


{kind=link}
{kind=link}
{kind=link}
{kind=link}
古生代psammichnitids(砂迹类)的行为习性演化及其生物古地理迁移*
[张立军1  , 庞志超
, 庞志超1 , 杨琦琦1 , 樊柳1 , 魏凡2, 3 , 宋慧波1 , 牛永斌1 ]
, 庞志超|
|

第一作者简介 张立军,男,1982年生,副教授,主要从事遗迹学和沉积学的教学和研究工作。E-mail: Ljzhanghpu@gmail.com。
Psammichnites和 Dictyodora为古生代海相环境典型的弯曲带状遗迹,是由具有虹吸器官的生物所形成的遗迹化石,一般被归入到psammichnitids。通过文献资料调研和样品分析,对古生代 Psammichnites和 Dictyodora的形态功能对比和时空分布特征进行研究。结果表明: (1) Psammichnites通常具有短、浅的凹槽; 随着时间的推移, Dictyodora的壁高却逐渐增大。(2) Psammichnites主要分布于正常浅海环境; Dictyodora大都分布于深海环境,也有个别类型分布于浅海环境。(3) Psammichnites的种级多样性在石炭纪浅海环境呈快速增加,属于晚期辐射类型,但 Psammichnites gigas广泛分布于寒武纪早期,具有很好的地层指示意义; Dictyodora的种级多样性在奥陶纪快速增加,属于早期辐射类型。(4)寒武纪—奥陶纪, Psammichnites和 Dictyodora主要分布于南半球中高纬度的Iapetus和Rheic大洋,其中Laurussia大陆的形成以及Iapetus和Rheic大洋的闭合对其古地理分布具有显著影响; 自石炭纪开始, Psammichnites和 Dictyodora均向新的空白生态位进行迁移,反映出造迹生物对于新生态位的适应和迁移均受环境变迁的强烈影响; 二叠纪之后, Psammichnites和 Dictyodora在地层中没有发现,可能暗示它们的造迹生物均在二叠纪末生物大灭绝事件中遭受了毁灭性打击。上述对 Psammichnites和 Dictyodora行为习性演化特征及其生物古地理迁移特征的系统总结和归纳,可为从精细和精深角度深入解读遗迹化石所反映的遗迹类群的演化生态学和生物古地理迁移特征提供新资料和新认识。
About the first author ZHANG Lijun,born in 1982,is an associate professor of Henan Polytechnic University. He is mainly engaged in teaching and researches on ichnology and sedimentology. E-mail: Ljzhanghpu@gmail.com.
Psammichnites and Dictyodora are grouped into psammichnitids,representing the typical slug-like bulldozers with a snorkel living in the marine environments during the Palaeozoic. Through the morphological and functional analysis of Paleozoic Psammichnites and Dictyodora,combining with their spatio-temporal distribution characteristics,it is found: (1) Psammichnites generally has short and shallow grooves,and the wall height of Dictyodora gradually increased with time. (2) Psammichnites is mainly distributed in the shallow-marine environment,whereas Dictyodora is mainly distributed in deep-marine environment with occasional occurrences of certain ichnospecies in the shallow-marine environment. (3)The ichnodiversity of Psammichnites ichnospecies rapidly increased in the Carboniferous shallow-marine environment,belonging to the top-heavy pattern. Psammichnites gigas is distributed widely in the Early Cambrian strata,which is of biostratigraphic importance. The ichnodiversity of Dictyodora ichnospecies rapidly expanded in the Ordovician,representing the bottom-heavy pattern. (4) Psammichnites and Dictyodora were mainly distributed in the Iapetus and Rheic oceans in the middle and high latitudes of the southern hemisphere during the Cambrian to Ordovician. Due to the formation of Laurussia and the closure of Iapetus and Rheic oceans, Psammichnites and Dictyodora migrated to the new empty ecological niches from the Carboniferous to Permian,indicating that the adaptation and migration of trace makers to new ecological niches might be strongly influenced by environmental changes. Until now Psammichnites and Dictyodora have not been found in the post-Permian strata,possibly suggesting that their trace markers went extinct in the end-Permian mass extinction. On the basis of the above systematic summary and induction of the behavior habits,biogeographic migration characteristics and evolutionary characteristics of Psammochnites and Dictyodora,it can provide new data and new understanding of the evolutionary ecology and biogeographic migration characteristics of trace groups reflected by trace fossils from a detailed and profound perspective.
砂迹类遗迹化石(psammichnitids)泛指古生代海相环境中呈带状的履带式移迹, 其中间具有线形凹槽, 可能是软躯体造迹生物的虹吸器官在沉积物表面所留下的遗迹(Seilacher, 1997, 2007)。psammichnitids主要包括2个遗迹属, 即Psammichnites和Dictyodora (Seilacher-Drexler and Seilacher, 1999; Seilacher, 2007)。Psammichnites可能是一类具有虹吸器官的地下食泥动物的机会主义觅食策略形成的潜穴(Seilacher and Gá mez-Vintaned, 1995), 广泛分布于古生代滨浅海环境(Hä ntzschel, 1975; McIlroy and Heys, 1997; Má ngano et al., 2002, 2022; MacNaughton et al., 2021), 以寒武纪纽芬兰世最为发育; 依据形态特征的细微差别, 将寒武纪的Psammichnitites系统修订为Psammichnites gigas gigas, P.gigas circularis, P.gigas arcuatus 和 P.saltensis(Má ngano et al., 2022)。Dictyodora造迹生物是通过虹吸器官连接到水— 沉积物界面, 其蜿蜒曲折的形态被认为是进行复杂觅食策略的证据, 螺旋形或环状蜿蜒形态进一步表明造迹生物能够系统和有效地开发特定区域的食物资源(Benton, 1982; Baucon and Neto de Carvalho, 2008)。Dictyodora长期以来被认为是古生代重要的深海遗迹化石, 在欧洲和北美地区的奥陶纪— 密西西比亚纪尤为重要(Benton and Trewin, 1980; Pickerill et al., 1987; Pek and Zapletal, 1990; Orr et al., 1996; Chlupá č , 1997; Neto de Carvalho, 2001; Mikulá Š et al., 2002, 2004; Uchman, 2004; Uchman et al., 2005; Jensen et al., 2016)。在奥陶纪深海沉积物中, Dictyodora通常与雕画迹一同出现(Seilacher et al., 2005)。近年来, 在阿根廷中西部志留纪晚期和早— 中泥盆世的边缘海环境(Luca and Basilici, 2013; Pazos et al., 2015a)、中国西秦岭二叠纪乐平世陆架边缘斜坡环境(Zhang et al., 2020; 党志英和张立军, 2020a, 2020b)也发现了Dictyodora, 这些新发现对Dictyodora是否可以作为深海环境的指相化石提出了质疑(Pazos et al., 2015a)。
近几年, 笔者在华南寒武纪和秦岭二叠纪地层中发现了形态特征精美的Psammichnites和Dictyodora。文中通过对笔者发现的这些遗迹化石的形态功能分析, 结合构建的显生宙psammichnitids数据库(附表1), 系统总结和归纳了psammichnitids的行为习性演化及其生物古地理迁移特征, 这为从精细和精深角度深入解读遗迹化石所反映的遗迹类群的演化生态学和生物古地理迁移特征提供了新资料和新认识。
笔者采集的Psammichnites主要保存于云南晋宁二街寒武系红井哨组厚层细粒石英砂岩的上层面(图1-A, 1-B, 1-E)。红井哨组被认为形成于寒武纪第二世第三期(胡世学等, 2014), 代表三角洲前缘和近滨中部的沉积环境(罗惠麟等, 2008)。Psammichnites呈水平带状, 常弯曲或杂乱无序地相互交切(图1-A, 1-B), 潜穴内部具紧密排列的蹼状构造(图1-E), 中间为1条直或略弯的凹槽, 宽0.5~1.2 mm。移迹宽8~16 mm, 最长可达到140 cm。尚没有发现实体化石与Psammichnites共生。根据最新的Psammichnites分类学特征(Má ngano et al., 2022), 晋宁寒武系的Psammichnites应归属于Psammichnites gigas circularis。
 | 图1 古生代不同形态的Psammichnites A, B— 云南晋宁二街, 寒武系红井哨组, 厚层砂岩表面发育的Psammichnites gigas circularis, 黄色和白色箭头指示的是Psammichnites中的蹼状构造; C— 澳大利亚Carnarvon盆地, 瓜德鲁普统Kennedy群, Mungadan砂岩中的Psammichnites plummeri(引自Feng et al., 2021); D, E— 云南晋宁梅树村, 寒武系石岩头组, Psammchnites gigas circularis, E为垂直切面方向, 可见Psammichnites里面的蹼状构造Fig.1 Different morphology of Psammichnites in the Paleozoic |
Dictyodora主要来自于西秦岭甘南地区二叠系乐平统毛毛隆组灰色厚层细粒石英砂岩(图2)。甘南地区毛毛隆组发育大套巨厚层— 厚层砾屑石灰岩, 砾屑石灰岩中可见到大量宾夕法尼亚亚纪和乌拉尔世的腕足类、 
 | 图2 西秦岭甘南卡加曼乡乐平统毛毛隆组发育的Dictyodora A— 中层细粒石英砂岩上浮雕保存的Dictyodora cf. scotica; B— A的示意图, 展示了Dictyodora的详细形态特征, 包括基部潜穴(绿色箭头)和顶部壁结构(红色箭头); C— Dictyodora zimmermanni, 上浮雕保存; D— 凹形(白色箭头)和凸形(黄色箭头)呈下浮雕形式保存的基部潜穴, 后者具有微弱的蹼状构造(弯曲的虚线); E— D. zimmermanni的垂直截面图, 显示逐渐变细的壁结构; F, G— 中层细粒石英砂 岩上浮雕保存的D. zimmermanniFig.2 Dictyodora from the Lopingian Maomaolong Formation in Kajiaman, Gannan, west Qinling |
通过对大量文献资料调研和样品观察, 笔者总结和归纳了Psammichnites和Dictyodora的异同点(表1)。相同点是两者潜穴的表面均具有隆脊、凹槽或壁, 可能是由于造迹生物虹吸器官运移所致(图1-D; 图2-E)。不同之处表现为: (1)Psammichnites没有壁、有凹槽, 而Dictyodora具有壁, 可能是由于造迹生物虹吸器官的长短所致, 即Psammichnites具有较短的虹吸器官, 而Dictyodora具有较长的虹吸器官; (2)Psammichnites潜穴内部可见蹼状构造, 而Dictyodora仅在壁中可见蹼状构造。
| 表1 古生代psammichnitids(砂迹类)形态特征对比 Table1 Morphology comparison of the Paleozoic psammichnites |
通过对遗迹化石的形态功能进行解析, 可以清楚地归纳出Psammichnites和Dictyodora的造迹生物的行为习性特征。Psammichnites截面呈椭圆形, 顶部可见由短虹吸器官运移所形成的凹槽, 整体形态没有增大或加深, 而是保持一个相对稳定的状态, 说明该造迹生物并没有侵入沉积物内部, 而是通过类似推土机的方式进行平移清除前面沉积物, 并在身体末端进行回填, 从形成了潜穴内部的蹼状构造(图1)。Dictyodora的造迹生物使用觅食策略和Psammichnites的相同, 但虹吸器官随着时间演化而越来越长, 至石炭纪时Dictyodora壁上的蹼状构造宽度都小于1 mm, 成为最为显著的特征。而较长的虹吸器官可以允许造迹生物在深阶层的沉积物内部进行觅食, 因此Dictyodora并不是在沉积物— 水界面进行觅食的, 而是在沉积物内部进行觅食, 且Dictyodora的蛇曲形态越密集, 代表沉积物内部食物越多。
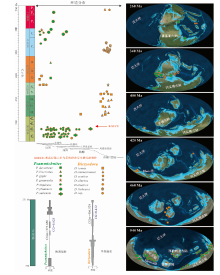
依据将psammichnitds投影到古地理、古环境图(图3)中而得到的时空分布特征为: (1)Psammichnites和Dictyodora均分布于寒武纪— 二叠纪, 在二叠纪之后的地层中尚没有相应的报道。(2)Psammichnites和Dictyodora均受岩相高度控制, 主要保存在砂— 泥岩互层界面附近。(3)Psammichnites在寒武纪早期(纽芬兰世至第二世)分布范围广泛、丰度很高、分异度很低(1个种), 至石炭纪边缘海环境中分异度增大, 但分布范围和丰度较低: 奥陶纪, Dictyodora在深水环境中快速辐射, 丰度和分异度都很高; 在志留纪晚期和泥盆纪初期, 浅海环境仍有发育; 石炭纪只有1个种D. libeana, 分布于深海环境; 二叠纪的Dictyodora仅在中国西秦岭地区有报道。(4)Psammichnites为晚期辐射类型, 说明在Psammichnites演化后期物种的多样性增多; 而Dictyodora为早期辐射的类型, 表明Dictyodora演化的前期物种的多样性快速增长(Zhang et al., 2022)。
 | 图3 古生代psammichnitids的时空分布和多样性分异(古大陆底图源自互联网https://deeptimemaps.com/)Fig.3 Spatio-temporal distribution and diversification of the Paleozoic psammichnitids (base map of palaeocontinent from https: //deeptimemaps.com/) |
欧洲和华南寒武纪的Psammichnites都呈蜿蜒曲折形的潜穴, 且一直延续到北美石炭纪和西澳二叠纪的浅海环境中(图4)。寒武纪早期, Psammichnites顶部的线形凹槽呈正弦曲线形, 而到石炭纪和二叠纪末, 线形凹槽变得平直、短, 说明造迹生物的虹吸器官— — 通气管变得更短, 且在牧食过程中造迹生物身体也不再摆动, 进而导致中部的凹槽变得更为平直。
 | 图4 古生代psammichnitids形态素描图(据Seilacher, 2007; 有修改)Fig.4 Sketch showing morphological changes of the Paleozoic psammichnitids (modified from Seilacher, 2007) |
奥陶纪至泥盆纪, Dictyodora大多在层面上呈现出蜿蜒曲折的觅食迹; 到了石炭纪和二叠纪, 锥形壁逐渐增高, 说明造迹生物的虹吸器官加长, 其可以在更深阶层进行觅食; 虹吸器官在沉积物中仍然具有开辟道路的能力, 进而在锥形壁上形成蹼纹构造(图4)。在垂直切面的锥形壁上可以看到纤细、倾斜的运移轨迹, 在水平面切上可以看到越向上锥形壁越小, 说明这是由于虹吸管落后于身体运动的结果。在奥陶纪— 泥盆纪, Dictyodora的形态特征整体上变化不大, 大致为简单的蜿蜒蛇曲形态, 显得更为粗放。但在密西西比亚纪, Dictyodora呈现出复杂的变化特征: 首先, 在几个相邻的裂片之后, 蛇曲脱离了起始的螺旋, 变成了类似于奥陶纪— 泥盆纪的Dictyodora的粗放特征(图4); 其次, 大多数的蛇曲形裂片均保持一定的间距, 形成更为紧密的螺旋形态(Seilacher, 2007)。密西西比亚纪Dictyodora的形态呈现出更为规则的螺旋形锥形壁, 揭示出该时期Dictyodora造迹生物可以更加规则地利用身体来获得最大回报的食物来源。
综上所述, 古生代不同时期Psammichnites和Dictyodora的形态特征具有明显的继承性, psammichnitids可能是单系遗迹类群, 即它们有共同的起源生物, 而不是因进化趋同所形成的(Zhang et al., 2022)。这样持续的缓慢形态变化特征, 可以解释为行为习性的进化特征受到一个相似的基因组控制的表达形式(Seilacher, 2007)。
Psammichnites gigas和P. saltensis主要分布于寒武纪纽芬兰世和第二世, 在苗岭世— 早泥盆世早期尚未见有Psammichnites的可靠种报道。泥盆纪, 浅海环境保存的Psammichnites devonicus和P. biseriatus仅有1个命名层位, 其他层位未有相应的报道。笔者认为, 寒武纪第二世和苗岭世之交的灭绝事件(ROECE)可能对早期快速辐射的Psammichnites造成了沉重打击, 但随着长期的缓慢进化和复苏, 在晚古生代浅海环境中仍然可以看到形态特征精美的Psammichnites grummula, P. implexus, P. plummeri, 不过仅分布于美洲和西欧地区。Psammichnites的古地理迁移与Dictyodora不同, 其并没有明显受到海盆扩张和古陆碰撞的影响。
Dictyodora主要产出于奥陶纪— 志留纪中期的深海浊积岩中, 具有D. tenuis、D. zimmermanni、D. scotica等多种形态(Benton and Trewin, 1980; Orr and Pickerill, 1995; Orr, 1996; Orr and Howe, 1999; Uchman et al., 2005)。与志留纪晚期— 中泥盆世三角洲系统中零星分布的Dictyodora相比, 石炭纪的D. liebeana与Cosmorhaphe、Paleodictyon、Nereites等典型复理石相遗迹化石共生, 且主要保存在深海浊积岩序列中(Lang et al., 1979; Benton, 1982; Stepanek and Geyer, 1989; Pek and Zapletal, 1990; Orr et al., 1996; Llompart and Wieczorek, 1997; Mikulá Š et al., 2004; Baucon and Neto de Carvalho, 2008; Luca and Basilici, 2013; Pazos et al., 2015b)。在西秦岭乐平统毛毛隆组浊流沉积的斜坡环境中, 也发现了形态丰富的Dictyodora。
从古生代psammichnitids的时空分布数据(图3)可知, psammichnitids主要产出于古生代的南半球。Psammichnites主要分布于纽芬兰统和第二统。Dictyodora的第1次辐射发生在早、中奥陶世, 范围是从浅海环境到深海环境。在奥陶纪生物多样性辐射期间, 海洋无脊椎动物多样性显著增加导致浅海环境生物生存竞争加剧(Servais et al., 2010; Servais and Harper, 2018), Dictyodora的造迹生物可能作为先驱者探索并迁移到了深海中空缺的生态位, 并最终在奥陶纪晚期和志留纪统治了Iapetus洋。
由于波罗地海和劳伦西亚大陆的碰撞, Iapetus洋在文洛克世晚期关闭(Torsvik and Cocks, 2016)。在此期间, Dictyodora可能迁移到新的生态位, 如位于Rheic洋西缘阿根廷的志留纪晚期— 早、中泥盆世的边缘海环境(Luca and Basilici, 2013; Pazos et al., 2015b; Domeier, 2016)和位于古特提斯洋的中国西秦岭志留纪晚期的半深海环境(杨式溥和胡云绪, 1992)。在晚泥盆世生物大灭绝期间, 生态危机对生物多样性的影响更为严重(Stigall, 2012; 戎嘉余和黄冰, 2014), 海洋陆架基质从泥盆纪以碎屑岩为主变为密西西比亚纪以碳酸盐岩为主, 这导致陆架CaCO3积累增加(Walker et al., 2002; Rong et al., 2007)。在密西西比亚纪, Rheic洋闭合(Domeier, 2016; Torsvik and Cocks, 2016), Dictyodora的造迹生物可能无法适应浅海环境中的生态变化, 从而退回到古特提斯洋的半深海环境, 且一直延续到二叠纪末期。此外, 也不排除Dictyodora产出自Gondwana周边地区的浅海环境, 然后再迁移到其他海洋(如Rheic洋、Iapetus洋、古特提斯洋), 可能是由于古生代发生了Rheic洋和Iapetus洋的闭合, 导致Dictyodora在这些地区均已灭绝, 但在古特提斯洋中, 其一直存活到二叠纪末期。Dictyodora的演化趋势表明, 造迹生物对新生态位的适应和迁移受到了环境变化的强烈影响。
1)通过对古生代psammichnitids的详细分析与解剖, 认为Psammichnites和Dictyodora是古生代海相浅水和深水环境中典型的蛇曲形潜穴, 一般是具有虹吸器官的一类生物所形成的遗迹化石。Psammichnites具有短的、浅的凹槽, Dictyodora的形态随时间推移壁高逐渐增大。Psammichnites种级多样性在石炭纪快速增加, 属于晚期辐射类型, Dictyodora种级多样性在奥陶纪快速增加, 属于早期辐射类型。
2)生物古地理迁移研究发现, Laurussia大陆的形成和Iapetus和Rheic大洋的闭合, 对古生代Psammichnites和Dictyodora的古地理分布具有显著影响。Psammichnites和Dictyodora自石炭纪开始向新的空白生态位迁移, 反映出造迹生物对于新生态位的适应和迁移受环境变迁的强烈影响。尚没有发现二叠纪之后的psammichnitids地层记录, 可能暗示了二叠纪末生物大灭绝事件对其造成沉重打击导致其灭绝。
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|