




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
广西南宁下泥盆统华南鱼类化石新发现及其地层和古地理意义*
[张淑荣1, 2  , 山显任
, 山显任1, 2 , 盖志琨1, 2 ]
, 山显任, 盖志琨]
|
|

第一作者简介 张淑荣,女,1999年生,硕士研究生,从事人类活动和环境过程研究。E-mail: zhangshurong0929@126.com。
笔者描述了广西南宁下泥盆统布拉格阶那高岭组下部盔甲鱼类的新发现,建立了华南鱼科 2个新种——丘比特华南鱼(Huananaspis cupido sp. nov.)和梁氏亚洲鱼(Asiaspis liangi sp. nov.)。前者以头甲个体较小、不发育中背棘、两角末端超出头甲后缘等特征区别于属型种武定华南鱼(H. wudingensis),后者以头甲长大于宽、角向头甲侧后方延伸、角后缘发育锯齿、中背孔呈纵长椭圆形等特征区别于属型种宽展亚洲鱼(A. expansa)。广西南宁地区那高岭组具有可靠的地质时代证据,其中所产的徐家冲鱼类化石组合能够为华南地区早泥盆世地层的对比研究提供鱼类化石标准,而华南鱼属(Huananaspis)和亚洲鱼属(Asiaspis)在那高岭组的发现,丰富了华南鱼类的多样性,扩大了徐家冲鱼类组合的化石内容,并为云南武定坡松冲组与广西南宁那高岭组之间的对比提供了直接化石证据。广西南宁莲花山组与那高岭组过渡层中的徐家冲鱼类化石组合物种多样性最高,几乎涵盖了其他地区该组合的所有属种,说明广西南宁地区可能是盔甲鱼类辐射和扩散的中心之一。该地区早泥盆世布拉格期的古鱼类、早期维管植物和腕足动物均表现出显著的地方性色彩,可能跟华南海是一个半封闭的陆表海湾有关,即由于封闭海效应,鱼类在这个封闭的环境中独立演化,表现出很高的多样性和很强的土著性,最终在华南板块上形成一个独立的动物区系——“盔甲鱼—云南鱼”动物区系。
About the first author ZHANG Shurong,born in 1999,is a master’s degree candidate of Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences. She is engaged in the study of human activities and environmental process. E-mail: zhangshurong0929@126.com.
Two new species of Huananaspidae(Huananaspidiformes,Galeaspida), Huananaspis cupido sp. nov. and Asiaspis liangi sp. nov.,are described from the lower part of the Nagaoling Formation(Pragian,Early Devonian)in Nanning,Guangxi,China. H. cupido sp. nov. differs from H. wudingensis in smaller headshield,lacking median dorsal spine,and cornual processes extending beyond the posterior margin of the headshield. A. liangi sp. nov. differs from A. expansa in headshield length greater than width,cornual processes extending posterolaterally,serrated posterior margin of cornual processes,and longitudinal oval median dorsal opening. The Nagaoling Formation yields reliable index fossils indicating a Pragian age,thus the Xujiachong vertebrate assemblage yielded from this formation can provide a palaeoichthyological standard for stratigraphic correlations of Early Devonian in South China. New findings of Huananaspis and Asiaspis from the Nagaoling Formation enriches the biodiversity of huananaspids,adds new members to the the Xujiachong vertebrate assemblage,and provides direct fossil evidence for the correlation between the Posongchong Formation in Wuding of Yunnan and the Nagaoling Formation in Nanning of Guangxi. The Xujiachong vertebrate assemblage is the most diversified in the transitional beds between the Lianhuashan and Nagaoling formations,almost containing all genera in adjacent regions,indicating that Naning of Guangxi is one of the centers for galeaspids radiating and dispersing. Fish assemblage displays a similar high level of endemism to that of the vascular plants and brachiopod fauna during the Pragian of Early Devonian,which probably caused by the South China sea,a large semi-closed epicontinental gulf. Because of the effect of a closed sea,fishes show exceptional taxonomic diversity and high levels of endemism,forming an independent biogeographic region known as‘galeaspid-yunnanolepid province’ in South China.
广西南宁地区的泥盆系发育良好, 是中国泥盆系研究的关键区域之一。该区的下泥盆统自下而上划分为莲花山组、那高岭组、郁江组、莫丁组和那叫组(图1), 5个组之间均呈整合接触关系, 代表了一个连续的海侵沉积序列(王钰等, 1964; 邝国敦等, 1989; 钟铿等, 1992)。六景剖面是中国著名的海相泥盆系经典剖面之一, 位于南宁市中心往东约50 km的横县六景镇附近(图1-A)。该剖面全长4.6 km, 层序完整, 露头良好, 下泥盆统除了产出丰富的无脊椎动物化石外, 还产出丰富的早期脊椎动物化石, 主要包括盔甲鱼类: 莲花山真盔甲鱼(Eugaleaspis lianhuashanensis)(刘时藩, 1986)、宽展亚洲鱼(Asiaspis expansa)(潘江等, 1975)和角箭甲鱼(Antiquisagittaspis cornuta)(刘玉海, 1985); 胴甲类: 计氏云南鱼(Yunnanolepis chii)(潘江和王士涛, 1978; 刘时藩, 1992)、秀丽曲靖鱼(Chuchinolepis gracilis)(刘时藩, 1992)和沾益鱼未定种(Zhanjilepis sp.)(刘时藩, 1992); 节甲类: 多瘤亚洲棘鱼(Asiacanthus multituberculatus)(刘时藩, 1982a)、六景郁江鱼(Yujiangolepis liujingensis)(王士涛等, 1998; Dupret et al., 2009)和北极鱼未定种(Szelepis sp.)(刘时藩, 1982b)。
 | 图1 广西南宁下泥盆统剖面位置及地层柱状图 A— 地理位置图, 示五象岭、仙葫、五合、六景剖面的地理位置; B— 六景剖面下泥盆统地层柱状图, 含鱼化石层位及腕足类、牙形类地层分布(Lu et al., 2016; Qie et al., 2018; Yu et al., 2019; Guo et al., 2019); C— 五合剖面郁江组野外剖面照片; D— 五象岭剖面那高岭组野外 剖面照片(本文化石产地); E— 仙葫剖面莲花山组野外剖面照片Fig.1 Location map of sections and stratigraphic column of the Lower Devonian in Nanning, Guangxi |
令人遗憾的是, 自1983年六景剖面被列为省级地质遗址保护区以来, 在该剖面开展野外挖掘已经十分困难, 因此在近20年的时间里, 该地区鲜有早期脊椎动物化石被发现和报道, 亟需寻找新的替代剖面开展野外挖掘工作。随着南宁的城镇化和房地产开发, 在南宁至六景之间暴露出许多人工开凿的临时剖面。自2010年以来, 笔者在该地区开展了大量的野外调查工作, 陆续在南宁的仙葫、五象岭和五合等地分别发现了莲花山组、那高岭组和郁江组3个新剖面(图1-C, 1-D, 1-E), 并抢救性地挖掘出大量早期脊椎动物化石, 主要包括无颌类的盔甲鱼类, 盾皮类的胴甲类、节甲类、瓣甲类以及肉鳍鱼类的杨氏鱼属(Youngolepis)和斑鳞鱼属(Psarolepis)(曾广春等, 2018)。在上述化石中, 除了Qiao和Zhu(2015)报道了五合郁江组的肺鱼— — 曾氏卡思尔斯喙鱼(Cathlorhynchus zengi)和Gai等(2018)报道了五象岭那高岭组的多鳃鱼类— — 曾氏南宁鱼(Nanningaspis zengi)外, 其他化石均尚未报道。文中报道了在广西南宁恒大城施工工地(图1-A的五象岭剖面)那高岭组下部发现的盔甲鱼类华南鱼科2个新种, 新材料的发现丰富了早泥盆世华南鱼类的多样性, 扩大了徐家冲组合的化石内容和古地理分布, 并为邻区早泥盆世含鱼地层的对比提供了进一步的参考。
盔甲鱼亚纲Galeaspida Tarlo, 1967
多鳃鱼超目Polybranchiaspidida Janvier, 1996
华南鱼目Huananaspidiformes Janvier, 1975
华南鱼科Family Huananaspidae Liu, 1973
华南鱼属HuananaspisLiu, 1973
属型种 武定华南鱼Huananaspis wudingensisLiu, 1973
特征(修订) 头甲呈头盔形, 长大于宽, 吻缘延伸为狭长的吻突; 角发育, 侧向延伸, 呈略向后弯的窄镰刀形, 角的末端与头甲后缘大致齐平或超出头甲后缘, 头甲最宽处位于两角末端; 中背孔呈心脏形, 后缘向前凹进, 横宽稍大于纵长; 眶孔侧位, 于头甲侧缘呈缺刻状; 侧线系统不详; 纹饰为粒状突起; 鳃囊约为10对。
丘比特华南鱼 Huananaspis cupido sp. nov.
词源 cupido, 拉丁词源, 意为爱神丘比特, 取自新种的中背孔形态呈心形, 形似丘比特之箭。
正模 一件保存完整的头甲外模 IVPP V26674。
产地与层位 中国广西壮族自治区南宁市五象岭, 下泥盆统布拉格阶那高岭组底部。
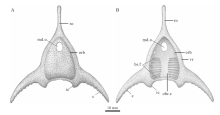
种征 头甲呈头盔形, 长约59.0 mm, 宽约77.0 mm, 侧缘光滑; 吻突发育, 呈狭长棒状, 长约26.0 mm; 头甲后缘平直, 中背脊和中背棘均不发育; 角发育, 侧向延伸, 呈前缘凸、后缘凹的镰刀型, 角的末端超出头甲后缘; 中背孔呈心脏形, 横宽大于纵长; 眶孔侧位, 较小; 纹饰由细小而稀疏的粒状突起组成, 每平方毫米有4~6个突起; 鳃囊约为10对; 松果孔及侧线系统不详。
测量 见表1。
| 表1 丘比特华南鱼(新种)和梁氏亚洲鱼(新种)标本测量(mm) Table1 Measurements of Huananaspis cupido sp. nov. and Asiaspis liangi sp. nov.(mm) |
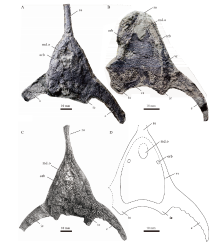
描述 正型标本(IVPP V26674)为一件近乎完整保存的头甲外模, 仅右侧角的末端有所缺失(图2-A)。头甲吻缘向前延伸形成狭长的吻突, 后缘向两侧延伸形成一对角, 使得头甲整体形态呈头盔形。头甲最宽处位于两角末端, 最大宽度为77.2 mm, 最大长度为58.5 mm, 头甲中线长(吻突前缘至头甲后缘的距离)为49.1 mm(表1)。吻突呈细长的棒状, 长度(吻突前缘至中背孔前缘的距离)为26.3 mm, 中部宽3.1 mm。角呈前缘凸、后缘凹的镰刀型, 向头甲侧后方弯曲, 较窄。角的长度(角的基部至角的末端之间的直线距离)为29.2 mm, 中部宽度为2.9 mm, 两角末端之间的距离为77.2 mm(表1)。角的末端远远超过头甲后缘, 头甲后缘至两角末端水平连线之间的距离为9.4 mm, 约占头甲总长度的1/5。头甲后缘较为平直, 不发育中背脊和中背棘(图2-A)。
 | 图2 丘比特华南鱼(新种)正型标本照片和解释性素描 A— 1件近乎完整的头甲外模, 正型标本, IVPP V26674; B— 正型标本的解释性素描图(许勇绘)。简写说明: br.f— branchial fossa(鳃穴); c— cornual process(角); md.o— median dorsal opening(中背孔); obr.c— oralobranchial chamber(口鳃腔); orb— orbital opening(眶孔); ro— rostral process(吻突)Fig.2 Photograph and interpretative drawing of the holotype of Huananaspis cupido sp. nov. |
中背孔保存完好, 位于头甲前端, 前缘向前凸出, 后缘中部向前凹进, 整体呈心形(图2-A, 2-B)。中背孔的纵长为4.4 mm, 横宽为5.6 mm。眶孔位于中背孔之后的头甲侧缘, 其前缘与中背孔后缘大致齐平, 背视呈缺刻状(图2-A; 图3-A), 长轴长约2.5 mm, 两眶孔之间的距离为15.2 mm(表1)。松果孔和感觉管系统在正型标本中均未被保存。
 | 图3 丘比特华南鱼(新种)头甲复原图(许勇绘) A— 头甲背侧; B— 头甲腹侧。 vr— ventral rim(腹环); br.o— branchial opening(鳃孔); 其他简写说明见图2Fig.3 Restoration of the headshield of Huananaspis cupido sp. nov.(artwork credit: Xu Yong) |
头甲侧缘光滑, 不发育锯齿。纹饰由均匀分布的粒状突起组成, 排列较稀疏, 每平方毫米约有4~6个突起。头甲外模中央可见2个对称分布的纵向椭圆形凹陷, 指示了容纳鳃囊的鳃区(图2-A, 2-B; 图3-B), 鳃区纵长约14.2 mm, 横宽约8.3 mm, 左侧鳃区可见10个左右纵向排列的横条状凹陷, 为鳃囊在保存过程中在头甲背侧产生的印痕。
比较与讨论 新材料头甲呈头盔形、吻端发育细长的吻突、头甲后侧发育侧向延伸的角、中背孔呈心形、眶孔侧位, 可被归入华南鱼目下的华南鱼科(Huananaspidae)(刘玉海, 1973)。华南鱼科由刘玉海(1973)依据华南鱼属(Huananaspis)建立, 该科的其他成员包括亚洲鱼属(Asiaspis)、南盘鱼属(Nanpanaspis)、龙门山鱼属(Lungmenshanaspis)、大窗鱼属(Macrothyraspis)、中华四川鱼属(Sinoszechuanaspis)、箐门鱼属(Qingmenaspis)和王冠鱼属(Stephaspis)(刘玉海, 1965; 潘江等, 1975; 潘江和王士涛, 1978, 1981; Pan, 1992; 盖志琨和朱敏, 2007)。其中, 后4者因头甲背侧具有背窗或侧背窗而又被归入大窗鱼亚科(Macrothyraspinae)(Pan, 1992; Zhu and Gai, 2006)。新材料头甲不发育背窗或侧背窗, 可与大窗鱼科成员明显区分。在中背孔形状方面, 新材料的中背孔呈心形, 与华南鱼属和龙门山鱼属的中背孔较为相似, 而与亚洲鱼属和南盘鱼属所具有的亚圆形或卵圆形中背孔明显不同。在眶孔位置方面, 新材料的眶孔位于头甲侧缘, 与华南鱼属的眶孔位置较为相似, 而与龙门山鱼属所发育的背位眶孔差异显著。此外, 新材料的角呈略向后弯曲的窄镰刀形, 也属于华南鱼属的典型特征。因此, 笔者将新材料归入华南鱼属。
然而, 新材料与属型种武定华南鱼(H. wudingensis)在以下几个方面存在显著差异: (1)个体大小存在差异, 新材料的头甲最大宽度为77.2 mm, 最大长度为58.5 mm, 而武定华南鱼的头甲最大宽度为150.0 mm, 最大长度为96.0 mm, 后者头甲宽度约为前者的2倍, 长度约为前者的1.6倍; (2)新材料的头甲后缘平直, 不发育中背棘, 而武定华南鱼的头甲后缘向后突出形成中背棘; (3)新材料的两角末端远远超过其头甲后缘, 而武定华南鱼的两角末端未超出头甲后缘。基于以上对比, 笔者认为新材料与属型种武定华南鱼的特征差别明显, 故将其命名为华南鱼属下一新种(图6)。
 | 图6 丘比特华南鱼(新种)(右)和梁氏亚洲鱼(新种)(左)生态复原图(史爱娟绘)Fig.6 Life restoration of Huananaspis cupido sp. nov.(right)and Asiaspis liangi sp. nov.(left)(artwork credit: Shi Aijuan) |
亚洲鱼属AsiaspisP’ an et al., 1975
属型种 宽展亚洲鱼Asiaspis expansaP’ an et al., 1975
特征(修订) 头甲呈头盔形, 两侧缘约呈60° 夹角, 头甲后缘中部前凹, 不具中背棘, 头甲最宽处位于两角末端; 吻突狭长, 前部渐尖, 横切面为扁圆形; 角向侧方或侧后方延伸, 棘状, 末端超过头甲后缘水平线, 角内缘光滑或发育锯齿; 内角较短, 呈倒三角形, 向后延伸; 中背孔呈亚圆形或纵长椭圆形; 眶孔背位, 较小, 呈椭圆形, 靠近头甲侧缘; 侧线系统为多鳃鱼型, 目前仅知中背纵管中只有“ V” 形后眶上管发育, 侧背纵管发育, 主侧线上保存2对侧横管和背联络管; 纹饰由具粒状突起组成; 鳃囊11对。
梁氏亚洲鱼Asiaspis liangi sp. nov.
词源 liang, “ 梁” 的汉语拼音, 赠予化石标本发现者梁江先生。
正模 一件保存不完整的头甲内外模 IVPP V26673a, V26673b。
产地与层位 中国广西壮族自治区南宁市五象岭, 下泥盆统布拉格阶那高岭组底部。
种征 头甲呈头盔形, 长约96.0 mm, 宽约90.0 mm, 侧缘光滑; 吻突发育, 呈狭长棒状, 长约35.0 mm; 头甲后缘略向前凹进, 呈拱形, 中背脊和中背棘均不发育; 角和内角均较为发育, 其夹角约为90° ; 角向侧后方延伸, 前缘光滑, 后缘发育细小锯齿, 两角末端超出头甲后缘; 内角呈倒三角形, 较短, 指向后方; 中背孔呈纵长椭圆形, 纵长约为横宽的1.6倍; 眶孔背位, 较小, 靠近头甲侧缘, 其前缘与中背孔后缘大致齐平; 纹饰由稀疏分布的粗大瘤状突起组成, 每平方毫米有1~2个突起; 松果孔、侧线系统及鳃囊数量不详。
测量 见表1
描述 正型标本(IVPP V26673)为一件近乎完整保存的头甲及其外模, 仅左侧角的末端有所缺失(图4-A, 4-B)。头甲呈头盔形, 最宽处位于两角末端, 最大宽度为90.1 mm, 最大长度为95.7 mm, 头甲中线长76.2 mm(表1)。头甲吻缘向前延伸形成细长的棒状吻突, 吻突在中背孔前缘向前8.0 mm处迅速变尖, 长35.0 mm, 中部宽3.6 mm。角和内角均较为发育, 位于头甲侧后方(图4-A, 4-B)。角向侧后方延伸, 由基部向末端逐渐变窄, 角和内角之间的夹角约为90° 。角的前缘光滑, 后缘发育小的锯齿。两角末端远远超过头甲后缘, 头甲后缘至两角末端水平连线之间的距离为19.5 mm, 约占头甲总长度的1/5。角的基部至角的末端之间的直线距离为35.7 mm, 两角末端之间的距离为90.1 mm(表1)。内角呈三角形, 较短, 指向后方, 两内角末端之间的距离为24.2 mm。2个内角之间的头甲后缘略向前凹进, 呈拱形, 不发育中背棘和中背脊(图4-B, 4-D; 图5-A, 5-B)。
 | 图4 梁氏亚洲鱼(新种)正型标本照片和解释性素描 A— 1件近乎完整的头甲内模, 正型标本, IVPP V26673a; B— 1件近乎完整的头甲外模, 正型标本, IVPP V26673b; C— 正型标本内模的解释性素描图(曾广春绘); D— 正型标本外模的解释性线图。ic— inner cornual process(内角); 其他简写说明见图2和图3Fig.4 Photographs and interpretative drawing of the holotype of Asiaspis liangi sp. nov. |
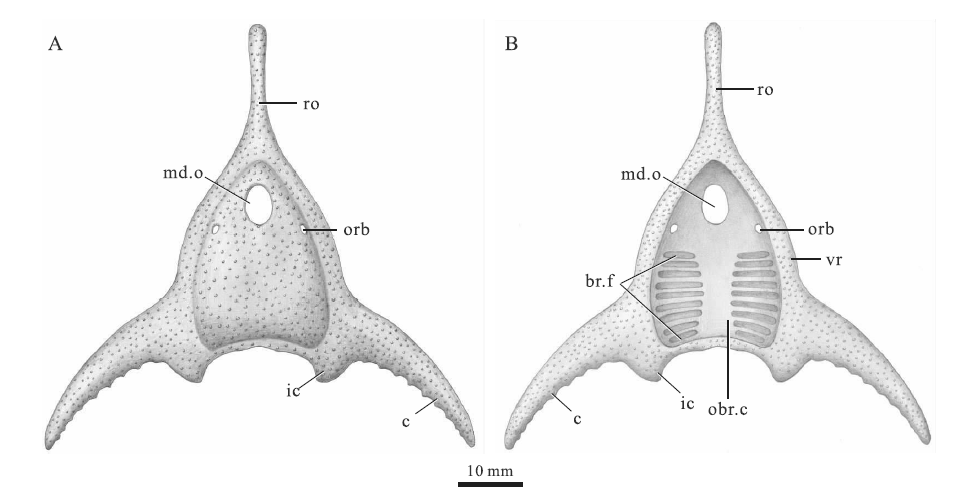 | 图5 梁氏亚洲鱼(新种)头甲复原图(许勇绘) A— 头甲背侧; B— 头甲腹侧; 简写说明见图2、 图3和图4Fig.5 Restoration of the headshield of Asiaspis liangi sp. nov.(Artwork credit: Xu Yong) |
中背孔在正型标本的外模中保存较好(图4-B, 4-D), 位于头甲前部中央, 呈纵长椭圆形, 长轴与头甲中线重合, 纵长9.4 mm, 横宽5.8 mm, 纵长约为横宽的1.6倍(表1)。眶孔背位, 位于中背孔后侧方(图4), 眶孔前缘与中背孔后缘大致齐平, 眶孔侧缘距离头甲侧缘约4.0 mm。眶孔呈纵长椭圆形, 较小, 长轴长为3.5 mm, 短轴长为2.0 mm, 2个眶孔之间的距离为16.6 mm(表1)。松果孔和感觉管系统在正型标本中均未被保存。
纹饰由粗大的瘤状突起组成, 分布稀疏, 每平方毫米有1~2个突起(图4-A, 4-B)。在正型标本的外模中(图4-B), 头甲后缘和侧缘可见部分腹环, 腹环由角的基部向头甲前方逐渐变窄, 包围着头甲中央的口鳃腔(图5-B)。沿腹环内侧可见2个对称分布的纵向椭圆形凹陷, 指示了容纳鳃囊的鳃区, 鳃区纵长约20.2 mm, 横宽约10.0 mm, 但鳃孔或鳃囊的印痕均未被保存。
比较与讨论 新材料头甲呈头盔形、吻端发育细长的吻突、头甲后侧发育侧向延伸的角、中背孔呈卵圆形, 可被归入华南鱼目下的华南鱼科。新材料头甲不具有背窗或侧背窗, 可与华南鱼科中的大窗鱼亚科成员(大窗鱼属、中华四川鱼属、箐门鱼属和王冠鱼属)明显区分。就头甲形状和角的位置而言, 新材料的头甲呈头盔形, 角位于头甲基部并向侧方延伸, 可与南盘鱼属明显区分, 后者的头甲略呈五边形, 角位于头甲中部。就中背孔形状而言, 新材料的中背孔呈卵圆形, 与亚洲鱼属的中背孔形状较为相似, 而与华南鱼属和龙门山鱼属所具有的心形中背孔明显不同。此外, 新材料的眶孔位于头甲背侧, 头甲发育内角, 角向侧后方弯曲, 均属于亚洲鱼属的典型特征。因此, 笔者将新材料归入亚洲鱼属。
亚洲鱼属由潘江等(1975)依据宽展亚洲鱼(A. expansa)建立, 新材料与宽展亚洲鱼在以下几个方面存在显著差异: (1)头甲比例。新材料的头甲长95.7 mm, 宽90.1 mm, 长略大于宽, 而宽展亚洲鱼的头甲长80.0~85.0 mm, 宽105.0~110.0 mm, 宽大于长; (2)角的延伸方向。新材料的角向头甲侧后方延伸, 角和内角之间的夹角约为90° , 而宽展亚洲鱼的角向头甲侧方延伸, 仅末端略向后弯曲, 角和内角之间的夹角约为120° ; (3)锯齿发育情况。新材料的角后缘发育锯齿, 而宽展亚洲鱼的角后缘圆滑, 不发育锯齿; (4)中背孔形态。新材料的中背孔呈纵长椭圆形, 纵长与横宽之比约为2.5, 而宽展亚洲鱼的中背孔为亚椭圆形, 横宽稍大于纵长, 二者之比约为1。基于以上对比, 笔者认为新材料与属型种宽展亚洲鱼差异明显, 作为亚洲鱼属一新种成立(图6)。
广西南宁附近的早期鱼类化石主要采自莲花山组紫红色砂岩(图1-E)和那高岭组灰绿色砂岩(图1-D)。该区下泥盆统主要有4个含鱼层位, 分别是莲花山组中下部、莲花山组与那高岭组的过渡层、那高岭组上部和郁江组中部石洲段(图-B), 其中莲花山组顶部六坎口段与那高岭组底部的高岭段为海陆交互相的连续沉积, 在野外工作中, 对于二者的分界线经常难以区分, 因此笔者把两者的含鱼层位统称为莲花山组与那高岭组的过渡层, 时代上应属于同一时期。广西地区莲花山组中下部的横县段和伶俐段产胴甲类计氏云南鱼(Y. chii)、秀丽曲靖鱼(C. gracilis)和沾益鱼未定种(Zhanjilepis sp.)(刘时藩, 1992), 这几个属种均首次发现报道于滇东下泥盆统西屯组, 属于翠峰山组合(早期脊椎动组合Ⅱ )的常见分子(Zhu et al., 2000), 表明莲花山组中下部大致与滇东的西屯组相当, 在时代上属于早泥盆世洛赫考夫期中晚期。但广西地区莲花山组未见滇东下泥盆统西山村组中常见的西山村组合(早期脊椎动组合I)的分子, 特别是未见盔甲鱼类和多鳃鱼类的大量出现。在广西地区, 莲花山组直接与下伏寒武系呈不整合接触, 而在滇东地区, 西屯组之下的西山村组与志留系普里道利阶上部玉龙寺组为整合接触的连续沉积。因此, 广西地区的莲花山组很可能缺失与滇东地区西山村组相当的洛赫考夫期早期的沉积(刘时藩, 1992)。
早期鱼类化石, 特别是无颌类盔甲鱼化石, 在莲花山组与那高岭组的过渡层突然变得丰富起来。目前已发表的盔甲鱼类化石主要有真盔甲鱼科的真盔甲鱼属(Eugaleaspis)、古木鱼科的南宁鱼属(Nanningaspis)、华南鱼科的亚洲鱼属(Asiaspis)和三岔鱼科的箭甲鱼属(Antiquisagittaspis), 以及尚未进行系统描述但分别与多鳃鱼科的多鳃鱼属(Polybranchiaspis)和东方鱼属(Dongfangaspis)、三岔鱼科的三岔鱼属(Sanchaspis)、三歧鱼科的三岐鱼属(Sanqiaspis), 鸭吻鱼科的鸭吻鱼属(Gantarostrataspis)、裂甲鱼属(Rhegmaspis)和乌蒙山鱼属(Wumengshanaspis)及大窗鱼亚科的大窗鱼属(Macrothyraspis)、龙门山鱼属(Lungmenshanaspis)和箐门鱼属(Qingmenaspis)非常相似的新材料(Gai et al., 2022)。上述新材料中, 除真盔甲鱼属、多鳃鱼属和三岔鱼属新材料采自南宁仙葫莲花山组顶部的紫红色砂岩外, 其余均采自南宁五象岭那高岭组底部的灰绿色砂岩中。文中描述的华南鱼科2个新种— — 丘比特华南鱼(Huananaspis cupido sp. nov.)和梁氏亚洲鱼(Asiaspis liangi sp. nov.)也均产自那高岭组底部, 而华南鱼属此前仅见于云南武定地区的坡松冲组(刘玉海, 1973)。
云南武定地区的坡松冲组为一套总厚不到100 m的碎屑砂岩, 曾被称为“ 翠峰山组” (刘玉海, 1975), 之后改称为坡松冲组, 并沿用至今(廖卫华等, 1978; 王俊卿和朱敏, 1994; 朱敏等, 1994; 王俊卿等, 1996)。这套砂岩直接不整合于寒武系或奥陶系之上, 与上覆的坡脚组呈整合接触。坡松冲组主要分布于滇东南的文山、广南、滇东北的昭通以及滇中的武定地区(朱敏等, 1994; 王俊卿等, 1996)。在昭通地区, 坡松冲组产有盔甲鱼类: 三歧鱼属(Sanqiaspis)、剑裂甲鱼属(Rhegmaspis)、昭通鱼属(Zhaotongaspis)、大窗鱼属(Macrothyraspis)、龙门山鱼属(Lungmenshanaspis)和箐门鱼属(Qingmenaspis)(潘江和王士涛, 1981; Pan, 1992; 王俊卿和朱敏, 1994; 王俊卿等, 1996; Gai et al., 2015); 盾皮鱼类: Yiminaspis和短瓣鱼属(Brevipetalichthys)(Dupret, 2008; 姬书安和潘江, 1999); 以及肉鳍鱼类: 多孔弓鱼属(Arquatichthys)、东生鱼属(Tungsenia)和箐门齿鱼属(Qingmenodus)(Lu and Zhu, 2008, 2010; Lu et al., 2012)。在文山地区, 坡松冲组产有盔甲鱼类: 三歧鱼属(Sanqiaspis)、鸭吻鱼属(Gantarostrataspis)、文山鱼属(Wenshanaspis)、大窗鱼属(Macrothyraspis)和古木鱼属(Gumuaspis)(王俊卿和王念忠, 1992; 赵文金等, 2002; 王俊卿等, 2005)。然而, 武定地区坡松冲组中的鱼类化石较少, 目前仅产有盔甲鱼类华南鱼属(Huananaspis), 该属虽然在科一级的分类阶元上能够和其他地区的鱼类化石进行对比, 如广西南宁那高岭组中的亚洲鱼属(Asiaspis), 但在属一级的分类阶元上却缺少可与之直接对比的化石。
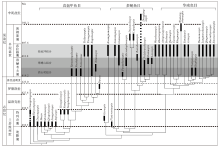
上述2个地区所产的盔甲鱼类均以带吻突的华南鱼类为主, 这些脊椎动物化石被称为Sanchaaspis-Asiaspis组合(早期脊椎动组合Ⅲ )(Zhu et al., 2000)或徐家冲组合(Zhao and Zhu, 2010)。早泥盆世是盔甲鱼类在华南地区发生快速辐射演化的关键时期, 因此盔甲鱼类在华南地区早泥盆世区域地层对比问题上能够发挥重要作用。盔甲鱼类在早泥盆世经历了3个主要的演化阶段, 被归入3个脊椎动物化石组合, 分别是洛赫考夫期的西山村组合(早期脊椎动组合I)、翠峰山组合(早期脊椎动组合Ⅱ )和布拉格期的徐家冲组合(早期脊椎动组合Ⅲ )(图7)(Zhao and Zhu, 2010, Zhu et al., 2000)。其中, 洛赫考夫期的盔甲鱼类以多鳃鱼目和真盔甲鱼目的辐射演化为主要特征, 仅有零星的华南鱼目成员开始出现。到了布拉格期, 盔甲鱼类中的华南鱼目开始大量出现并快速辐射, 且这一时期的真盔甲鱼目和华南鱼目成员以及部分多鳃鱼目成员均平行进化出了吻突结构, 因此徐家冲组合中发育吻突的盔甲鱼类具有重要的生物地层意义(Gai et al., 2022)。徐家冲组合成员目前在广西南宁的莲花山组与那高岭组过渡层、云南曲靖徐家冲组、四川江油关山坡组、云南文山、昭通和武定的坡松冲组及越南Si Ka组都有发现(图8), 而以广西南宁莲花山组与那高岭组过渡层中的徐家冲组合物种多样性最高, 笔者近几年在该层位发现的新材料几乎涵盖了可与其他地区对应的所有属种, 尤其是盔甲鱼类几乎能够在属一级的分类水平上与其他地区的成员形成一一对比(Gai et al., 2022)。例如, 三岔鱼科的三岔鱼属(Sanchaspis)新材料与发现于云南曲靖徐家冲组的宽大吻突三岔鱼(S. magalarotrata)非常相近; 鸭吻鱼科的鸭吻鱼属(Gantarostrataspis)新材料与发现于云南曲靖徐家冲组和文山坡松冲组的耿氏鸭吻鱼(G. gengi)非常相近; 乌蒙山鱼属(Wumengshanaspis)新材料与发现于云南彝良缩头组的寸田乌蒙山鱼(W. cuntianensis)非常接近; 裂甲鱼属(Rhegmaspis)新材料与发现于云南昭通坡松冲组的剑裂甲鱼(R. xiphoidea)非常接近; 大窗鱼亚科的大窗鱼属(Macrothyraspis)新材料与发现于云南文山、昭通坡松冲组的长角大窗鱼(M. longicornis)以及四川江油关山坡组的中华四川鱼(Sinoszechuanaspis)非常接近; 龙门山鱼(Lungmenshanaspis)新材料与发现于四川江油关山坡组的江油龙门山鱼(L. kiangyouensis)以及云南昭通坡松冲组的云南龙门山鱼(L. yunnanensis)非常相近; 多鳃鱼科的东方鱼属(Dongfangaspis)新材料与发现于四川江油关山坡组的硕大东方鱼(D. major)非常相近; 古木鱼科的南宁鱼属(Naningaspis)与发现于云南文山坡松冲组的古木鱼属(Gumuaspis)非常接近(Gai et al. 2022)。
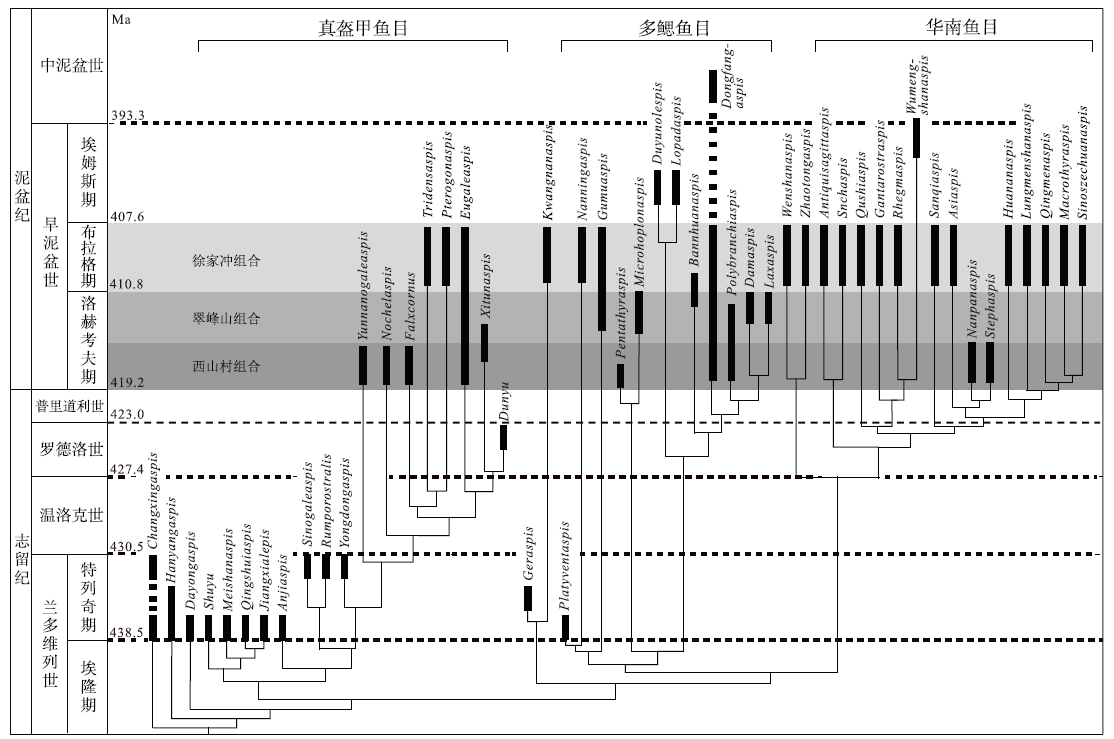 | 图7 盔甲鱼类的系统发育关系与地史分布(修改自Zhu and Gai, 2006)Fig.7 Phylogenetic relationship and temporal distribution of galeaspid genus(modified from Zhu and Gai, 2006) |
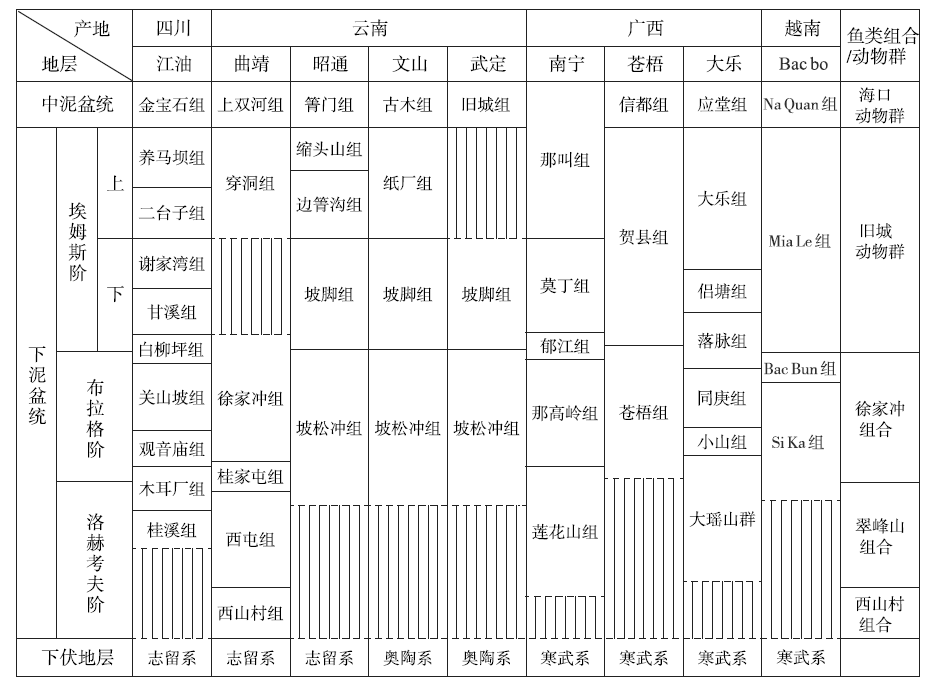 | 图8 华南下泥盆统鱼类化石组合及含鱼地层对比(修改自Zhao and Zhu, 2010)Fig.8 Stratigraphic map showing vertebrate assemblages and correlations of fish-bearing formations of the Lower Devonian in South China(modified from Zhao and Zhu, 2010) |
另外, 三岐鱼科的三歧鱼属(Sanqiaspis)更是徐家冲组合的典型分子, 该属同时出现在广西南宁那高岭组、云南昭通和文山地区的坡松冲组、曲靖地区的徐家冲组、四川江油地区的关山坡组以及越南北部的Si Ka组, 为这些地层之间的直接对比提供了属一级的化石证据(Gai et al., 2022)。特别是越南Si Ka组的越南三歧鱼(S. vietnamensis), 因其与多鳃鱼属(Polybranchiaspis)共生在一起, Janvier等(2009)认为其出现的时代比中国的三歧鱼出现的时代要稍早一些, 属于早泥盆世洛赫考夫期的西山村组合(早期脊椎动组合I)(图8)。但是, 广西南宁莲花山组与那高岭组过渡层中多鳃鱼科的多鳃鱼属(Polybranchiaspis)和东方鱼属(Dongfangaspis)的发现表明多鳃鱼属的延续时代较长, 可能从早泥盆世洛赫考夫期一直延续到布拉格期, 而东方鱼属(Dongfangaspis)甚至可能延伸到中泥盆世的海口组沉积时期(Meng et al., 2022)。
在滇东地区, 坡松冲组的上覆地层坡脚组中产有东京石燕动物群(‘ Spirifer’ tonkinensis Fauna), 该动物群在滇东地区的时代为埃姆斯期早期, 表明坡松冲组的时代可能为布拉格期(郝守刚, 1988; 刘时藩, 1994; 王俊卿等, 1996; Guo et al., 2019)。王怿(1994)报道了产自云南文山坡松冲组和坡脚组的孢子组合, 并通过和国内外相应组合的对比认为这2个组的时代分别为布拉格期和埃姆斯期早期。在广西南宁那高岭组下部, 与徐家冲鱼类组合共生的化石包括牙形刺Eognathodus kuangi(E. sulcatus)、E. nagaolingensis、Polygnathus trilinearis和Pandorinellina exigua philipi, 其延限可能位于布拉格期的Eognathodus kindlei带之内(Lu et al. 2016)。此外, 那高岭组上覆地层郁江组下部产有牙形刺Polygnathus pireneae、P. skolovi和东京石燕动物群(‘ Spirifer’ tonkinensis Fauna), 其时代分别为布拉格期和布拉格期至埃姆斯期(图1-B)(Yu et al., 2018; Guo et al., 2019; Qie et al., 2019)。因此, 广西南宁地区那高岭组具有可靠的地质时代证据, 其中的徐家冲鱼类组合能够为华南地区早泥盆世地层的对比研究提供鱼类化石标准。华南鱼属(Huananaspis)在广西南宁那高岭组中的首次发现, 为其与滇东武定地区同样产华南鱼属(Huananaspis)的坡松冲组的对比提供了直接的化石证据(图8)。
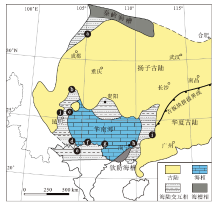
目前徐家冲鱼类组合的9个产地和层位, 分别是:广西南宁崇左地区的那高岭组, 云南曲靖地区的徐家冲组, 云南昭通、文山和武定地区的坡松冲组, 四川江油地区的关山坡组, 广西梧州地区的苍梧组, 越南北部的Khao Loc组和Si Ka组, 在古地理构造上均属于由扬子古陆和华夏古陆组成的华南板块。早泥盆世, 华南板块位于低纬度古赤道地区, 并被古特提斯洋与古太平洋从劳亚古大陆和冈瓦纳大陆隔离开来。在扬子古陆和华夏古陆之间存在一个陆表海湾, 即华南海(图9)(Young and Janvier, 1999; Torsvik and Cocks, 2004; Ma et al., 2009; Zhao and Zhu, 2010; Huang et al., 2018; Xue et al., 2018)。洛赫考夫期, 中国南部发生海侵, 海水从西南方向向广西中部和西部侵入。布拉格期, 在广西造山运动之后, 华南海覆盖了华南板块南缘的大部分地区, 这使得华南板块广泛发育了近岸含化石的硅质碎屑岩和石灰岩沉积。广西六景剖面的那高岭组除了含有丰富的鱼类化石外, 还含有珊瑚、菊石、胞石、腕足类、牙形石等多门类海相化石, 指示了浅海相沉积(钟铿等, 1992; Ma et al., 2009), 而云南曲靖徐家冲组、昭通坡松冲组、文山坡松冲组、四川江油关山坡组、广西梧州苍梧组、越南北部Khao Loc组和Si Ka组均为华南海附近的近岸至三角洲相沉积, 为这一时期的植物、鱼类和腕足类化石的保存提供了有利的沉积环境。在这一时期, 古鱼类组合、早期维管植物组合和腕足动物东京石燕(‘ Spirifer’ tonkinensis)组合表现出显著的地方性色彩(Tô ng Dzuy et al., 1996; Zhao and Zhu, 2010; Xue et al., 2018; Zhang and Ma, 2019), 和世界其他地方的动物群面貌明显不同, 比如尽管在澳大利亚、北美、欧洲和西伯利亚等地有大量早泥盆世鱼类化石发现, 但它们与中国华南板块的早泥盆世古鱼类面貌明显不同(Tô ng Dzuy et al., 1996), 特别是华南板块的无颌类盔甲鱼类和盾皮鱼类云南鱼类化石, 呈现出很浓厚的土著性色彩。Young(1981, 1993)将早泥盆世华南板块的古鱼类(尤其是盔甲鱼类和云南鱼类)作为一个独立的动物区系, 并命名为“ 盔甲鱼— 云南鱼” 动物区系(galeaspid-yunnanolepid province)。
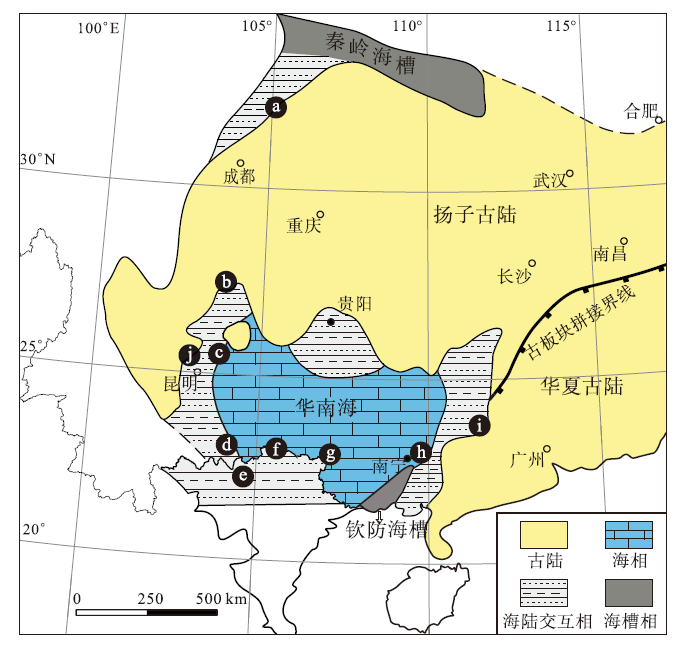 | 图9 华南板块早泥盆世古地理图及徐家冲组合的分布 (修改自Gai et al., 2022) a— 四川江油关山坡组; b— 云南昭通坡松冲组; c— 云南曲靖徐家冲组; d— 云南文山坡松冲组; e— 越南河江Khao Loc组; f— 越南安明Si Ka组和Bac Bun组; g— 广西崇左莲花山组和那高岭组; h— 广西南宁 莲花山组和那高岭组; i— 广西梧州苍梧组; j— 云南武定坡松冲组Fig.9 Palaeogeographic map of South China Block during the Early Devonian showing distribution of the Xujiachong vertebrate assemblage(modified from Gai et al., 2022) |
岩相古地理资料显示华南海可能是一个半封闭的陆表海湾, 并有1个间歇性向南的开口(Zhang et al., 2013; Xue et al., 2018), 因此, 鱼类、腕足动物和维管植物等在华南板块上的地域性分布可以被解释为封闭海效应(Gai et al., 2022), 生物在这个封闭的环境中独立演化, 表现出很高的多样性和很强的土著性。华南鱼属在广西南宁地区的新发现, 将该属的分布从华南海的西缘扩大至东缘, 同时将徐家冲鱼类组合的古地理分布扩大至云南武定地区(图9)。云南武定和广西南宁分别位于华南海的西缘和东缘, 华南鱼属同时在上述2个地区的发现表明盔甲鱼类在华南海这一独立的演化区域进行交流和扩散, 为支持华南板块早泥盆世古鱼类作为一个独立的动物区系提供了进一步的化石证据。同时, 广西南宁莲花山组与那高岭组过渡层中的徐家冲鱼类组合物种多样性最高, 几乎涵盖了与其他地区对应的所有属种, 说明广西南宁地区可能是盔甲鱼类辐射和扩散的中心之一。
1)系统描述了广西南宁下泥盆统布拉格阶那高岭组下部的盔甲鱼类新材料, 基于详细的形态学对比和讨论, 建立了华南鱼科2个新种— — 丘比特华南鱼(Huananaspis cupido sp. nov.)和梁氏亚洲鱼(Asiaspis liangi sp. nov.)。
2)华南鱼属在那高岭组的发现丰富了早泥盆世华南鱼类的多样性, 扩大了徐家冲鱼类组合的化石内容, 为云南武定坡松冲组与广西南宁那高岭组之间的对比提供了直接化石证据, 进而将该组合的古地理分布扩大至云南武定地区。
3)广西南宁莲花山组与那高岭组过渡层中的徐家冲组合物种多样性最高, 几乎涵盖了与其他地区对应的所有属种, 说明广西南宁地区可能是盔甲鱼类辐射和扩散的中心之一。
4)早泥盆世布拉格期的古鱼类组合、早期维管植物组合和腕足类动物组合表现出显著的地方性色彩, 可能跟华南海是一个半封闭的陆表海湾有关。由于封闭海效应, 生物在这个封闭的环境中独立演化, 表现出很高的多样性和很强的土著性, 最终在华南板块上形成一个独立的动物区系, 即“ 盔甲鱼— 云南鱼” 动物区系。
致谢 感谢广西南宁曾广春和梁江先生赠送华南鱼标本, 许勇、史爱娟、曾广春绘制文章插图, 高丽芳制作华南鱼模型, 北京大学薛进庄老师对地层古地理意义的讨论并提供古地理图模板。
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|