特征), 丰度高; E—沙蚕营造的垂直管状潜穴(层面特征), 穴口周围沉积物呈黄褐色晕圈, 部分潜穴已被填充; F—沙蚕营造的Y形、U形、垂直管状潜穴(横剖面特征); G—沙蚕营造的层面爬行迹/拖迹; H—沙蚕营造的层面觅食迹; I—寄居蟹的拖迹和足辙迹Fig.4 Lebensspuren in the interbedded deposits of sand and mud within tidal flat near Luoguozipu site, Luanhe delta |
螃蟹(Brachyura)主要为大眼蟹, 其营造的居住潜穴形态多样, 主要呈大型不规则垂直管状、U形、J形、Y形和复杂的分枝形(图4-A, 4-B, 4-C)。大部分以高角度倾斜于层面向下延伸, 深度15~50 cm, 直径大小不一, 与造迹生物形体大小有关。潜穴口近似长圆形, 穴口常有高出沉积物表层1~2 cm的凸起。U形潜穴与遗迹化石Arenicolites(沙蠋迹)的形状特征类似, J形、Y形和复杂的分枝形潜穴的形态特征与遗迹化石Psilonichnus(裸迹)类似。该区螃蟹营造的层面遗迹主要是爬行过程中产生的足辙迹, 与遗迹化石Diplichnites(双趾迹)类似。
寄居蟹(Paguridae)营造的遗迹主要是在层面爬行过程中产生的足辙迹和拖迹(图4-I), 与遗迹化石Bipodomorpha(双足趾迹)和Gordia(线形迹)类似。寄居蟹的爬行迹丰度极低, 且不易保存。
3.3.3 莲花池村附近潮间带 观察点C和D处沉积物以粉砂和细砂为主, 含少量泥, 其中粉砂含量为31.66%~53.72%, 细砂含量为27.53%~51.74%, 泥含量为9.81%~12.78%, 中砂含量为5.55%~6.7%, 粗砂含量约0.42%, TOC含量为2.46%~2.52%, 盐度偏低, 为27.92‰ ~28.13‰ 。受河水和海水共同作用, 但主要以海水作用为主, 仍属潮间带沉积环境。造迹生物主要为沙蚕(Nereis succinea)、螃蟹(Brachyura)、扁玉螺(Glossaulax didyma)、蝼蛄(Gryllotalpa)、泥甲虫(Coleoptera)及其幼虫、鸟类。层面遗迹主要是沙蚕的爬行迹和星射状觅食迹、螃蟹的足辙迹和扁玉螺的拖迹。层内遗迹包括沙蚕营造的层内潜穴、螃蟹营造的层内潜穴、蝼蛄和泥甲虫及其幼虫营造的与层面平行的居住和觅食潜穴。
沙蚕和螃蟹营造的遗迹形状特征与罗锅子铺附近潮间带相同。沙蚕遗迹主要分布于低洼积水处边缘或潮沟两侧, 离水较近、反复被水淹没, 沉积物中泥质含量高、含水率高、藻类和有机质含量高, 沉积物较软, 利于生物在层内的掘穴和觅食。螃蟹营造的遗迹在高、中、低潮区均有分布。由于沉积物细颗粒含量高, 具有一定的粘结性, 该处生物的层内潜穴易于保存。
扁玉螺(Glossaulax didyma)一般在中、低潮区分布, 在局部低洼积水处高度密集。遗迹主要是层面上的拖迹, 宽3~5 mm, 长约数厘米, 呈曲沟状、任意弯曲形, 两侧有对称或不对称的隆起, 表面光滑, 不分枝, 有时有交切(图5-G)。有时在拖迹的末端可见扁玉螺的停息迹。拖迹形状特征与遗迹化石Gordia(线形迹)类似。
 | 图5 滦河三角洲莲花池村附近潮滩砂泥互层沉积中的现代生物遗迹 A—蝼蛄营造的层面潜穴, 有交切; B—蝼蛄营造的层面潜穴, 上表面有裂纹, 底层含水率较低; C—蝼蛄营造的高角度分枝的层面潜穴, 底层含水率较高; D—泥甲虫及其幼虫营造的层面潜穴, 底层含水率较高; E—泥甲虫及其幼虫营造的复杂网状层面潜穴; F—泥甲虫及其幼虫营造的近平行于层面的潜穴及高角度倾斜于层面的潜穴; G—扁玉螺营造的层面拖迹和停息迹Fig.5 Lebensspuren in the interbedded deposits of sand and mud within tidal flat near Lianhuachi village, Luanhe delta |
泥甲虫(Coleoptera)及其幼虫的遗迹主要分布在高潮区, 其营造的遗迹包括与层面平行的表层潜穴和高角度倾斜于沉积物层面的垂直管状潜穴。平行于层面的表层潜穴直径2~3 mm, 潜穴表面光滑或凸起呈细瘤状或有纵向锯齿状裂纹, 沿表层不规则弯曲, 未观察到潜穴内有衬壁。这些潜穴有的呈高角度分枝, 有的不分枝、直或微弯曲状(图5-D), 有的为复杂网状结构(图5-E), 形状特征与遗迹化石Multina 类似(Buatois, 2017)。泥甲虫营造的与层面近似垂直的管状潜穴, 深1~10 cm, 内壁光滑, 有时潜穴下部膨大(图5-F), 与遗迹化石Skolithos(石针迹)类似。
蝼蛄(Gryllotalpa)营造的遗迹主要是平行于沉积物表面的表层觅食潜穴, 主要分布在潮间带上部至潮上带, 尤其是植物根系发达的地方。潜穴大小与蝼蛄个体大小有关, 直径2~6 cm, 长25~85 cm, 甚至更长, 穴内无填充物, 潜穴内表面有蝼蛄爬行过程产生的“人”字形抓痕, 但未观察到潜穴衬壁。潜穴形状有时为直形或微弯曲形, 有时有分枝, 分枝近直角, 有时呈巨大网状(图5-A, 5-B, 5-C)。潜穴顶部沉积物隆起, 沉积物特性对蝼蛄潜穴形态有影响, 在颗粒较细、含水率较高的湿软沉积物中, 层面上隆起的潜穴表面无裂纹或裂纹较小, 而含水率较低的沉积物中, 隆起的潜穴表面裂纹较大, 顶层有时呈碎薄片状, 有时有纵向贯通的裂纹。蝼蛄营造的遗迹形状特征与遗迹化石Multina 类似, 通常比泥甲虫及其幼虫营造的层面潜穴的直径更粗大。
此外, 该区沉积物表面还有大量的鸟类足迹与波痕共生。
基于对生物遗迹组成和分布特征及其与底层沉积物性质和沉积环境关系的分析, 将滦河三角洲潮坪现代遗迹划分为3种类型的遗迹组合。
3.4.1 遗迹组合I——滦河三角洲潮坪砂质沉积中的遗迹组合 塔子沟附近B和E处潮滩位于滦河三角洲前缘的最北部, 水动力条件强, 海浪冲刷作用强, 受海水影响大, 属于潮间带咸水环境。沉积物呈黄褐色, 较疏松, 颗粒较粗, 以细砂和中砂为主, 仅含少量泥质, 有机质含量低。该环境下的生物遗迹组成是以底层中Y形和垂直管状潜穴为特征(图6), 仅分布在局部有机质含量相对较高的沉积物中, 潜穴形状特征与遗迹化石Skolithos(石针迹)、Polykladichnus(多枝迹)、Psilonichnus(裸迹)类似。表层潜穴和层面拖迹不发育, 有时层面会有与遗迹化石Diplichnites(双趾迹)类似的爬行迹以及螃蟹潜穴开口, 造迹生物主要为沙蚕和螃蟹。该遗迹组合指示了三角洲滨岸潮间带水动力条件较强的砂质潮滩沉积环境。
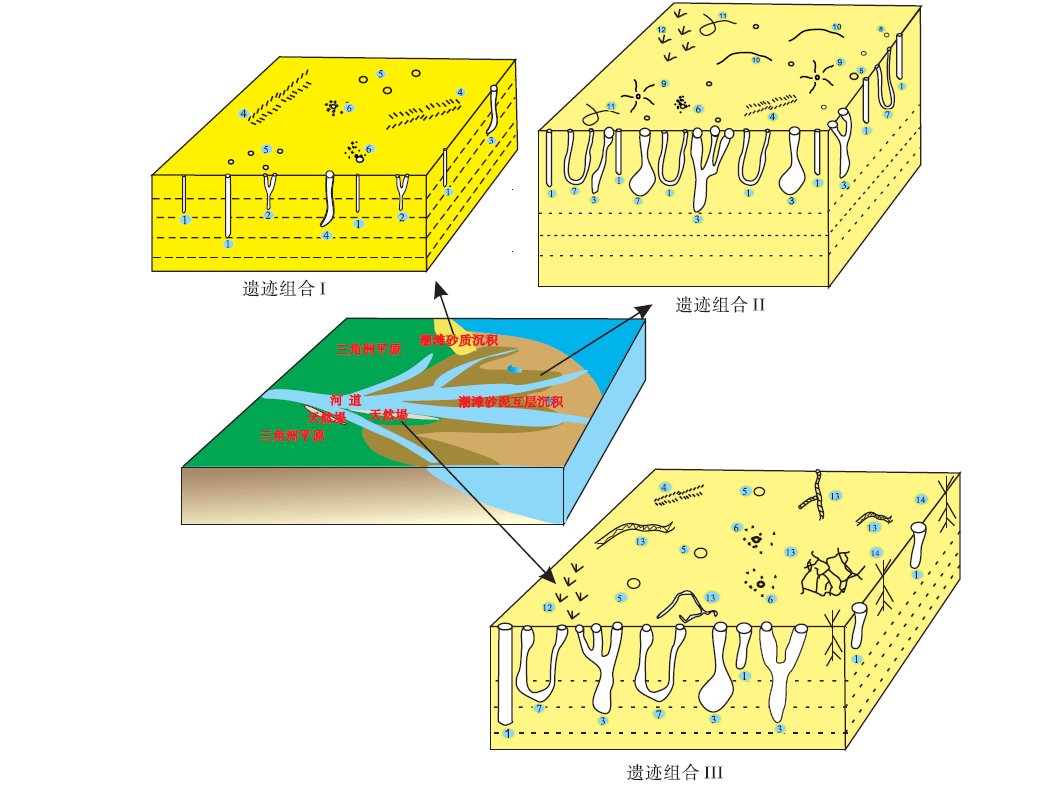 | 图6 滦河三角洲滨岸潮间带生物遗迹组合的沉积环境模式 1—似Skolithos(沙蚕、螃蟹、泥甲虫及其幼虫垂直管状潜穴); 2—似Polykladichnus(沙蚕Y型潜穴); 3—似Psilonichnus(螃蟹J形、Y形及复杂分枝形潜穴); 4—似Diplichnites(螃蟹足辙迹); 5—周围无砂球的螃蟹潜穴开口; 6—周围有砂球的螃蟹潜穴开口; 7—似Arenicolites(沙蚕和螃蟹U形潜穴); 8—沙蚕潜穴开口; 9—沙蚕觅食迹; 10—似Aulichnites(沙蚕层面爬行迹/拖迹); 11—似Gordia(扁玉螺和寄居蟹层面爬行迹/拖迹); 12—鸟足迹; 13—似Multina(蝼蛄、泥甲虫及其幼虫平行层面潜穴); 14—植物根迹Fig.6 Sedimentary environment model of ichnoassemblages in tidal flat of Luanhe delta |
3.4.2 遗迹组合Ⅱ——滦河三角洲潮坪中低潮区砂泥互层沉积中的遗迹组合 罗锅子铺和莲花池村附近A、C和D处潮间带沉积物以细砂和中砂为主, 含少量泥, 砂泥交互沉积, 富含有机质, 潮滩低洼不平, 暂时性水渠和积水洼地较多, 盐度略低于海水标准盐度, 受河水和海水共同影响。该区域中低潮区的生物遗迹组成是以层内各种形态潜穴和大量的层面拖迹为特征(图6):层内潜穴包括与遗迹化石Skolithos(石针迹)类似的垂直管状潜穴、与Arenicolites(沙蠋迹)类似的U形潜穴、与Polykladichnus(多枝迹)类似的Y形潜穴, 以及与Psilonichnus(裸迹)类似的J形、Y形和复杂分枝形潜穴, 潜穴丰度极高, 造迹生物主要是沙蚕和螃蟹。层面拖迹包括4种类型: (1)与遗迹化石Bipodomorpha(双足趾迹)和Gordia(线形迹)类似的寄居蟹爬行迹; (2)与遗迹化石Gordia(线形迹)类似的扁玉螺拖迹; (3)与遗迹化石Aulichnites(犁沟迹)类似的沙蚕拖迹; (4)与遗迹化石Diplichnites(双趾迹)类似的螃蟹足辙迹。此外, 层面上还分布有螃蟹潜穴口、沙蚕潜穴口、沙蚕粪粒和鸟类足迹等。该遗迹组合指示了三角洲潮坪中低潮区的沉积环境。
3.4.3 遗迹组合Ⅲ ——滦河三角洲潮坪高潮区砂泥互层沉积中的遗迹组合 滦河三角洲潮坪高潮区发育砂泥互层沉积, 但相对来说泥多砂少, 生物遗迹组成以与层面垂直或高角度相交的层内潜穴、与层面平行的层内潜穴以及大量的层面爬行迹为特征(图6)。层内垂直潜穴常见3种类型: (1)与遗迹化石Skolithos(石针迹)类似的垂直管状潜穴和J形潜穴; (2)与遗迹化石Arenicolites(沙蠋迹)类似的U形潜穴; (3)与遗迹化石Psilonichnus(裸迹)类似的J形、Y形和复杂分枝形潜穴。层内垂直潜穴造迹生物主要是沙蚕、螃蟹、泥甲虫及其幼虫。与层面平行的表层潜穴主要有与遗迹化石Multina相似的潜穴, 造迹生物为蝼蛄、泥甲虫及其幼虫。层面上的运动迹主要是与遗迹化石Diplichnites(双趾迹)类似的螃蟹足辙迹以及层面上大量大小不一、形态各异的鸟类足迹。该遗迹组合指示了三角洲潮坪高潮区的沉积环境。
上述3种遗迹组合指示了中纬度季风区三角洲潮坪砂质沉积及高潮区和中低潮区砂泥互层沉积背景中的生物遗迹组合特征(图6)。现代生物遗迹的组成与分布, 与沉积底层的粒度、水动力条件、盐度、固结程度及食物来源等有着密切的关系。本研究可为古代三角洲造迹生物及其沉积环境的底层性质、水动力及水化学条件等生态环境因素研究提供实证材料, 既可作为研究古代三角洲潮坪沉积环境的判识依据, 也可为相似遗迹化石组合的沉积环境恢复提供佐证。
滦河三角洲潮坪上的现代生物遗迹在空间分布上存在不均衡性, 整体来看, 低潮坪水动力条件较强的砂质潮滩中生物遗迹的分异度和丰度较低, 中低潮区砂泥互层的沉积环境以及高潮区中生物遗迹的分异度和丰度较高。另外, 生物遗迹分布特征的精细差别还受到潮坪环境的盐度、水动力强度、粒度、TOC含量、沉积物含水率的影响。
盐度的变化和生物耐盐性是影响生物物种分布的关键因素之一。沙蚕(Nereis succinea)、螃蟹(Brachyura)均属于广盐性生物, 因此在研究区高、中、低潮滩均有分布, 而蝼蛄(Gryllotalpa)、泥甲虫(Coleoptera)及其幼虫仅分布于高潮区, 扁玉螺(Glossaulax didyma)、寄居蟹(Paguridae)仅分布于中、低潮区。受盐度变化影响, 研究区不同沉积环境中造迹生物分布不同: C处潮滩盐度相对较低, 受滦河河水作用和海水作用共同影响, 造迹生物包括沙蚕(Nereis succinea)、螃蟹(Brachyura)、扁玉螺(Glossaulax didyma)、蝼蛄(Gryllotalpa)、泥甲虫(Coleoptera)及其幼虫、鸟类, 生物遗迹组成是以与层面垂直或高角度相交的层内潜穴、与层面平行的层内潜穴以及大量的层面爬行迹为特征(图5), 整体上生物扰动程度较高, 生物遗迹的丰度和分异度也较高。距离滦河较近的A处潮滩以海水作用为主, 同时受到滦河河水影响, 盐度略低于海水标准盐度, 造迹生物包括沙蚕(Nereis succinea)、螃蟹(Brachyura)、寄居蟹(Paguridae)和鸟类, 生物遗迹是以层内各种形态潜穴和大量的层面拖迹为特征(图4), 丰度和分异度均较高, 与高潮区大体持平, 但在潮道两侧局部非常密集, 可能是受到潮水反复淹没的影响。B和E处的潮滩属于咸水环境, 造迹生物主要是喜盐性生物沙蚕(Nereis succinea)和螃蟹(Brachyura), 生物遗迹的丰度和分异度最低。
水动力、粒度及TOC含量三者之间是相互联系的, 其中水动力条件与沉积物粒度相关, TOC含量也与粒度特征相关, 三者对造迹生物的分布有一定的影响。研究区B和E处潮滩的水动力条件最强, 沉积物中泥质含量很少、粗砂含量高、TOC含量低、有机物较少, 因此生物数量较少, 生物遗迹的丰度和分异度均较低。位于河口附近的C处潮滩水动力条件相对较弱, A、D处潮滩水动力条件最弱, 沉积物中泥和细砂含量相对较高, TOC含量A处潮滩最高, 其次为C和D处潮滩, 丰富的有机质来源使得该处潮滩造迹生物数量较多, 因此生物遗迹的丰度和分异度均较高。
整体上, 研究区潮坪上生物遗迹分布具有分带性, 生物遗迹分布特征的精细差别还与沉积物含水率有关。沙蚕(Nereis succinea)在高、中、低潮区均有分布, 扁玉螺(Glossaulax didyma)主要分布于中低潮区砂泥质沉积物中, 但二者均在潮坪上低洼积水处边缘以及潮沟两侧反复被水淹没的地方密集分布, 这是由于该区域的底质含水率较高, 沉积物较软, 适合沙蚕(Nereis succinea)、扁玉螺(Glossaulax didyma)生存, 因此, 在潮坪上含水率较高的沉积物中沙蚕(Nereis succinea)、扁玉螺(Glossaulax didyma)丰度很高, 其营造的生物遗迹的丰度也最高。距离河道较近的高潮区, 水动力条件较弱, 含水量相对较低、半固结的泥质沉积物中, 适合蝼蛄(Gryllotalpa)、泥甲虫(Coleoptera)及其幼虫生存, 因此其营造的生物遗迹的丰度也较高。
三角洲潮坪环境容易受温度、盐度、水体能量和食物供应等的影响, 在这样动荡环境下生存的生物需要有较强的适应性, 其遗迹组合和分布特征亦有助于识别三角洲潮坪沉积环境。本次研究识别出的3种遗迹组合总体上来说与前人的研究成果(杨式溥等, 2004)一致, 但中低潮坪砂泥互层沉积环境中的生物扰动较砂质沉积中更强烈, 这可能与不同沉积环境的盐度、水动力条件、粒度、有机质含量和食物来源有关。此外, 本次研究进一步对三角洲潮坪环境受不同沉积物性质(盐度、水动力条件、粒度、TOC含量、含水率)影响的造迹生物种类、生物遗迹特征进行了分析, 为类比研究古潮坪的沉积环境提供了现代遗迹方面的实证材料。
1)观察到的滦河三角洲潮坪现代生物遗迹包括层面遗迹和层内潜穴。层面遗迹以节肢动物、环节动物、腹足动物的爬行迹为主, 还有觅食迹、停息迹、沙蚕粪粒和螃蟹掘穴过程产生的砂球, 造迹生物主要有沙蚕、螃蟹、扁玉螺、寄居蟹。层内潜穴以居住迹丰度最高, 且类型多样, 主要有垂直于层面或与层面高角度相交的管状、U形、Y形和多分枝形潜穴等, 造迹生物主要是螃蟹、沙蚕、蝼蛄、泥甲虫及其幼虫等。平行于层面分布的潜穴一般为蝼蛄、泥甲虫及其幼虫营造的潜穴。
2)基于生物遗迹的组合和分布特征以及沉积物底层的特点, 建立了3种不同沉积背景条件下的生物遗迹组合。遗迹组合I指示了三角洲潮坪水动力条件较强的砂质沉积环境, 以底层中Y形和垂直管状潜穴为特征, 局部丰度较高。遗迹组合Ⅱ指示了三角洲滨岸潮间带中低潮区砂泥互层沉积环境, 以层内各种形态潜穴和大量的层面爬行迹/拖迹为特征。遗迹组合Ⅲ 指示了三角洲潮坪高潮区砂泥互层沉积环境, 以与层面垂直或高角度相交的层内潜穴、与层面平行的层内潜穴以及大量的层面爬行迹/拖迹为特征。
3)在滦河三角洲潮坪不同沉积背景下, 造迹生物营造了不同形态和生态属性的生物遗迹, 其组成和分布与现代沉积环境之间具有明显的响应关系, 这可为类比研究古代三角洲造迹生物及其沉积环境的底层性质、水动力及水化学条件等生态环境因素提供实证材料。
(责任编辑 张西娟; 英文审校 刘贺娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|