


{kind=link}
{kind=link}
{kind=link}
古海洋氧化还原条件的遗迹化石定量表征特征:以华南二叠纪末生物大灭绝事件为例*
[丁奕1, 2  , 张立军
, 张立军3 ]
, 张立军]
|
|

第一作者简介 丁奕,男,1989年生,讲师,主要从事沉积学和遗迹学的教学和研究工作。E-mail: dingyi@nwu.edu.cn。
地质历史时期的重大生物—环境事件往往伴随着古海洋海水氧化还原条件的改变,而遗迹化石作为原位保存的生物成因沉积构造,对于解读古海洋氧化还原条件具有显著的优势。通过对前人常用的遗迹学参数进行分析总结,发现遗迹化石多样性、生物扰动强度、潜穴直径、特征遗迹化石组合这4项定量参数可以表征古海洋氧化还原条件的变化。文中以华南二叠系乐平统遗迹化石及生物扰动构造作为研究对象,系统分析了二叠纪末生物大灭绝事件前后遗迹化石参数表征的古海洋氧化还原条件变化特征:自吴家坪期晚期华南古海洋开始出现缺氧,然而该缺氧状态在长兴期不具有持续性,而是呈现出周期性缺氧/贫氧→富氧/有氧的波动特征;在二叠纪末生物大灭绝之前,煤山剖面高精度的定量遗迹学参数指示长兴组24e层顶部存在缺氧事件,并与大灭绝事件有着良好的对应关系。这一实例具体展示了遗迹学参数在古海洋水体氧化还原条件重建中应用前景广阔。
About the first author DING Yi,born in 1989,is a lecturer of Weinan Normal University. He is mainly engaged in teaching and researches of sedimentology and ichnology. E-mail: dingyi@nwu.edu.cn.
Major biotic and environmental events in Earth history are often accompanied by palaeo-ocean redox condition changes,as biogenetic sedimentary structures in situ,trace fossils play a significant role in reconstructing palaeo-ocean redox conditions. This study analyzes and summarizes the previous ichnological parameters,and suggests the ichnodiversity,bioturbation degree,burrow size,and key ichnotaxa can be quantitatively used to indicate the palaeo-ocean redox conditions. Additionally,Lopingian trace fossils and bioturbation structures in South China are systematically analysed in order to reconstruct the redox conditions during the end-Permian mass extinction. The quantitative ichnological parameters reveal that the ocean anoxic event occurs in late Wuchiapingian,and the ocean anoxia is not permanent in Changhsingian,instead,obvious periodic fluctuations between anoxic/dysoxic and oxic can be observed. The high-resolution quantitative ichnological proxies at Meishan section suggest that ocean anoxia occurs in the top of Bed 24e in the Changhsing Formation,appears to correlate with the biotic mass extinction. The above examples show that the application of ichnological parameters in the reconstruction of palaeo-ocean redox conditions has broad prospects.
遗迹化石是地史时期生物在沉积物或其他底质(如生物壳体)内部或表面遗留下的各种生命活动的记录(杨式溥等, 2004)。作为地质历史时期生物与环境相互作用的重要媒介, 遗迹化石在早期生命起源探索、后生动物演化、古生态与古环境重建等研究中发挥了至关重要的作用(龚一鸣等, 2009; 罗茂等, 2021)。地质历史转折时期重大生物演化事件往往伴随着海洋水体氧化还原状况的改变(Song et al., 2017; Li et al., 2020; Zhang et al., 2022), 古海洋氧化还原条件的研究已成为近年来地球科学的热点与前沿(Li et al., 2010; Wang et al., 2018; Fang et al., 2021)。目前, 判断古海洋水体氧化还原条件的参数指标有很多, 总体上可分为3类(表1)。 第1类是岩石矿物指标, 如水平纹层发育的黑色富有机质岩石常被认为形成于缺氧环境(Wignall, 1994), 黄铁矿形态、大小和矿化程度也可作为水体氧含量的重要标志(Bond and Wignall, 2010), 后者已成为海洋缺氧事件研究的重要手段(如Huang et al., 2019; Zhang et al., 2020; Emmings et al., 2022), 但利用黄铁矿重建区域氧化还原环境通常存在局限性与不确定性, 需结合其他参数进行综合判断(Algeo et al., 2011; Kershaw et al., 2018)。第2类是地球化学指标, 因为氧化还原条件的改变会造成元素行为的变化, 从而留下丰富的地球化学记录。常规氧化还原地球化学指标包括Mo、U、V、Ce等微量元素(如Arnold et al., 2004; Kipp and Tissot, 2022), S、C、N等同位素(如Zhang et al., 2014; He et al., 2020), 以及姥鲛烷植烷比(Pr/Ph)、藿烷混合比(Tm/Ts)等生物标志化合物(如Grice et al., 2005; Cao et al., 2009); 然而元素行为受到多种因素影响, 因此利用地球化学指标反演水体氧化还原环境亦存在多解性, 可能会显示失真信号(张明亮等, 2017; Jin et al., 2018)。第3类是古生物学指标, 因为生物对环境氧含量的变化十分敏感(Allison et al., 1995; Semenza, 2007), 生物种类、形态、食性等均会随着氧化还原条件的变化发生相应改变, 因此化石能够作为指示水体氧含量变化的良好指标(如Lethiers and Whatley, 1994; Forel et al., 2009; Liu et al., 2010)。
| 表1 常见海洋水体氧化还原条件指示标志(修改自张明亮等, 2017; 樊秋爽等, 2022) Table1 Common parameters used for indicating ocean redoxconditions(modified from Zhang et al., 2017; Fan et al., 2022) |
遗迹化石数量多、分布广, 保存了造迹生物一系列生命活动的重要信息, 且形成后难以搬运改造, 是特定环境下生物活动的直接证据, 因此遗迹化石是指示水体氧化还原条件的良好标志(杨式溥等, 2004)。大量研究表明, 遗迹化石多样性、生物扰动强度、潜穴大小、阶层及特征遗迹化石组合的出现, 都受到氧化还原条件的制约(Savrda and Bottjer, 1986, 1989; Ekdale and Mason, 1988)。近年来, 古氧相研究中各类遗迹学参数越来越受到重视, 已被广泛应用于指示古海洋水体氧化还原条件(如Rodrí guez-Tovar and Uchman, 2010; Ferná ndez-Martí nez et al., 2021; Rodrí guez-Tovar, 2021)。文中聚焦遗迹化石在古海洋氧化还原条件研究中取得的新进展与新发现, 对各类遗迹学参数进行了系统梳理、讨论与总结, 并以二叠纪末生物大灭绝事件为例, 具体展示了遗迹学参数在古海洋水体氧化还原条件重建中的应用。同时, 得益于数学理论及计算机技术的引入, 新世纪以来, 遗迹学研究逐渐从定性向定量发展, 文中最后对未来遗迹学古氧相的定量研究进行了展望, 期望更多的遗迹学定量指标能够应用于氧化还原条件的研究中。
遗迹多样性主要用以描述遗迹属或种的数量或分异度(Seilacher, 1974; Uchman, 2004), 很早就被用于指示水体氧化还原条件(Savrda and Bottjer, 1989)。富氧环境适宜造迹生物生存与活动, 从而能够形成种类丰富的遗迹化石; 当水体处于贫氧环境时, 造迹生物因难以适应氧含量下降而大量死亡, 遗迹多样性随之显著降低; 当水体氧含量进一步减少, 处于缺氧环境时, 造迹生物几乎无法生存, 难以形成遗迹(表2)。
| 表2 重建海洋水体氧化还原条件的遗迹学指标(修改自Savrda and Bottjer, 1989; Zhang et al., 2018; Zheng et al., 2022) Table2 Ichnological parameters used for reconstruction of ocean redox environment(modified from Savrda and Bottjer, 1989; Zhang et al., 2018; Zheng et al., 2022) |
遗迹多样性与造迹生物多样性之间并无明确对应关系, 同一造迹生物可以形成不同遗迹, 不同造迹生物也能形成相似遗迹, 所以遗迹多样性并不等于生物多样性。遗迹化石分布也受到水动力条件的制约(Seilacher, 1967), 滨外与浊流环境中遗迹多样性往往较高, 高能的滨海环境(如潮上带)虽然水体氧含量充足, 但水动力与底质并不稳定, 遗迹多样性相对较低(Buatois and Má ngano, 2013)。因此, 在采用该参数指示氧化还原条件时, 应确保沉积相一致, 以减少水动力条件对结果造成的误差。
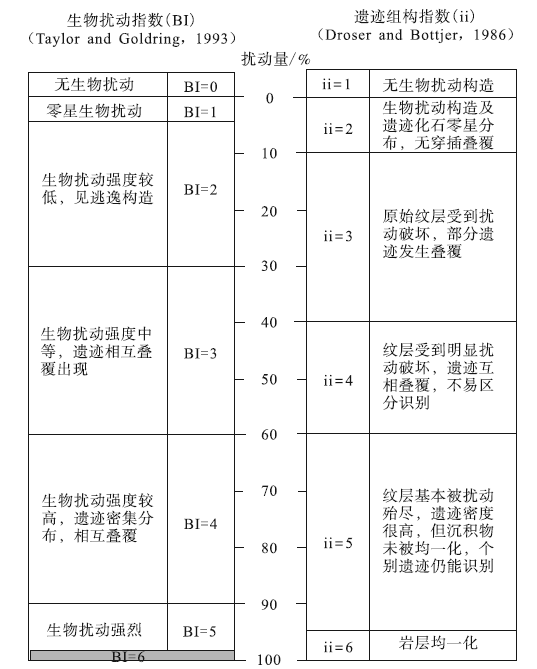
生物扰动指造迹生物对周围沉积物进行混合、破坏、搅动等改造作用形成的一系列构造(Meysman et al., 2006; Kristensen et al., 2012)。生物扰动强度与水体氧化还原条件之间具有良好对应关系(Savrda and Bottjer, 1986, 1989, 1991):富氧环境中, 造迹生物活跃, 生物扰动通常强烈复杂; 随着水体氧含量减少, 生物扰动强度相应减小; 缺氧环境下, 生物扰动极为微弱甚至完全停止(表2)。目前主要采用生物扰动指数(BI, Bioturbation index)与遗迹组构指数(ii, Ichnofabric index)2个参数对生物扰动强度进行定量表示(表3)。
| 表3 生物扰动指数(BI)与遗迹组构指数(ii)比较(修改自Luo et al., 2020) Table3 Comparison between bioturbation index(BI)and ichnofabric index(ii)(modified from Luo et al., 2020) |
生物扰动指数与遗迹组构指数的划分主要依靠人为判断, 易产生主观偏差, 因此, 基于网格法的定量统计工作已逐渐应用于生物扰动强度的研究中, 即随机选取沉积物一定区域, 设定适当网格比例, 统计有扰动构造网格所占百分含量, 从而精确统计生物扰动强度。该方法操作简单、统计准确, 已在遗迹学研究中取得了良好效果(Heard and Pickering, 2008; Marenco and Bottjer, 2010)。
体型大小是生物最重要的特征之一, 能为生物生理、行为、生态等研究提供重要证据(Cotgreave, 1993; 黄冰, 2008)。近年来, 化石体型所蕴含的生物学信息愈发受到关注, 为古环境与古生态重建提供了重要证据(如: Song et al., 2011; Chu et al., 2015)。生物体型与水体氧化还原条件之间关系紧密, 生物体型会随着水体氧含量的降低而减小(Rhoads and Morse, 1971)。遗迹化石虽非生物个体直接石化形成, 但作为生物行为习性的直接反映, 遗迹化石大小和造迹生物体型具有良好的相关性(Twitchett and Barras, 2004; Schlacher et al., 2016), 因此, 潜穴大小能够作为水体氧含量的可靠指标(表2)。水体处于富氧环境时, 最大潜穴直径(MBD, maximum burrow diameter)通常大于10 mm; 随着水体氧含量降低, 潜穴也随之减小; 当水体处于缺氧环境时, 最大潜穴直径通常小于5 mm(Savrda et al., 1984; Smith et al., 2000)。
利用潜穴大小指示水体氧化还原条件时, 应选取同一遗迹属种进行比较, 避免不同遗迹属种差异带来的误差。同时, 底质性质会影响遗迹的保存, 使原始潜穴的大小发生改变, 从而影响准确性, 同时在研究时, 也应尽量消除后期改造对真实结果产生的干扰。
一些特征遗迹化石的分布受到氧化还原条件的制约, 常被作为水体氧含量的重要指示标志(表2)。大部分甲壳类对水体氧化还原条件十分敏感, 一旦氧含量减少便难以生存(Harper et al., 1991; Song et al., 2014)。甲壳类通常难以形成实体化石, 但其产生的遗迹(如: Rhizocorallium、Thalassinoides)却能够保存下来, 作为富氧环境的典型标志(Savrda and Bottjer, 1986; Watkins and Coorough, 1997; Knaust et al., 2012)。然而, Rhizocorallium和Thalassinoides的造迹生物并非只有甲壳类, 多毛类也能形成类似潜穴(Knaust, 2013), 但由多毛类形成的Rhizocorallium和Thalassinoides并无明显古氧相指示意义, 因此在采用Rhizocorallium和Thalassinoides分析水体氧化还原条件时, 需确定其造迹生物类型。若Rhizocorallium潜穴壁发育微弱抓痕或Thalassinoides分枝处发生膨大, 则可判断其造迹生物为甲壳类(Luo et al., 2020), 可作为富氧环境的标志。
Chondrites是一类具树枝状分枝系统的特殊潜穴, 发育于显生宙各类沉积环境(Kim and Pickerill, 2003; Mekki et al., 2019)。Chondrites常形成于造迹活动的最后阶段, 且当水体氧化还原状态发生改变时, 其总是最早出现或最后消失, 因此常被作为指示低氧环境的典型标志(Bromley and Ekdale, 1984)。然而, Chondrites可以和其他遗迹共生于富氧环境中(龚一鸣, 2004; Baucon et al., 2020), 因此只有当直径小于1 mm且遗迹多样性很低时, Chondrites才能作为低氧环境的标志(晋慧娟等, 2003; 马会珍等, 2010), 直径大于1 mm、个体粗大的Chondrites并不能作为缺氧或贫氧环境的标志。
Zoophycos也被认为是一类能指示水体氧含量的特殊遗迹。Zoophycos具蹼状构造, 广泛分布于显生宙海相沉积中(Zhang, 2014; Zhang et al., 2015a, 2015b; Zhang and Zhao, 2016), 常见于氧化还原界面附近(张立军和龚一鸣, 2011), 被视为贫氧环境的典型标志(Ekdale and Lewis, 1991)。晋慧娟等(2003)认为, 在纵切面上呈新月型展布的Zoophycos才可作为贫氧环境的标志, 而发育在水平层面、呈喇叭形或螺旋形的Zoophycos并不能代表贫氧环境。Zoophycos蹼层中充填物颜色也受到水体氧含量影响, 郑锋利等(2022)对华北太原组不同环境中的Zoophycos进行了对比, 发现水体氧含量减少时, Zoophycos蹼层中充填物颜色会逐渐由灰白色变为黑色。以上研究成果为综合分析Zoophycos的古氧相意义提供了重要借鉴, 并再次印证特征遗迹化石虽与水体氧化还原条件存在一定联系, 但仍需结合多种因素综合分析, 才能得出更可靠的结论。
遗迹化石在沉积界面之下的垂直分布, 称为遗迹阶层或梯序(Ausich and Bottjer, 1982)。遗迹阶层分布受到水体氧化还原条件的影响(表2), 随着沉积界面之下孔隙水含氧量的减少, 较深阶层的遗迹逐渐增多, 最终完全取代浅阶层遗迹(Bromley and Ekdale, 1984; Savrda and Bottjer, 1986, 1989)。在很多遗迹学研究中, 通常采用最大下潜深度(maximum penetration depth)作为度量阶层的参数, 但最大下潜深度并不能完全反映遗迹阶层的变化, 应尽量选用平均下潜深度(mean penetration depth)作为反应阶层变化的指标(Luo et al., 2020), 通常平均下潜深度随着水体氧含量减少而逐渐减小。阶层并非只受水体氧含量的单一控制, 沉积物性质、捕食压力均会对遗迹的垂直分布产生影响(Bottjer and Ausich, 1986), 因此利用阶层重建水体氧化还原状态时, 也应充分考虑其他控制因素的影响。对遗迹阶层进行研究时, 应分别对浅阶层、中阶层及深阶层遗迹进行系统分析, 从而准确还原遗迹阶层所蕴含的古环境信息。
二叠纪末生物大灭绝事件(EPME, end-Permian mass extinction)是显生宙规模最大的生物灭绝事件, 约81%的海洋生物灭绝(Stanley, 2016; Fan et al., 2020), 持续了近2亿年之久的“ 古生代演化动物群” 被“ 现代演化动物群” 取代(Sepkoski et al., 1981; 戎嘉余和黄冰, 2014)。二叠纪末生物大灭绝研究吸引了众多学者的关注, 经过近30年多学科交叉研究与综合论证, 对这场地史时期最大生物灾难的严酷性已有了基本共识, 但对生物灭绝的诱因与机制依然存在争议。全球范围大量二叠纪— 三叠纪之交地层的沉积学和地球化学研究工作证实了二叠纪末海洋水体缺氧事件的普遍存在(如Huang et al., 2017; Liao et al., 2020; Xiang et al., 2020, 2022), 说明缺氧事件对二叠纪末海洋生态系统的全面崩溃产生了影响, 因此, 二叠纪末海洋缺氧事件目前已成为多学科领域共同深入研究的重大地学问题。前人选取了沉积相、地球化学、实体化石等诸多指标对二叠纪— 三叠纪之交海洋水体氧化还原条件进行了重建(如 Li et al., 2016; Fang et al., 2021; Newby et al., 2021), 但由于多解性与不确定性的限制, 造成了对该时期缺氧事件的认识并不统一。近10年来, 二叠纪末海洋缺氧事件的遗迹学研究受到了越来越多的重视, 中国遗迹学工作者更是在此方向取得了令人振奋的成果。
二叠纪末海洋缺氧事件研究中, 对海洋水体缺氧发生的时限(尤其是起始时间)争议较大。Isozaki(1997)认为, 二叠纪末海洋缺氧事件是一次持续时间超过20 Ma的超级缺氧事件, 从二叠纪吴家坪期初期海洋水体氧含量便开始减少, 直至三叠纪安尼期才恢复正常。但数值海洋模型显示海洋难以维持如此之久的分层状态(Hotinski et al., 2001), 同时后续古生代晚期大气氧含量数据也不支持超级缺氧事件的存在(Sperling et al., 2015), 因此缺氧事件的时限或许并没有20 Ma之久, 海洋缺氧事件可能发生在长兴期, 长达2 Ma的海洋缺氧是造成这次重大生物灾难的主要原因(Cao et al., 2009; Clarkson et al., 2016; Takahashi et al., 2021)。然而, 该观点依然存在诸多争议, 有学者认为二叠纪末海洋并非长期处于缺氧状态, 在大灭绝事件发生之前, 水体氧含量才发生明显下降(Brennecka et al., 2011; Fang et al., 2021; Pimentel-Galvan et al., 2022)。
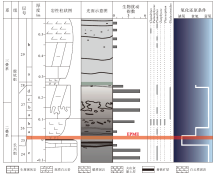
中国华南乐平统遗迹化石和生物扰动发育, 为重建晚二叠世海洋水体氧化还原条件提供了不可多得的遗迹学证据(图1)。广西来宾蓬莱滩北岸剖面乐平统发育连续, 遗迹丰富(图2), Ding等(2016)对该剖面吴家坪组和大隆组中生物扰动强度及遗迹化石分布进行了系统研究, 发现生物扰动强度在吴家坪组顶部明显降低(ii=1~2), 仅发育Zoophycos及小型Chondrites, 代表了典型的贫氧环境(图1-a)。四川广元上寺剖面乐平统遗迹发育, Zheng等(2022)对该剖面吴家坪组与大隆组中各项遗迹学参数进行了全面统计, 发现吴家坪期晚期地层中生物扰动强度显著下降(图1-b), 遗迹组构指数由4~6下降至1, 最大潜穴直径也由11~23 mm减至不足5 mm, 仅小型Planolites零星分布, 代表水体氧含量发生明显下降。由此可见, 二叠纪晚期海洋缺氧事件发生的时间远比生物灭绝事件要早, 在吴家坪期晚期, 华南地区海洋水体就已存在缺氧状态。
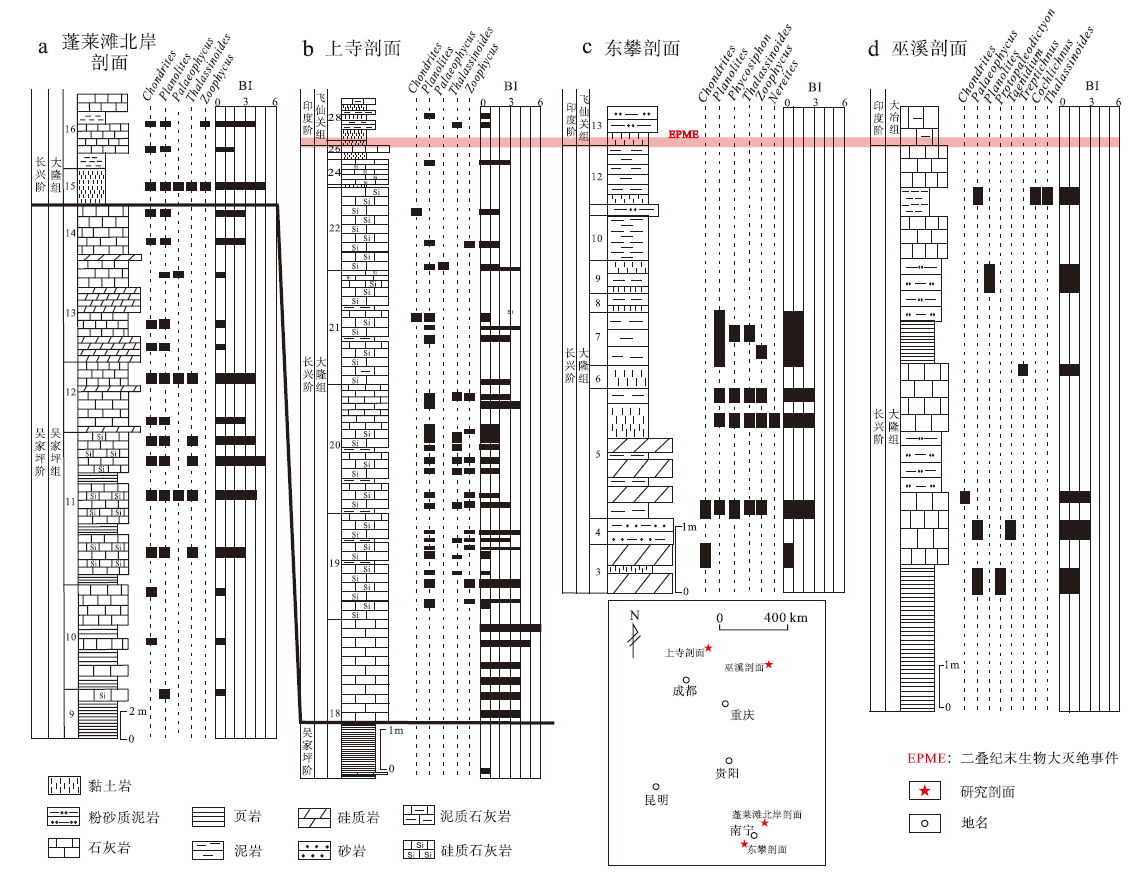 | 图1 华南二叠纪晚期遗迹化石分布及生物扰动变化 a— 广西蓬莱滩北岸剖面吴家坪组和大隆组遗迹化石分布及生物扰动指数变化(修改自Ding et al., 2016); b— 四川上寺剖面大隆组遗迹化石分布及生物扰动指数变化(修改自Zhang et al., 2018, 2020; Zheng et al., 2022); c— 广西东攀剖面大隆组遗迹化石分布及生物扰动指数变化(修改自Zhang et al., 2018); d— 四川巫溪剖面大隆组遗迹化石分布及生物扰动指数变化(修改自张静, 2015)Fig.1 Distribution of trace fossils and bioturbation index during the Late Permian in South China |
 | 图2 广西来宾蓬莱滩北岸剖面露头及典型遗迹化石(修改自Ding et al., 2016, 2021) a— 吴家坪组地层露头; b— 大隆组地层露头; c, d— 吴家坪组中丛藻迹Chondrites(Ch); e, f— 大隆组中海生迹Thalassinoides(Th); g— 大隆组中动藻迹Zoophycos(Zo); h— 吴家坪组中漫游迹Planolites(Pl)Fig.2 Outcrops of northern Penglaitan section in Laibin of Guangxi, and typical trace fossils from the studied section(modified from Ding et al., 2016, 2021) |
虽然吴家坪期晚期海洋水体氧含量就出现明显下降, 但遗迹学证据清晰表明, 二叠纪晚期海洋并非一直处于缺氧状态。Zheng等(2022)识别出上寺剖面大隆组遗迹组构指数与最大潜穴直径存在明显波动(图1-b), 在大隆组某些岩层中, 遗迹组合以小型Chondrites、Planolites和Zoophycos为主, 生物扰动强度较低(ii=1~2), 最大潜穴直径不足5 mm, 反映海洋处于缺氧或贫氧状态; 但大隆组中也同样发育了遗迹组构指数较高(ii=4~5)的岩层, 潜穴最大直径5~15 mm, Rhizocorallium和Thalassinoides发育, 代表典型的富氧环境。综上, 上寺剖面二叠纪长兴期海洋水体氧含量存在明显波动, 并非一直处于缺氧状态。这一发现也与Zhang等(2018, 2020)基于上寺剖面生物扰动指数、遗迹化石分布及黄铁矿粒径等重建出的二叠纪末周期性海水氧化还原波动结论一致。除上寺剖面外, Zhang等(2018)对广西东攀剖面大隆组上部各遗迹学参数进行了系统研究, 发现该剖面也记录了相似的氧含量波动, 虽然大隆组的生物扰动整体强度较低(BI=0~1), 遗迹化石罕见, 指示水体氧含量处于较低水平, 但该剖面依然存在生物扰动较强(BI=3)、遗迹化石多样性较高的岩层(大隆组第5层), 反映水体氧含量显著增加(图1-c)。张静(2015)基于四川巫溪剖面大隆组的遗迹化石多样性、生物扰动强度等遗迹学参数(图1-d), 对海洋氧化还原状态进行了重建, 结果也显示长兴期海水存在明显氧含量波动, 在大隆组中上部岩层中, 生物扰动从0激增至50%以上, 潜穴最大直径可达20 mm, 反映海洋水体从缺氧状态迅速变为富氧状态。
虽然二叠纪晚期华南地区从吴家坪期晚期开始海洋水体氧含量就发生明显下降, 但并非一直处于低氧状态, 依然存在氧含量正常时期, 水体氧含量存在明显波动, 这一发现也与前人通过地球化学指标重建的水体氧化还原状态(Xiang et al., 2016; Lei et al., 2017)相一致。由此可知, 长期缺氧或许并非造成二叠纪末海洋生物灭绝的主要原因, 造成水体氧含量波动的原因目前依然没有定论, 可能与米兰科维奇旋回或事件沉积(如风暴沉积、浊流沉积)相关(Ding et al., 2016; Zhang et al., 2018, 2020; Zheng et al., 2022)。
二叠纪末生物大灭绝的发生机制与诱因一直未有定论, 海洋缺氧是否是真凶依然成谜。相关争议很大程度上受限于氧化还原参数的多解性, 以及缺氧事件在不同地区的时序关系对比上存在困难(Cao and Zheng, 2009)。浙江煤山剖面是三叠系底界的全球界线层型剖面, 具高精度生物地层资料(Yin et al., 2001; Yuan et al., 2014), 遗迹化石及生物扰动非常发育, 为二叠纪末生物灭绝与缺氧事件的耦合研究补充了宝贵的遗迹学证据。笔者对煤山剖面二叠纪— 三叠纪之交的遗迹学研究进行了总结, 统计分析了该剖面24e层至29层的遗迹化石分布及生物扰动强度(图3), 并基于遗迹学参数, 恢复了二叠纪— 三叠纪之交水体氧化还原状态, 探讨缺氧事件与灭绝事件的相关性。
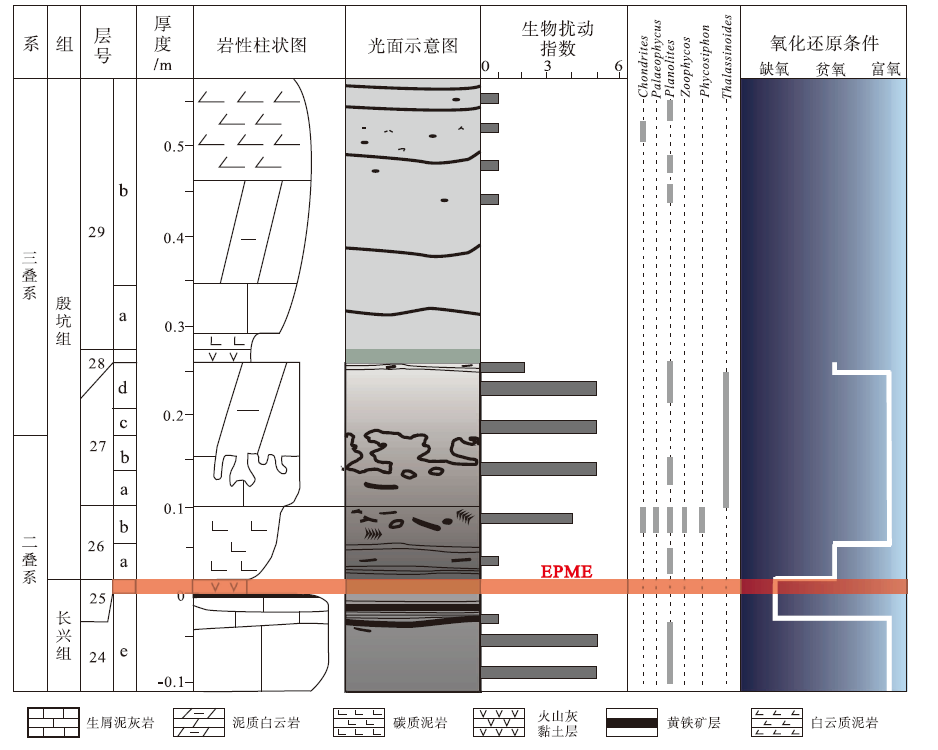 | 图3 煤山剖面二叠纪— 三叠纪之交岩性柱状图及遗迹学参数变化(数据来源Cao and Zheng, 2009; Zhao and Tong, 2010; Zheng et al., 2013; 丁奕, 2016)Fig.3 Lithostratigraphy and ichnological paremeter changes across the Permian-Triassic boundary at the Meishan section(data from Cao and Zheng, 2009; Zhao and Tong, 2010; Zheng et al., 2013; Ding, 2016) |
煤山剖面24e层为厚10 cm的深灰色生物碎屑泥灰岩, 下部生物扰动强烈(BI=5~6), 见大型Planolites, 指示海水氧含量正常, 而顶部有机质纹层出现, 生物扰动微弱(BI=0~1), 未见遗迹化石, 反映水体氧含量明显下降。25层为2 cm厚灰白色黏土层, 未见遗迹化石及生物扰动构造, 因此该层沉积时间短, 遗迹学指标无法反应水体氧化还原状态。26层可分为2段: 下部26a层为6 cm厚的黑色碳质泥岩, 纹层发育, 生物扰动微弱(BI=1), 小型Planolites零星分布, 反映水体处于贫氧状态; 上部26b层为5 cm厚的灰黑色碳质泥岩, 原始纹层被强烈扰动破坏(BI=4~5), 发育Chondrites、Palaeophycus、Phycosiphon、Planolites及Zoophycos, 沉积学和遗迹学参数均指示海水氧含量明显升高。27层依据岩性自下而上分为4段: 27a层为3 cm厚的灰色含生物碎屑泥灰岩, 生物扰动强烈(BI=5~6), 遗迹丰富, 以Planolites与Thalassinoides为主; 27b层为3 cm厚的深灰色生物碎屑泥质石灰岩, 扰动强烈(BI=5~6), 见大型Thalassinoides; 27c层为3 cm厚的灰色泥质白云岩, 扰动强烈(BI=5~6), 遗迹化石组合与27b层相似, 以大型Thalassinoides及Planolites为主; 27d层与27c层类似, 为约8 cm厚的含生物碎屑泥质白云岩, 生物扰动强烈(BI=5~6), 距该层顶部3 cm处, 出现水平纹层, 仅发育小型Planolites, 生物扰动指数下降(BI=2~3), 至顶部岩性过渡为黑色碳质泥岩, 未发育遗迹及扰动构造; 综上, 27层总体生物扰动强烈, 指示当时海洋水体处于氧含量正常状态, 至顶部生物扰动强度明显下降, 水体氧含量显著降低。28层与25层类似, 为不足1 cm厚的灰绿色黏土层, 缺乏遗迹学参数。29层可分为2段: 下部29a层为7 cm厚的灰色泥灰岩, 原始暗色纹层完好, 无遗迹及生物扰动构造; 上部29b层为22 cm厚的灰色泥质白云岩, 向上逐渐过渡为白云质泥岩, 生物扰动微弱(BI=1), 偶见Planolites。虽然29层对应的生物属种显著减少, 生物扰动强度与遗迹多样性降低, 但这并不能指示水体氧化还原状态的变化, 而是对应了生态环境的急剧改变。
总之, 煤山剖面二叠纪— 三叠纪之交的水体氧含量发生过明显波动, 并非一直处于缺氧状态, 进一步证实长期缺氧并非二叠纪末生物大灭绝的主要原因。煤山剖面遗迹学参数指示二叠纪末海洋水体在24e层顶部开始变为缺氧状态, 其恰好发生在二叠纪末大灭绝事件前极短时间内, 与生物大灭绝存在较好的对应关系。
随着科学技术的不断发展, 水体氧化还原条件的遗迹学角度研究必将从定性向定量发展, 数据分析、图像处理等技术为遗迹学定量研究注入了新的活力。Dorador和Rodrí guez-Tovar(2014)开发的IDIAP(Ichnological Digital Images Analysis Package)扩展包可在Adobe Photoshop 软件内对遗迹图像进行处理分析, 统计生物扰动构造百分含量, 并将此扩展包应用于IODP339航次的岩心分析中, 详细统计了各类遗迹化石及扰动构造的百分含量。Kikuchi等(2018)利用MIDIRT(Measurement of Ichnofossil Diversity by Image-Resampling Technique)算法对剖面上的遗迹多样性进行定量分析, 以减少露头地层出露不完全对统计造成的误差。综上, 有关遗迹学的定量学研究发展迅速, 各项定量指标的研究手段层出不穷, 未来必会有更多遗迹学定量参数成为重建水体氧化还原状态的重要指标。
笔者通过分析总结前人在氧化还原研究中所采用的各类遗迹学参数与指标, 认为遗迹化石多样性、生物扰动强度、潜穴大小、特征遗迹化石组合可以作为反映水体氧化还原条件的良好指标。当氧含量正常时, 遗迹化石多样性丰富, 生物扰动强烈, 潜穴直径大于10 mm, Thalassinoides、Rhizocorallium等遗迹化石出现; 当水体处于贫氧状态时, 遗迹化石多样性显著减少, 生物扰动微弱, 潜穴直径5~10 mm, 仅小型Zoophycos、Chondrites零星出现; 当水体完全缺氧时, 遗迹化石及生物扰动构造完全消失。但水体氧含量并非唯一影响遗迹化石分布的环境因素, 水动力条件、营养供给、底质性质等因素都会造成遗迹学参数的改变, 此时遗迹化石多样性降低、生物扰动减弱、潜穴减小反映的其实是水体环境整体的恶化。因此, 遗迹学参数能够作为水体氧含量正常环境的准确指标, 但在指示缺氧或贫氧环境时, 单独的遗迹学参数并不能准确反映水体氧化还原条件的改变。利用遗迹学参数重建水体氧化还原状态时, 需综合判断其他因素的影响, 方能得出可靠结论。遗迹学指标目前已成为水体氧化还原条件研究的重要指征参数, 且随着各种定量方法的引入, 遗迹学在未来古氧相的研究中必将拥有更为广阔的前景。
致谢 审稿专家对本文提出宝贵意见, 在此致以衷心感谢!
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|