



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
扬子板块西北缘下泥盆统平驿铺组双壳类遗迹化石*
[张小乐1, 2  , 王怿
, 王怿1 , 刘建波3 , 黄璞1 , 徐洪河1 ]
, 王怿|
|

第一作者简介 张小乐,男,1986年生,中国科学院南京地质古生物研究所助理研究员,研究方向为沉积学、遗迹学。E-mail: xlzh@nigpas.ac.cn。
四川北部广元地区下泥盆统平驿铺组产出丰富的双壳类遗迹化石,但尚未开展过系统的遗迹学工作,也未进行过行为学与古生态学分析。文中针对四川广元马家剖面平驿铺组中部地层开展沉积学和系统遗迹学研究,在陆棚至近滨带下部沉积中共识别出3个属种的双壳类遗迹化石,包括 Lockeia siliquaria, Protovirgularia rugosa, Ptychoplasma vagans,以及其他无脊椎动物门类遗迹化石 Cruziana problematica, Dimorphichnus isp., Lophoctenium isp.等。对双壳类遗迹化石的出现方式、形态以及保存特征的分析表明,平驿铺组中部曾出现过2种类型的双壳类,分属具分叉足(原鳃目)和楔状足的类群。原鳃目分布于过渡带和陆棚区,营沉积物摄食生活; 受高能事件或沉积物掩埋影响,它们由停息状态变为“逃逸”状态的行为产生了 Pro. rugosa。具楔状足的双壳类依生态类型可分为2类: 在过渡带与陆棚区,漫游的、营沉积物摄食的类型产生了 Pty. vagans;而在近滨下部,滤食性双壳类居群则在不同期次高能事件的间隙对沉积物进行大规模殖居,产生了 L. siliquaria。平驿铺组中部遗迹群落面貌的垂向变化,反映了随着浅海环境水动力增强,发育在具有一定硬度基底的 Cruziana遗迹相被 Skolithos遗迹相逐渐替代的过程。本研究强调,通过对双壳类遗迹化石的出现形式、保存和形态的精细观察,可以对造迹生物类群、古生态学和行为习性学特征等进行可靠的解释。
About the first author ZHANG Xiaole,born in 1986,is an assistant research fellow in Nanjing Institute of Geology and Palaeontology,Chinese Academy of Sciences. He is engaged in sedimentology and ichnology. E-mail: xlzh@nigpas.ac.cn.
Abundant bivalve trace fossils are found in the Lower Devonian Pingyipu Formation in Guangyuan area,northern Sichuan Province,but systematic ichnological work and behavioral and paleoecological analysis have not been carried out. Close observations on the occurrence,preservation style and morphological features of bivalvian traces provide invaluable information to guide the interpretation of their trace maker,ethological and paleoecological features. Here we carried out sedimentary and systematic ichnology studies on the middle part of the Lower Devonian Pingyipu Formation at the Majia section,Guangyuan area. The results suggest occurrence of bivalvian trace fossils,including Lockeia siliquaria, Protovirgularia rugosa, Ptychoplasma vagans as well as trace fossils of other invertebrate groups such as Cruziana problematica, Dimorphichnus isp.,and Lophoctenium isp. in the lower shoreface to off shore settings. The overall morphological and taphonomic features of the trace fossils suggest that,two major groups,including the cleft-footed(protobranchia)and wedge-footed bivalves occurred in the middle part of the Pingyipu Formation. The protobranchia were deposit-feeding bivalves distributed in the transitional zone and off shore settings. The transition from resting to “escape”state of these bivalves,probably due to erosive events or a higher sedimentation rates,produced the trace fossil Pro. rugosa. The wedge-footed bivalves could be further divided into two groups. The vagile,deposit-feeding type occupied the transitional zone and off shore settings. Their crawling behavior,probably linked to an effort to search for edible material from the sediment generated the trace fossil Pty. vagans. While in the lower shoreface setting,colonies of suspension-feeding bivalves colonized the sediment during relatively tranquil intervals between erosional events. The vertical transition of trace fossil composition in the middle Pingyipu Formation suggests a substitution of Cruziana ichnofacies developed on a relative stiff substrate by the latter Skolithos ichnofacies in an upward shallowing succession.
双壳类是现代海洋中数量最多、种类最丰富的无脊椎动物类群之一(Stanley, 1970; Pohlo, 1982), 分布范围跨越了从浅海到深海的广阔区间, 表现出极高的生态多样性和行为多样性(Bromley, 1996; Hanken et al., 2001; Gingras et al., 2008; Zonneveld and Gingras, 2013)。相较于其他种群, 双壳类对水动力、氧气含量、盐度等环境因素有着更宽的适应范围, 这使得它们可以在生存压力较大的环境中存活并发展出个体数量庞大的种群(Heckle, 1972; Miller, 1984; Gingras et al., 2008)。
古代沉积物中保存了大量双壳类遗迹化石。其中, 以反映双壳类静止状态的LockeiaJames, 1879(Seilacher, 1953; Osgood, 1970; Nara, 2003)以及反映双壳类侧向运动的PtychoplasmaFenton and Fenton, 1937和 ProtovirgulariaM'Coy, 1850(Seilacher and Seilacher, 1994; Uchman et al., 2011)的时空分布最为广泛。这些遗迹化石不仅蕴含着解读古代双壳类行为学、古生态学特征的关键信息, 也为古环境恢复与重建提供了重要依据(Rindsberg, 1994; Seilacher and Seilacher, 1994; Má ngano et al., 1998; Ekdale and Bromley, 2001; Bromley et al., 2003; Uchman et al., 2011; Luo et al., 2017)。
四川北部广元地区下泥盆统平驿铺组产出丰富的双壳类遗迹化石。前人(如Zhang et al., 2020)曾对其进行了初步报道, 但尚未开展过系统的遗迹学工作, 也未进行过行为学与古生态学分析。文中聚焦四川广元马家剖面下泥盆统平驿铺组中部, 通过开展系统的沉积学研究, 构建沉积框架; 重点关注其中的双壳类遗迹化石, 开展系统遗迹学研究, 对相关遗迹属种分类学特征进行回顾与梳理。
基于遗迹化石形态、保存与出现形式的详细观察, 结合近年来对现生和古代不同类群双壳生态行为学特征的新认识, 讨论造迹双壳类所属类群、行为学及古生态学特征, 并分析其古环境意义。
研究区位于扬子板块西北缘(图1-A), 属于早古生代龙门山盆地的一部分(侯鸿飞等, 1988; 鲜思远等, 1995)。下泥盆统海相碎屑沉积在该区出露广泛, 以平驿铺组为代表(鲜思远等, 1995; Ma et al., 2009)。前人在平驿铺组中报道了丰富的无脊椎动物化石, 包括介形类、腹足类、双壳类和腕足类等(林文球等, 1986; Ma et al., 2009)。植物化石如Zosterophyllum sp.和Taeniocrada sp.以及鱼类化石Sanqiaspis sichuanensis, Sichuanthus sp. 和Polybranchias sp.的共同出现, 表明该组的地质时代为早泥盆世洛赫考夫期— 布拉格期(Edwards et al., 2016)。系统的沉积学研究指出, 平驿铺组的沉积特征总体反映了三角洲— 滨岸环境(侯鸿飞等, 1988; 郑荣才等, 1997; Ma et al., 2009)。该组产丰富的遗迹化石, 前人针对这些遗迹化石的分类、古生态以及沉积环境重建等开展了一系列研究(林文球等, 1986; 张立军和龚一鸣, 2013; Zhang, 2014; Zhang and Zhao, 2015)。
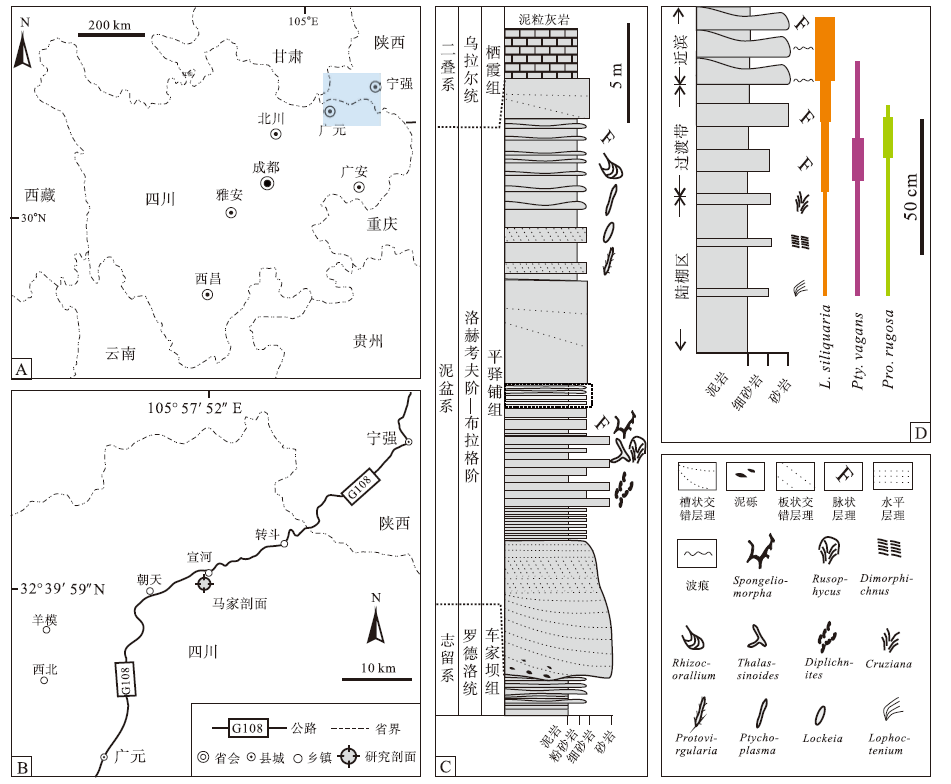 | 图1 四川广元马家剖面地理位置、交通图以及下泥盆统平驿铺组沉积序列特征 A— 研究区(蓝色方框)的地理位置; B— 马家剖面的交通图; C— 岩性柱状图, 示平驿铺组主要沉积岩石特征(据Zhang et al., 2020; 有修改); D— 平驿铺组中部(C中红色曲线方框)沉积相划分及所产出的遗迹化石种类与分布, 不同颜色线段的粗细代表各属种双壳类遗迹化石丰度Fig.1 Location maps and sedimentary sequence features of the Lower Devonian Pingyipu Formation of Majia section in Guangyuan area, Sichuan Province |
本文研究材料来自广元市宣河镇的马家剖面(32° 39'59″ N, 105° 57'52″ E)(图1-B)。下泥盆统平驿铺组以砂岩、粉砂岩与泥岩为特征, 与上覆二叠系乌拉尔统栖霞组中— 厚层泥粒灰岩以及下伏志留系罗德洛统车家坝组薄层粉砂岩和泥岩均呈平行不整合接触(图1-C)。
遗迹化石产自平驿铺组中部一套120 cm厚的地层中(图1-D), 该套地层以泥岩和细砂岩为特征, 向上砂岩层数及厚度均逐渐增加(图2-A)。底部为薄层泥岩夹少量薄层细砂岩, 产少量Lockeia siliquaria, Protovirgularia rugosa以及Ptychoplasma vagans, 伴生的遗迹化石包括Cruziana problematica, Dimorphichnus isp.以及Lophoctenium isp.。中部为薄层泥岩与砂岩互层, 砂岩中发育波状层理, 双壳类遗迹化石增多, 包括L. siliquaria, 丰富的Pty. vagans以及Pro. rugosa。上部为纵向叠覆的薄层透镜状砂岩, 夹少量泥岩, 发育波状层理和波痕(图2-B), Pty. vagans以及Pro. rugosa逐渐减少直至消失, L. siliquaria占主导并持续增多, 最终在近顶部形成了这一单一属种密集出现的局面。该序列被一套厚约5 m的中— 厚层砂体覆盖, 后者由透镜状砂岩纵向叠覆形成(图2-C), 发育槽状交错层理(图2-D)、板状交错层理(图2-E), 未发现遗迹化石。
 | 图2 四川广元马家剖面平驿铺组中部研究层段野外露头沉积序列与沉积构造特征 A— 研究层段以薄层砂岩夹薄层泥岩或薄层砂岩与薄层泥岩互层为特征, 向上砂岩层数及厚度逐渐增加, 地质锤长度约27 cm; B— 纵向叠覆的薄层砂岩呈透镜状, 并发育波状层理和波痕; C— 研究层段被中— 厚层砂岩覆盖, 白色虚线示两者的分界线, 注意该套砂体由透镜状砂岩纵向叠覆构成; D— 中— 厚层砂岩发育槽状交错层理; E— 中— 厚层砂岩中发育板状交错层理Fig.2 Outcrops showing sedimentary sequence and sedimentary features of studied succession from the middle part of Pingyipu Formation at Majia section in Guangyuan area, Sichuan Province |
研究序列上覆砂体由透镜状砂岩纵向叠覆形成, 发育板状交错层理与槽状交错层理, 表明这套沉积最可能为浅海沙坝沉积(Almar et al., 2010; Price et al., 2014)。序列顶部为纵向叠覆的薄层透镜状砂岩, 发育波状层理和波痕, 这些较高能沉积构造的出现及其与上覆沙坝沉积的密切关系表明它为近滨底部沉积(Reineck and Singh, 1980; Walker and Plint, 1992)。序列中部由发育波状层理的薄层细砂岩与泥岩组成, 分别对应有波浪牵引的较高能环境和水体较安静的环境, 支持将其划分为近滨向陆棚的过渡区(Reineck and Singh, 1980; Schrö der et al., 2011)。序列底部以泥岩为主, 除产出双壳类遗迹化石之外还发现有三叶虫遗迹Cruziana problematica, Dimorphichnus isp.(Seilacher, 2007)和Lophoctenium isp.。Ekdale 和 Bromley(2001)曾报道Lophoctenium与Protovirgularia和Lockeia构成复合遗迹化石, 表明其部分与底栖双壳类有关。然而, 文中Lophoctenium未表现出与其他双壳类遗迹的明显联系, 因而暂被解释为环节类遗迹(Fu, 1991)。上述岩石特征及Cruziana遗迹相分子的出现, 表明序列底部为滨外陆棚沉积(Walker and Plint, 1992)。
LockeiaJames, 1879
Lockeia siliquariaJames, 1879
图3-A至3-D
 | 图3 四川广元马家剖面平驿铺组中部Lockeia siliquaria, Ptychoplasma vagans及伴生遗迹化石(下浮雕正凸起) A— L. siliquaria以单一属种的方式大量产于薄层砂岩底板, 注意纵向叠覆的砂岩每层底部均有产出, 镜头盖直径7.5 cm; B— A的局部放大, 示L. siliquaria典型的杏仁状外形; C— 细砂岩产大量L. siliquaria, 较多Pty. vagans (Pty., 下同)以及少量Pro. rugosa (Pro., 下同), 箭头示与之紧密相连的类似Lockeia的杏仁体), 部分L. siliquaria中部发育平行于长轴的脊, 示Pty. vagans由一系列类似L. siliquaria、较为连续的杏仁状凸起构成; D— 薄层细砂底部L. siliquaria与Lophoctenium isp.(Loph.)共同产出, 但二者并无明显联系, 注意L. siliquaria (Lo.) 向一侧倾斜; E— 薄层细砂岩底部Pty. vagans与Cruziana problematica (Cru.)共同出现Fig.3 Lockeia siliquaria, Ptychoplasma vagans and accompanying trace fossils(convex hyporelief)from the middle part of Pingyipu Formation at Majia section in Guangyuan area, Sichuan Province |
鉴定特征: 表面光滑的杏仁— 椭圆体, 一端或两端收窄, 中间沿主轴有时具凸起的脊。以下浮雕正凸起形式产于岩层底面。
描述: 杏仁状或椭圆状凸起, 一端或两端变窄(图3-A, 3-B, 3-C)。以正凸起形式产于砂岩底面, 充填物与上覆砂岩一致。在近滨底部纵向叠覆的薄层砂岩中, 每一层底部均以高密度、单一属种的形式产出(图3-A), 但即使是在分布特别密集的区域(> 40个/100 cm2), 样本之间的交切也不常见; 样本排列没有方向性, 少数样本形状不规则, 表面光滑无纹饰; 长度8~25 mm, 宽度3~8.7 mm, 统计分析表明其长度分布呈明显正态分布(图4-A), 且长与宽分布呈明显的正相关(图4-B); 较大的样本高度约为7 mm, 但通常高度在2~4 mm之间。在过渡带与陆棚区, 与Pty. vagans, Pro. rugosa以及Lophoctenium isp.伴生, 并常与前两者构成复合遗迹化石, 长度3~20 mm, 宽度1~7.7 mm; 一些样本发育有沿长轴分布的脊(图3-C), 另一些样本有时向一侧倾斜(图3-D)。
 | 图4 四川广元马家剖面平驿铺组中部Lockeia siliquaria长度分布特征(A)及其长度与宽度分布的相关性(B)Fig.4 Statistic analysis showing length distribution(A)and relationship between the length and width(B) of Lockeia siliquaria from the middle part of Pingyipu Formation at Majia section in Guangyuan, Sichuan Province |
比较与讨论: 前人在Lockeia下建立了数个遗迹种, 包括L. siliquariaJames, 1879、L. amygdaloidesSeilacher, 1953、L. ornateBandel, 1967、L. czarnockiiKaraszewski, 1975、L. elongateYang, 1984、L. avalonensisFillion and Pickerill, 1990、L. serialisSeilacher and Seilacher, 1994、L. triangulichnusKim, 1994、L. cordataRingsberg, 1994、L. hunanensisZhang and Wang, 1996以及L. cunctatorSchlirf et al., 2001。Schlirf 等(2001)根据德国南部法兰克尼亚地区上三叠统的材料认为, L. amygdaloides, L. avalonensis, L. triangulichnus均是L. siliquaria的晚出异名; 而L. elongate和L. serialis因没有指定模式标本并对其描述, 属于无效命名。另外, L. hunanensis的照片与特征描述不清, 有效性存疑(Má ngano et al., 2002)。综上所述, 目前遗迹属Lockeia下共有5个有效遗迹种:L. siliquaria是最常见的遗迹种, 通常为表面光滑的杏仁— 椭圆体; L. ornatus以表面具同心状纹饰为特征, 在原产地常常与Protovirgularia rugosa构成复合遗迹化石; L. czarnockii以其较大的体型以及水滴状的形态区别于其他遗迹种; L. cunctator由分叉的棒状— 杏仁状凸起排布成列或梅花状; L. cordata则以表面光滑的箭头或心形形态为特征。文中材料的形态特征与L. siliquaria高度吻合, 支持将其划归于这一遗迹种。
PtychoplasmaFenton and Fenton, 1937
Ptychoplasma vagans Ksi
图3-C, 3-E; 图5-A, 5-B, 5-E
 | 图5 四川广元马家剖面平驿铺组中部Ptychoplasma vagans与Protovirgularia rugosa及伴生遗迹化石(下浮雕正凸起) A— 砂岩底板发育丰富的Pty. vagans 与Pro. Rugosa, 箭头示大型Lockeia isp.(Lo.); B— A中虚线方框的放大, 注意Pro. rugosa两侧的V字形脊及其向左逐渐朝上穿越岩层的趋势, 箭头示类似Lockeia的杏仁体; C— 细砂岩底部Pro. rugosa与类似Lockeia (箭头)的杏仁状凸起相连, 注意其中部明显的隆起以及V字形脊的开口背离Lockeia; D— 细砂岩底板Pro. rugosa上部细小的管状物(白色箭头), 可能为造迹生物虹吸管的遗迹, 其弯折处向一侧倾斜, 并只在一侧发育脊; E— 细砂岩保存较长的Pro. rugosa与Pty. Vagans; F— Protovirgulariaisp.与Dimorphichnus isp.(Dim.)共同出现Fig.5 Ptychoplasma vagans, Protovirgularia rugosa and accompanying trace fossils(convex hyporelief)from the middle part of Pingyipu Formation at Majia section in Guangyuan area, Sichuan Province |
鉴定特征: 由一系列近似连续的、横切面为龙骨状、外观为杏仁状的凸起组成的蜿蜒或环状的脊状物, 通常保存于岩层底面。
描述: 遗迹由一系列类似于Lockeia的凸起组成(图3-C)。相邻凸起或紧密连接在一起, 或有一定间距。最高处位于凸起中心, 向两端变低。遗迹呈线型, 长70~120 mm, 宽5~8 mm, 凸起高度为3~4 mm。沿层面展布, 横切面垂直于层面(图5-A, 5-B, 5-E), 或者朝转角内侧倾斜(图3-E)。
比较与讨论: Seilacher和Seilacher(1994)将由一系列杏仁状凸起组成的遗迹化石命名为Lockeia serialis。但是由于未指定正模并缺乏照片和描述, Schlirf等(2001)认为其无效。Uchman等(2011)将L. serialis划归到Ptychoplasma。根据他们的回顾与厘定, Ptychoplasma包含3个有效遗迹种, 并可通过凸起的形态和连续性加以区分:模式种Pty. excelsumFenton and Fenton, 1937由连续的杏仁体按叠瓦状形式排列构成; Pty. vagansKsiazkiewicz, 1977由近似连续的一系列杏仁体组成; Pty. conica Pień kowski and Uchman, 2009由不太连续的圆丘状凸起组成。Ptychoplasma偶尔也会出现V字形脊, 但频率和规整性远低于Protovirgularia (Uchman et al., 2011)。文中的材料由一系列近似连续的杏仁状凸起构成, 支持将其划归到Pty. vagans。
ProtovirgulariaM'Coy, 1850
Protovirgularia rugosaMiller and Dyer, 1878
图3-c; 图5
鉴定特征: 通常为保存在岩层底面的凸起, 由中部隆起向两侧延伸出V字形脊。长度较短, 与Lockeia isp.紧密相连。
描述: 水平— 倾斜的遗迹化石, 穿越层面(图5-A, 5-B, 5-C)。外形特征在层面上体现最明显, 呈S型蜿蜒(图5-B, 5-C, 5-D), 以下浮雕正凸起形式保存于砂岩底部。沿遗迹走向发育中间隆起, 并向两侧延伸出清晰的V字形脊(图5)。长度变化较大, 较短标本长为30~40 mm, 其他一些长度则达到200 mm。宽度为5~15 mm。相邻V字形脊间距变化较大, 范围在0.5~8 mm之间, 与主轴的夹角通常为60° ~90° 。一些标本在弯折处呈现出不对称性, 只发育V字形脊的其中一臂(图5-D)。多数样本与Lockeia siliquaria紧密相连, 且V字形脊的开口方向多与之背离(图5-B, 5-C)。
比较与讨论: 前人在Protovirgularia下建立了不同遗迹种。Han和Pickerill(1994)对该属进行了厘定, 认为模式种P. dichotoma是唯一有效的遗迹种。但经过Seilacher和Seilacher(1994)、Uchman(1998)、Má ngano等(2002)的系统工作, 一般认为该遗迹属下包括6个有效种(Luo and Shi, 2017, 分别为Pro. dichotomaMcCoy, 1850、Pro. pennatusEichwald, 1860、Pro. tuberculateWilliamson, 1887、Pro. rugosaMiller and Dyer, 1878、Pro. longespicataDe Stefani, 1885以及Pro. bidirectionalisMá ngano et al., 2002。Pro. dichotoma是一个龙骨状的遗迹化石, 有明显的中隆, 从其中部向两侧伸出V字形脊; Pro. pennatus具有排列紧密但细小的V字形脊; Pro. tuberculate中部具明显的沟槽而呈现出双叶状, 其V字形脊上具间距规则的小瘤; Pro. rugosa是较短的Protovirgularia遗迹化石, 一端与Lockeia相连; Pro. longespica个体较大, 其V字形脊呈乳头状; Pro. bidirectionalis是一个较浅的、具厚的衬里、整体呈U型的潜穴, 两端V字形脊以相反的方向排列。文中的材料大多为较短的遗迹化石, 以一定角度与层面交叉, 具中间隆起, 两侧为V字形脊, 且末端常与Lockeia相连。这些特征支持将其划归为Pro. rugosa。
得益于系统性观察, 目前对现生双壳不同类群的生态与行为学特征已经有了较深入了解。研究发现, 双壳类的爬行、挖掘等行为主要通过发达的足来完成。依照足的形态, 可划分为两大类群: 占双壳类大部分的具有楔状足的类群, 以及具有分裂足的原鳃目(Yonge, 1939; Stanley, 1970; Seilacher and Seilacher, 1994)。在挖掘与爬行过程中, 这两大类群使用了相似的策略, 均包括足的前伸与扩张形成锚、相应肌肉收缩拖拽壳体整体前进这一基本过程(Yonge, 1939; Rhoads, 1963; Stanley, 1970)。伴随这一过程的反复出现, 双壳类能够以不同角度穿过砂— 泥沉积物界面(Má ngano et al., 1998)。在爬行过程中, 其壳体底部与足在沉积物底面形成了具有船型横切面的隆起。另外, 当双壳类处于静止状态时, 足的扩张形态可以在沉积物中记录下来, 形成杏仁状的凸起(Seilacher, 1953; Osgood, 1970; Seilacher and Seilacher, 1994)。有时两瓣壳缝合处的压痕也可能保存下来, 形成沿杏仁体长轴发育的脊(Má ngano et al., 1998)。
依据上述回顾可知, 平驿铺组中所产丰富的Protovirgularia, Ptychoplasma与Lockeia均为双壳类遗迹(图6)。其中L. siliquaria记录了处于静止状态的双壳类足的扩张形态, 或者指示壳在沉积物中的印痕(图6-B)。它们的造迹生物可能是具楔状足的双壳类, 也可能是原鳃目(Seilacher and Seilacher, 1994)。例如在过渡带和陆棚沉积中, L. siliquaria与Pty. vagans和Pro. rugosa共同出现或构成复合遗迹化石, 就是对这种情况的反映。Pty. vagans被解释为具楔状足双壳类的爬行遗迹(图6-C): 它们起伏的外观反映了双壳类爬行过程中的韵律式运动; 其船型的横切面对应双壳类在爬行过程中, 壳体底部与足在沉积物中形成的凹陷(Crimes et al., 1981; Kim, 1994; Rindsberg, 1994; Pień kowski and Uchman, 2009; Uchman et al., 2011); 而足伸缩方向的不同, 可能导致遗迹化石局部形态特征的差异, 例如倾斜或不对称性等的出现(Radley et al., 1998)。虽然该遗迹属有时也出现类似V字形脊的构造, 但它们可能是因壳体的外扩, 而不是足的张开产生的(Uchman et al., 2011)。Pro. rugosa则代表了原鳃目双壳类的爬行遗迹(Má ngano et al., 1998; Ekdale and Bromley, 2001; Ferná ndez et al., 2010; Nara and Ikari, 2011; Luo and Shi, 2017)(图6-D), 其隆起两侧特征性的V字形脊反映了爬行过程中, 原鳃目足前端2片巨大裂片张开后形成的锚的印痕(Seilacher and Seilacher, 1994)。
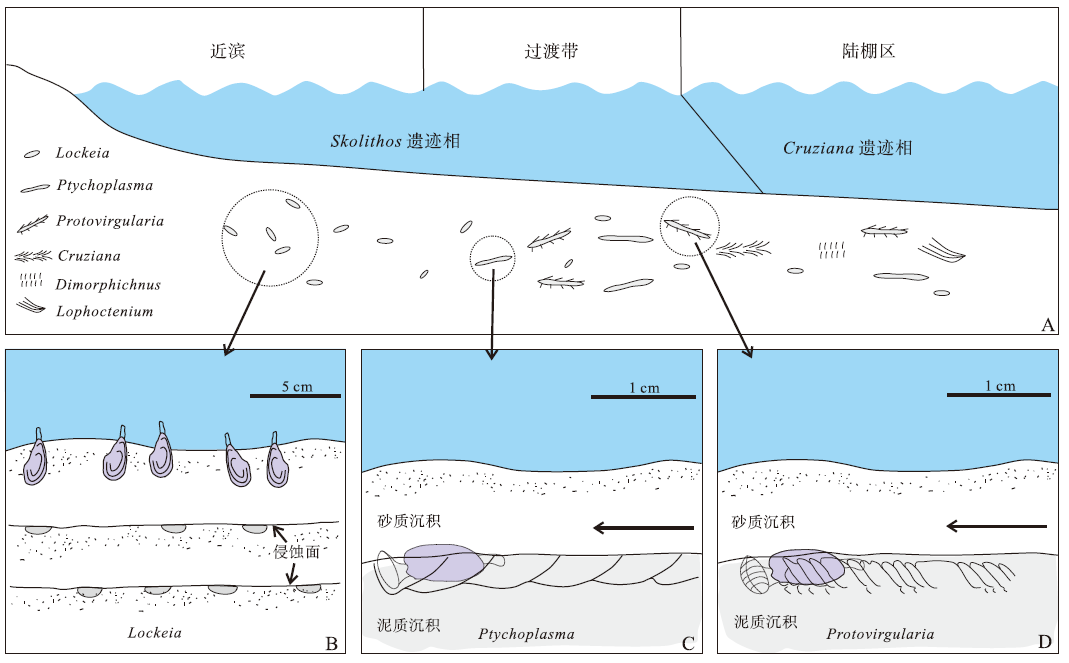 | 图6 四川广元马家剖面平驿铺组中部遗迹化石分布形式及不同类群双壳类遗迹化石形成机理 A— 平驿铺组中部遗迹化石分布形式, 箭头示移动方向; B— 具楔状足的滤食性双壳类居群在不同期次的高能事件的间隙对沉积物进行大规模定居, 在近滨纵向叠覆砂岩底部每一层均形成Lockeia; C— 具楔状足双壳类爬行过程形成Ptychoplasma示意图, 箭头指示移动方向; D— 原鳃目的分裂足在爬行过程中形成Protovirgularia示意图(修改自Nara and Ikari, 2011)Fig.6 Distribution pattern of ichnofossils and formation mechanism of bivalvian trace fossils from the middle part of Pingyipu Formation at Majia section in Guangyuan area, Sichuan Province |
以往的研究表明, 通过对Lockeia, Protovirgularia和Ptychoplasma遗迹化石形态、保存与出现形式的精细观察, 可以获得丰富的行为学与古生态学信息(Rindsberg, 1994; Seilacher and Seilacher, 1994; Má ngano et al., 1998; Ekdale and Bromley, 2001; Nara, 2003; Uchman et al., 2011)。
平驿铺组中Pty. vagans被解释为具楔状足双壳类的爬行遗迹。它们倾向于与Cruziana以及Lophoctenium等典型觅食构造出现于近滨带下部与过渡带沉积。这些特征很可能表明Pty. vagans代表了漫游的、营沉积物摄食双壳类的爬行行为。这一推断被Pty. vagans的外形所支持, 其蜿蜒或环状的形态可能正是双壳类爬行以搜索食物的反映(Rindsberg, 1994; Uchman et al., 2011)。
平驿铺组中P. rugosa被解释为原鳃目的爬行遗迹。其保存特征表明它是一个较浅阶层的遗迹。现生原鳃目多生活在沉积物较浅层, 它们是高度漫游的生物, 利用须状的触手从沉积物中分选食物(Yonge, 1939; Bender and Davis, 1984)。因此, Pro. rugosa也极有可能是由一类漫游的、营沉积物摄食的原鳃目双壳所形成。值得注意的是, 平驿铺组中Pro. rugosa大多长度较短, 以一定角度与岩层界面交切, 表明大多情况下这些双壳类只沿着层面移动了很短的距离, 即向上穿越过层面。另外, 作为一个复合遗迹化石, 在很多标本中可观察到其V字形脊的开口方向背离Lockeia。鉴于V字形开口方向指示双壳类的前进方向(Seilacher and Seilacher, 1994), 因此, 上述现象可能代表了原鳃目由浅海高能事件(例如风暴)(Seilacher and Seilacher, 1994)或较高的沉积速率(Má ngano et al., 2002)所触发的由停息状态转变为“ 逃逸” 状态的行为。
平驿铺组中L. siliquaria遗迹化石以2种方式出现。其中第1种产于过渡带与陆棚环境, 常伴随Protovirgularia和Ptychoplasma, 或构成这些遗迹化石的一部分。鉴于Protovirgularia和Ptychoplasma一般被解释为爬行遗迹, 因此这些L. siliquaria可能代表了漫游的、营沉积物摄食双壳类的暂时停息行为(Seilacher, 1953; Hakes, 1976)。第2种是在近滨下部以单一属种的形式大量出现的类型, 其沉积和保存特征表明, 这些L. siliquaria可能是经历了高能事件的剥蚀之后保存下来的, 因而它们原本可能是相对较深的潜穴, 反映了双壳类的居住行为(Má ngano et al., 2002; Paranjape et al., 2013)。这些遗迹化石之间少有交错, 个体长度分布呈正态分布, 且长度与宽度的分布正相关性明显, 因而可能反映了一类个体数量庞大的、具楔状足的滤食性双壳类居群的出现(Má ngano et al., 1998)。它们的分布没有定向性, 或许是对不定向水流的响应。L. siliquaria在纵向叠覆的薄层砂岩每一层底部均大量出现, 可能反映了数次双壳类大量定居于海底, 而后被高能事件打断, 待水动力减小后, 再一次进行定居这一复杂过程(Osgood, 1970; Má ngano et al., 1998)(图6-B)。
如前文所述, 平驿铺组中部遗迹化石的分布有明显的规律性, 这反映了浅海环境下随着水动力条件的增加, 发育在具有一定硬度基底的Cruziana遗迹相逐渐被Skolithos遗迹相替代的过程。在陆棚区, 漫游的、营沉积物摄食的双壳类, 包括原鳃目与具楔状足的类型产生了少量Lockeia siliquaria, Ptychoplasma vagans以及Protovirgularia rugosa。值得注意的是, 不少Pro. rugosa的V字形脊轮廓清晰, 且相邻的脊排列密集, 反映了原鳃目在较为坚硬的基质中努力以小幅度方式前进的现象(Má ngano et al., 1998; Paranjape et al., 2013)。这些双壳类遗迹化石与少量Cruziana problematica, Dimorphichnus isp.以及Lophoctenium isp.共同出现, 代表了发育在具有一定硬度基底而非典型软基底的Cruziana遗迹相的出现(图6-A)。在过渡带, 双壳类遗迹化石丰度逐渐增高, 这有可能是因为在砂— 泥韵律式沉积中这些遗迹化石更易保存, 且后期岩石沿层面裂开后更容易被观察到。与此同时, 其他门类的遗迹化石基本消失不见, 或许是因为相对于其他类群, 双壳类对动荡环境具更高承受能力(Heckle, 1972; Miller, 1984)。在近滨带下部, 遗迹化石类型发生了明显的变化, 表现为漫游的、营沉积物摄食的双壳类遗迹, 如Pro. rugosa和Pty. vagans均消失不见, 沉积物被个体数量庞大的、具楔状足的滤食性双壳类居群定居, 表明随着水动力增强, Cruziana遗迹相彻底被Skolithos遗迹相取代。
1)在四川北部马家剖面平驿铺组中部沉积中识别出3个属种的双壳类遗迹化石, 分别为Lockeia siliquaria, Protovirgularia rugosa与 Ptychoplasma vagans。Pty. vagans是营沉积物摄食的、具楔状足双壳类的爬行遗迹。Pro. rugosa是营沉积物摄食的原鳃目的爬行遗迹, 其形态和保存特征表明它可能代表了浅海环境下原鳃目受高能事件或较高沉积速率影响后, 由停息状态变为“ 逃逸” 状态的行为。在过渡带与陆棚区, L. siliquaria代表具楔状足和原鳃目双壳类的停息遗迹, 而在近滨带下部, 密集分布的L. siliquaria反映了滤食性的、具楔状足双壳类居群的多次定居事件。
2)四川北部马家剖面平驿铺组中部双壳类及其他门类遗迹化石的垂向分布形式, 反映了浅海环境下随着水动力条件的增强, 发育在具有一定硬度基底的Cruziana遗迹相逐渐被Skolithos遗迹相替代的过程。
致谢 野外过程中得到中国科学院南京地质古生物研究所张红勇大力协助; 3位评审老师提出的宝贵意见和建议对本文的提升效果显著, 在此表示诚挚感谢!
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|