


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
豫西济源地区中下三叠统遗迹网络分析*
[许欣1  , 邢智峰
, 邢智峰1, 2 , 郑伟1, 2 , 齐永安1, 2 , 李婉颖1 , 吴盼盼1 , 张湘赟1 , 万恩召1 , 和俊淼1 , 李妲1, 2 ]
, 邢智峰, 李婉颖|
|

第一作者简介 许欣,女,1996年生,硕士研究生,主要从事遗迹学研究。E-mail: 3302386307@qq.com。
豫西济源地区下三叠统和尚沟组和中三叠统二马营组均为陆相河湖相沉积,本研究在和尚沟组和上覆二马营组中下段分别识别出遗迹化石5属6种和7属9种,这些化石材料成为探究大灭绝事件后陆地造迹生物复苏的良好材料。运用遗迹网络分析方法构建了和尚沟组和二马营组中下段遗迹网络,相关参数显示自早三叠世和尚沟组沉积期至中三叠世二马营组中下段沉积期,遗迹网络节点增加,平均度、平均加权度和图密度有所降低,网络直径、平均聚类系数和平均路径长度呈明显增加趋势,遗迹网络复杂化以及造迹生物对恶劣环境适应性的增强,这些可能是二叠纪—三叠纪之交灭绝事件(Permian-Triassic Mass Extinction, PTME)后生物复苏的潜在标志。推测PTME后华北地区生物复苏和生态恢复可能遵循着类似的模式,但后续研究仍需陆相实体化石证据的佐证与补充。
, XING Zhifeng, QI Yong’anAbout the first author XU Xin,born in 1996,a masteral degree candidate,is mainly engaged in research on ichnology. E-mail: 3302386307@qq.com.
The Lower Triassic Heshanggou Formation and Middle Triassic Ermaying Formation in western Henan Province represent terrestrial fluvial-lacustrine deposits. Six ichnospecies/five ichnogenera and nine ichnospecies/seven ichnogenera have been identified in the Heshanggou Formation and the lower-middle members of the Ermaying Formation in the Jiyuan area respectively. Based on ichnonetwork analysis,the ichnonetwork of the Heshanggou Formation and the lower-middle members of the Ermaying Formation was constructed to explore the variety of benthic metazoan after the Permian-Triassic Mass Extinction(PTME). The analysis of ichnonetwork shows that during the depositional period of the Heshanggou Formation to the lower-middle members of the Ermaying Formation,the number of nodes,the diameter,the average clustering coefficient and average path length of the ichnonetwork increase significantly. However,the average degree,average weighting degree and density of the ichnonetwork decrease slightly. The complexity of the ichnonetwork and the enhancement of the adaptability of trace makers to harsh environments of the Early and Middle Triassic ichnonetwork has become a potential sign of biological recovery after the mass extinction. Combined with previous studies,the biological recovery and ecological recovery after PTME may follow the similar pattern in North China, though this pattern still need to be corroborated and supplemented by the follow-up studies with more fossil evidences.
二叠纪— 三叠纪之交发生了显生宙以来最大规模的生物灭绝事件, 这一灭绝事件后生物复苏及生态系统重建成为一项重要的研究课题(Chen and Benton, 2012; 戎嘉余和黄冰, 2014; Guo et al., 2019; Roopnarine et al., 2019; Zhao et al., 2020; Huang et al., 2021)。大灭绝之后海洋生态系统按照‘ 金字塔模式’ 自下而上的重建过程已被广泛接受(Chen and Benton, 2012), 但陆相地层实体化石材料的匮乏使得大灭绝之后陆地生态系统的复苏过程并不清晰。遗迹化石作为地质历史时期生物生命活动和行为习性的记录, 在探究地球早期生命演化、生物与环境相互作用、重大地质转折期底栖生态系统对极端环境事件的响应以及古环境和古生态重建方面具有重要意义(郑伟等, 2021)。运用遗迹化石生态学研究方法重建古海洋造迹生物复苏过程, 已在全球多个地区被证实为一种有效的评估大灭绝后海洋生态系统复苏的方法(Chen et al., 2011; Luo et al., 2017), 而将该类方法应用于陆相地层中的遗迹化石群的研究相对较少(Guo et al., 2019; Xing et al., 2021)。
在众多运用遗迹化石复苏生态系统的方法中, 遗迹网络分析是一种较新的定量评估造迹生物群落的手段(Baucon and Fellett, 2013; Baucon et al., 2014, 2015; 党志英和张立军, 2020)。与大多数遗迹学研究的定性方法相比, 遗迹网络分析的优势在于它能够提供遗迹类群的定量描述, 并可基于此重建遗迹类群之间的关联关系, 促进不同尺度上遗迹类群的可视化和综合分析(Baucon et al., 2015)。因此, 通过对地质历史转折期地层自下而上遗迹网络相关参数的分析, 可增进对大灭绝事件后生态系统的认识, 促进生物复苏方面的研究。文中将遗迹网络分析应用于豫西济源地区中下三叠统和尚沟组及二马营组中下段遗迹化石研究, 旨在分析该研究区早中三叠世古遗迹群落组成及分布特征, 结合新遗迹学探究二叠纪— 三叠纪之交灭绝事件(Permian-Triassic Mass Extinction, PTME)后生物复苏前后古遗迹群落的恢复模式。
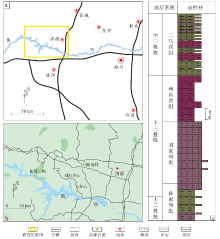
济源地区位于华北板块南缘, 为华北大型内陆克拉通型盆地的一部分, 南部与秦岭— 大别山造山带紧密相邻。该地区基底为太古代到早元古代的结晶岩系及中新元古代和古生代沉积岩, 先后经历三叠纪、早中侏罗世、晚侏罗世— 早白垩世的构造改造和叠加, 主要格局形成于古近纪(徐莉等, 2015)。济源地区下三叠统包括刘家沟组和和尚沟组, 为一套陆相紫红色碎屑岩沉积建造; 中三叠统包括二马营组和油房庄组, 为一套河流相— 湖相沉积序列(Yang et al., 2021)。
下三叠统和尚沟组剖面位于豫西济源方山, 实测厚度约为194.4 m, 岩性以紫红色砂岩、粉砂岩为主, 夹数层薄层粉砂质泥岩和砾岩。中三叠统二马营组剖面位于豫西济源市承留镇仙口村— 大峪镇陡沟村一线(图1-a, 1-b), 实测地层为二马营组中下段, 地层厚约338.5 m, 主要由紫红色泥岩、粉砂岩与灰黄色砂岩组成, 夹灰绿色、灰白色泥岩及薄层砾岩, 发育多层钙质结核(图1-c)。
 | 图1 豫西济源地区中下三叠统剖面位置及岩性柱状图Fig.1 Location and lithology histogram of the Lower-Middle Triassic in Jiyuan area, western Henan Province |
和尚沟组第1层至第4层以紫红色砂岩与泥岩为主, 发育水平层理、楔状交错层理、平行层理和钙质结核, 见少量遗迹化石; 第5层至第8层下部以紫红色砂岩与泥岩的频繁互层为特征, 上部发育厚层粉砂岩与砂岩, 自下而上相继发育平行层理、波状交错层理、水平层理、楔状交错层理和板状交错层理, 见多层磨圆度较好的砾岩, 未见遗迹化石; 第9层为紫红色泥岩与砂岩的频繁互层, 第10至第11层以粉砂岩和砂岩为主, 相继发育楔状交错层理、波状交错层理、板状交错层理和平行层理, 见较多遗迹化石。结合岩性及相关沉积构造, 和尚沟组整体为滨湖沉积(图2)。
 | 图2 豫西济源地区三叠系和尚沟组和二马营组中下段遗迹化石分布及沉积环境Fig.2 Distribution of trace fossils and sedimentary environments in the Triassic Heshanggou Formation and lower-middle members of the Ermaying Formation in Jiyuan area, western Henan Province |
二马营组中下段第12层至第14层发育厚层紫红色、灰黄色砂岩, 夹数层紫红色、灰白色、灰绿色泥岩, 具有“ 砂包泥” 的特征, 厚层砂岩中发育平行层理、楔状交错层理和波痕。其中, 下部为灰黄色砂岩, 发育平行层理和楔状交错层理, 砂岩向两侧厚度变小并由致密的细砂岩变为松散状粉砂岩, 向上发育肉红色泥质粉砂岩和厚层紫红色泥岩, 夹数层薄层砂质层, 泥岩中发育水平层理。砂体外观形态呈顶部较平、底部较凹的槽状。推测整体为3个辫状河“ 河床亚相+洪泛平原亚相” 的旋回。
第15层至第16层下部发育1层薄砾岩, 厚约20 cm, 砾石呈叠瓦状定向排列, 砾径0.3~1.5 cm, 分选性和磨圆性较差, 砾岩层上部夹有少量泥砾, 为河床亚相滞留沉积; 向上为紫红色、灰黄色砂岩夹薄层泥岩, 见平行层理, 层面发育泥裂, 表明处于周期性暴露环境; 再向上变为紫红色泥岩夹灰黄色砂岩和灰白色泥灰岩, 发育钙质结核和水平层理, 具有“ 泥包砂” 的特征, 整体识别为曲流河沉积。第17层至第24层下部发育厚层灰黄色砂岩夹紫红色粉砂岩、泥岩和薄层灰黄色砂岩, 砂岩中发育平行层理和楔状交错层理; 中部以灰黄色、灰白色砂岩与紫红色、灰绿色泥岩的频繁互层为特征, 发育楔状交错层理和平行层理, 见钙质结核; 上部为巨厚层灰黄色砂岩, 向上为灰黄色、紫红色砂岩与紫红色、灰绿色泥岩薄互层, 相继发育平行层理、楔状交错层理、包卷构造和钙质结核, 整体为滨浅湖沉积。岩性及沉积构造分析显示, 二马营组中下段自下而上发育辫状河— 曲流河— 滨浅湖沉积(图2)。
前人曾在豫西济源地区下三叠统和尚沟组鉴定出遗迹化石9属10种(胡斌等, 2009), 此次对济源方山剖面和尚沟组遗迹化石的鉴定共识别出遗迹化石5属6种, 包括垂直居住迹(Skolithos linearis、Cylindricum isp.), 简单水平遗迹(Palaeophylus tubularis、Planolites montanus、Planolites beverleyensis)和具新月形填充的水平遗迹(Taenidium barretti)。具体而言, 和尚沟组下段遗迹化石平均扰动指数为2, 中段及中上段未见遗迹化石, 上段及顶部遗迹化石较为丰富, 生物扰动指数可达到5(图2)。该组遗迹化石以简单的垂直潜穴和水平潜穴为主, 复杂的潜穴较为少见(图3)。
 | 图3 豫西济源地区下三叠统和尚沟组遗迹化石 a-d— Skolithos linearis(Sk); e— Planolites montanus(Pm); f— Planolites beverleyensis(Pb); g— Palaeophycus tubularis(Pt); h— Taenidium barretti(Ta); i— Cylindricum isp.(Cy)。图中硬币直径为2 cmFig.3 Trace fossils of the Lower Triassic Heshanggou Formation in Jiyuan area, western Henan Province |
中三叠统二马营组中下段目前识别出遗迹化石7属9种, 遗迹多样性低至中等、局部具有较高的密度, 除上述和尚沟组出现的遗迹化石外, 还包括具新月形填充和抓痕的Scoyenia gracilis、少量爬行迹Didymaulichnus roualti和Rhizoliths。具体而言, 二马营组下段发育遗迹化石6属8种, 以Skolithos、Palaeophycus、Planolites为主, 遗迹多样性和生物扰动指数相对较高。该组中段发育遗迹化石5属7种, 以Planolites为优势种, 遗迹多样性中等, 厚层灰黄色砂岩中遗迹化石较少, 仅灰黄色砂岩层面上及紫红色泥岩、粉砂岩中生物扰动指数较高, 可达到4~5(图2)。研究区中三叠统二马营组中下段典型遗迹化石照片如图4所示, 遗迹化石具体描述及推测造迹生物见表1。
 | 图4 豫西济源地区中三叠统二马营组中下段遗迹化石 a-b— Skolithos linearis(Sk); c-d— Palaeophycus tubularis(Pt) 和 Palaeophycus alternatus(Pa); e— Scoyenia gracilis(Sc); f-h— Taenidium barrette (Ta); i— Rhizoliths(Rh)和Taenidium barretti(Ta); j-k— Planolites beverleyensis(Pb) 和 Planolites montanus(Pm); l— Didymaulichnus roualti(Di)Fig.4 Trace fossils of the lower-middle members of Middle Triassic Ermaying Formation in Jiyuan area, western Henan Province |
| 表1 豫西济源地区中下三叠统遗迹化石简要描述及潜在造迹生物 Table1 Description of trace fossils and potential trace makers of the Lower-Middle Triassic in Jiyuan area, western Henan Province |
传统的遗迹学方法较为侧重遗迹化石的定性描述, 如遗迹相和遗迹组构的识别, 而将网络分析引入遗迹学研究领域则能够促进遗迹群落的定量描述和古生态系统中各遗迹类群的关系研究。目前, 遗迹网络分析已成为遗迹化石定量描述和系统对比的新兴工具, 在识别重要遗迹类群、遗迹组合及其环境意义解释方面也发挥着重要作用(Baucon et al., 2015)。
在同期造迹情形下, 遗迹网络中相互关联的遗迹类群至少同时出现在同一个样品中, 即它们可能享有共同的环境参数, 而多期造迹则不一定, 基于这一思想, 遗迹网络被构建起来并成为遗迹群落的综合和客观表征(Baucon et al., 2015)。遗迹网络的构建离不开节点和链接, 其中节点个数对应于遗迹属的数量, 链接是遗迹化石之间关联关系的表征, 链接的粗细取决于遗迹化石之间关联关系的强度, 用链接权重表示, 其大小对应于Jaccard指数(Baucon and Fellett, 2013; Baucon et al., 2014, 2015; 党志英和张立军, 2020)。文中将遗迹网络理论应用于下三叠统和尚沟组和中三叠统二马营组中下段遗迹网络的构建和分析, 能够增进对二叠纪— 三叠纪之交灭绝事件后生物复苏的了解。
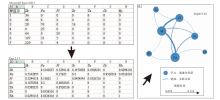
以豫西济源地区中三叠统二马营组中下段遗迹化石为例, 依据Baucon等(2015)提出的方法, 将遗迹化石源数据(图5-a)导入统计软件Past中进行运算即可得到邻接矩阵(图5-b), 将所得邻接矩阵导入Gephi软件中即可生成对应的遗迹网络图形(图5-c), 具体过程如图5所示。
 | 图5 豫西地区中下三叠统遗迹网络绘制流程图(据党志英和张立军, 2020) Sk— Skolithos, Sc— Scoyenia, Pa— Palaeophycus, Pl— Planolites, Ta— Taenidium, Di— Didymaulichnus, Rh— RhizolithsFig.5 Flowcharts of ichnonetwork of the Lower-Middle Triassic in Jiyuan area, western Henan Province(after Dang and Zhang, 2020) |
按这一流程, 分别绘制豫西济源地区中下三叠统和尚沟组和二马营组中下段遗迹网络(图6)。下三叠统和尚沟组中共鉴定出5个遗迹属, 因而该时期遗迹网络中共有5个节点(图6-a), 上覆二马营组中下段共鉴定出7个遗迹属, 该时期遗迹网络中节点即为7个(图6-b)。节点的数量对应于遗迹多样性, 常被认为是环境压力的反映, 孤立的节点可能代表了环境压力较大的生态系统(Buatois and Má ngano, 2011; Baucon et al., 2014)。节点位置无直接的拓扑意义, 因此可为遗迹类群的可视化而随意改变, 如图5-c和图6-b所代表的遗迹网络是等同的。
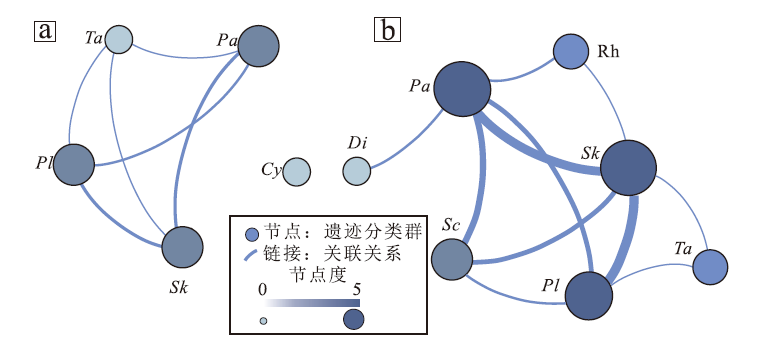 | 图6 豫西济源地区中下三叠统遗迹网络 a— 和尚沟组遗迹网络; b— 二马营组中下段遗迹网络。Fig.6 Ichnonetwork of the Lower-Middle Triassic in Jiyuan area, western Henan Province Sk— Skolithos, Cy— Cylindricum, Sc— Scoyenia, Pa— Palaeophycus, Pl— Planolites, Ta— Taenidium, Di— Didymaulichnus, Rh— Rhizoliths |
此外, 遗迹网络理论中, 节点度、网络直径、图密度、路径长度、聚类系数等参数常被用于遗迹类群的描述和分析。与给定节点相连的链接数被称为节点度(Baucon et al., 2015), 可通过节点的大小度量。平均度为遗迹网络中所有节点度的平均值, 平均加权度为遗迹网络中所有节点的加权度的平均值, 两值均可反映网络整体的连通程度。遗迹网络理论中, 距离被定义为节点间最短路径的长度, 而网络直径是指网络中任意2个节点间距离的最大值, 即遗迹网络中任意2个节点间最多的链接数。图密度为遗迹网络图中实际存在的链接数与最大可能链接数(取决于节点个数)的比率(Baucon et al., 2015)。路径长度是指任意2个节点之间的链接数, 平均路径长度常用于反映节点间的分离程度(Scott, 2000; Baucon et al., 2015)。聚类系数是表征任一节点与给定节点互连度的参数, 遗迹网络理论中较高的平均聚类系数指示联系紧密的遗迹类群的存在, 该值与平均路径长度一起常用于遗迹群落的划分(Boccaletti et al., 2006)。研究区遗迹网络各参数详见表2。
| 表2 豫西济源地区中下三叠统遗迹网络参数 Table2 Ichnonetwork parameters in the Lower-Middle Triassic in Jiyuan, western Henan Province |
由图6-a可知, 和尚沟组沉积期, 研究区内存在5个遗迹类群, 遗迹网络中节点度较高的遗迹类群包括Skolithos和Palaeophycus, Planolites、Taenidium和Cylindricum遗迹类群的节点度较低。研究区内遗迹网络平均度为4.400, 平均加权度为4.236, 网络直径为1, 网络图密度为60.0%, 因此一些遗迹类群并不会与其他遗迹类群同时出现, 如Cylindricum为孤立的节点。该遗迹网络中平均聚类系数为0.583, 平均路径长度为1.000, 显示该沉积期遗迹网络内遗迹类群聚集程度较低。依据图6-b所示, 中三叠统二马营组中下段沉积期研究区内存在7个遗迹类群, 遗迹网络中节点度较大的遗迹类群包括Skolithos、Palaeophycus和Planolites(节点度4~5), Scoyenia、Taenidium和Rhizoliths次之(节点度2~3), Didymaulichnus最小(节点度1)。该沉积期遗迹网络中平均度和平均加权度分别为3.143和1.051, 网络直径为3, 图密度显示该时期只有52.4%的链接存在, 因此一些遗迹类群并不会与其他遗迹类群同时出现, 例如Didymaulichnus与除Palaeophycus之外的其他遗迹类群均未产生链接。该时期平均聚类系数高达0.761, 平均路径长度为1.524, 表明该遗迹网络内遗迹类群聚集程度较为显著。
前人研究指出, 陆相环境中低的遗迹多样性可能意味着极端干燥或潮湿的气候, 而高的遗迹多样性可能与平衡的降水/蒸散比相对应(Baucon et al., 2014)。自和尚沟组沉积期至二马营组中下段沉积期, 研究区遗迹网络节点数由5个增至7个, 遗迹多样性明显增加, 且沉积物由以厚层红层沉积为主向紫红色泥岩、粉砂岩和灰黄色砂岩频繁互层转变, 可能反映了研究区气候由早三叠世的炎热干燥向中三叠世的温暖湿润转变。早三叠世至中三叠世遗迹网络的平均度、平均加权度和图密度有所降低, 网络直径、平均聚类系数和平均路径长度都呈明显增加趋势。早三叠世至中三叠世遗迹网络的复杂程度有所增强, 但结构化程度有所降低, 可能反映了PTME后造迹生物的多样化和生物对环境的适应和选择能力都有所增强, 从而可能成为大灭绝事件后生物复苏的潜在标志。
与同期意大利地区陆相遗迹网络相比, 豫西济源地区下三叠统和尚沟组沉积期遗迹网络节点数量较多, 且遗迹类群间链接的存在表明该时期研究区生态系统结构已有一定程度的恢复。研究区中三叠世遗迹网络中遗迹化石虽同样以简单的、无分支的水平或垂直潜穴为主, 但遗迹化石属种存在一定差异, 而更为显著的差异在于网络结构(Baucon et al., 2014)。意大利地区中三叠世遗迹网络中存在较多孤立节点, 图密度较低, 显示栖息地具有较为明显的碎片化特征, 而研究区遗迹网络结构较为完整, 图密度较高, 显示其生态系统结构化程度较高(Baucon et al., 2014)。产生以上差异的原因可能在于采样及选样误差, 但相对结构化的遗迹网络可能反映出研究区生态系统结构的恢复和重组。
此外, 从沉积环境考虑, 本次在豫西济源下三叠统和尚沟组滨湖沉积中识别出遗迹化石5属6种, 包括Skolithos linearis、Cylindricum isp.、Palaeophyus tubularis、Planolites montanus、Planolites beverleyensis和Taenidium barretti, 中三叠统二马营组中下段滨湖沉积中识别出遗迹化石5属7种, 包括Skolithos linearis、Palaeophycus tubularis、Palaeophycus alternatus、Planolites beverleyensis、Planolites montanus、Didymaulichnus roualti以及Rhizoliths。自和尚沟组至二马营组中下段湖相沉积中, 遗迹多样性稍有增大。遗迹多样性的增加和造迹生物对相对高能条件下砂质沉积环境适应能力的增强, 可能也指示了PTME后造迹生物的复苏。
从以上分析可看出, 遗迹网络能够定量地描述遗迹类群及相互的关联关系, 在重大地质历史转折期遗迹类群及生物复苏方面的研究中也发挥着重要作用, 但这种方法仍存在一定的不足。遗迹网络源数据来源于多块样品, 而样品的采集可能跨越几十米甚至几百米厚的地层, 如何衡量由于古群落不等时对网络分析结果的影响也成为一个难以解决的问题, 当这一方法用于某一地层中遗迹类群的连续演化时或可忽略这一问题, 但当多个剖面进行遗迹网络对比时仍需寻找更好的评价方法。此外, 现有遗迹网络的节点对应于遗迹属, 而无法表述遗迹化石种一级的信息, 生物扰动强度、阶层等方面的信息也无法通过遗迹网络表现出来(党志英和张立军, 2020); 将遗迹网络分析与传统的遗迹学手段和沉积学观测相结合, 能够更好地解释遗迹网络的拓扑模式(Baucon et al., 2014)。总之, 遗迹网络分析为遗迹学提供了一种新颖的研究方法, 但这一理论仍需进一步完善。
豫西济源下三叠统和尚沟组— 中三叠统二马营组中下段发育丰富的遗迹化石, 是研究PTME后华北地区生物复苏和生态恢复模式的良好材料。
1)在豫西济源下三叠统和尚沟组滨湖沉积中识别出遗迹化石5属6种, 包括Skolithos linearis、Cylindricum isp.、Palaeophyus tubularis、Planolites montanus、Planolites beverleyensis和Taenidium barretti; 中三叠统二马营组中下段滨湖沉积中共识别出遗迹化石5属7种, 包括Skolithos linearis、Palaeophycus tubularis、Palaeophycus alternatus、Planolites beverleyensis、Planolites montanus、Didymaulichnus roualti以及Rhizoliths。
2)基于遗迹网络分析方法构建了和尚沟组和二马营组中下段遗迹网络。相关参数显示, 自和尚沟组至二马营组, 遗迹网络节点增加, 遗迹网络的平均度、平均加权度和图密度有所降低, 网络直径、平均聚类系数和平均路径长度呈明显增加趋势。遗迹网络的复杂化以及造迹生物对恶劣环境适应性的增强, 可能反映了PTME后造迹生物的复苏。结合前人研究推测PTME后华北地区生物复苏和生态恢复可能遵循着类似的模式, 后续研究仍需陆相实体化石证据的佐证与进一步补充。
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|