{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
海相遗迹化石对显生宙生物大辐射事件的响应*
[许晴旸1  , 范若颖
, 范若颖1 , 龚一鸣1, 2 ]
, 范若颖]
|
|

第一作者简介 许晴旸,女,1997年生,博士研究生,主要研究方向为遗迹学和沉积学。E-mail: xuqy722@cug.edu.cn。
通过系统梳理前寒武纪和显生宙海相遗迹化石记录及笔者自己的研究,发现在寒武纪生命大爆发、奥陶纪生物大辐射、中生代海洋革命共3次里程碑式的生物大辐射过程中,海相遗迹化石的属级多样性变化和歧异度增减均与生物多样性呈正相关,生物扰动强度和深度明显增加,造迹生物的觅食策略和行为习性多样化明显增多。寒武纪生命大爆发时期,最有代表性的生物行为变化是出现了具有垂向分量的潜穴; 奥陶纪生物大辐射期间,海相遗迹化石的分布逐渐从滨、浅海扩展至半深海和深海,表现为造迹生物群落栖息地的扩张; 中生代海洋革命时期,海相和陆相遗迹化石同步增加,生物对生态空间利用的深度、广度和集约性同步增强,遗迹化石面貌表现为深海雕画迹的多样性和歧异度大幅增加、形态类型多样、多种觅食策略共存。地史时期的海相遗迹化石面貌受环境外因和生物内因控制,表现出形态由简到繁、分布范围由小到大的变化趋势,对生态空间的利用表现为由沉积物表层至浅层再到深层、由二维到三维、由局域(浅水)到广域(浅水和深水以及陆地)的发展,印证了生物获取生态机会的过程。
About the first author XU Qingyang,born in 1997,is a Ph.D. candidate. She is mainly engaged in ichnology and sendimentology. E-mail: xuqy722@cug.edu.cn.
In this paper,we analyzed the trace fossil records of the major Phanerozoic biodiversification events,including the Cambrian Explosion,the Great Ordovician Biodiversification Event,and the Mesozoic Marine Revolution. The ichnodiversity and ichnodisparity of marine trace fossils show a positive correlation with biodiversity during these three biodiversification events. The intensity and depth of bioturbations both increased distinctly. The feeding strategies and the behavior of marine trace-makers became more complex. The most prominent change in the trace-making behavior during the Cambrian Explosion is the appearance of vertical burrows(or vertical elements in burrows). The shallow-marine ichnofauna expanded to the bathyal-marine and deep-marine environments during the Ordovician,as represented by the appearance of morphologically complex deep-marine trace fossils. It shows a further intensification of the exploitation of the marine benthic ecological niches in the Cretaceous. Characteristic change during this period is the significant diversification of deep-marine graphoglyptids,with the development of multiple morphological types and feeding strategies. Under the influences of the environment and biotic changes,marine ichnofaunas became more complicated in morphology and ethology and adopted progressively wider environmental distribution during the Phanerozoic. The marine ichnofaunas displayed increasing penetration depth,transferred from two-dimensional to three-dimensional structures,and radiated from the shallow shelf to the deep sea and continent. These features may be attributed to the process that organisms acquire ecological opportunities.
尽管地质历史上生物大灭绝和大辐射多次交替出现, 但是物种数量和生态类型的增加仍然是生物演化的主旋律(Sepkoski, 1981, 1997; Sepkoski and Shechan, 1983; Fan et al., 2020)。文中重点关注的“ 大辐射” , 指的是生物多样性快速增加的时间段, 涉及到分类单元多样性(α -diversity)、群落生态多样性(β -diversity)和生物地理多样性(γ -diversity)3个方面(詹仁斌等, 2013)。显生宙全球生物大辐射可划分出3个里程碑式的阶段, 分别在寒武纪(寒武纪生命大爆发)、奥陶纪(奥陶纪生物大辐射)和中生代(中生代海洋革命)(Vermeij, 1977; Servais and Harper, 2018; Zhuravlev and Wood, 2018)。
随着科学技术的革新和学科交叉的推进, 关于生物大辐射的研究日渐深入, 生物门类的统计更加客观、系统和全面。实体化石是最直接的生物记录, 但存在2点不足: (1)除少量特异埋藏的化石外, 无硬壳的软躯体动物的实体化石难以保存, 加大了化石保存不完备性所造成的误差(Tweedt and Erwin, 2015); (2)实体化石体现的是生物的解剖学特征, 难以详细记录生物的行为习性和生存策略(Rindsberg, 2012)。这2点不足导致实体化石记录在被用来研究生物大辐射背景下的环境— 生物协同演化关系时具有明显的局限性。
遗迹化石恰好可以弥补实体化石的缺陷。软躯体动物无法留下大量实体化石, 却能够形成丰富的遗迹化石或生物扰动(Seilacher, 2007; Má ngano and Buatois, 2016a), 弥补了地质历史时期一部分缺失的化石记录。同时, 遗迹化石面貌通常与环境相适应, 既记录了造迹生物的行为, 又反映了古环境和古生态(Baucon et al., 2012; Má ngano and Buatois, 2016a)。因此, 遗迹化石是研究生物大辐射时期的古生物和古生态特征以及生物与环境协同演化不可或缺的良好材料。
基于前人对显生宙海相遗迹化石的统计和团队自身的研究, 笔者梳理了前寒武纪到第四纪的海相遗迹化石属级多样性和歧异度(构型差异)变化, 试图阐明显生宙重要生物大辐射期间遗迹化石的属级多样性和歧异度的变化趋势, 查明造迹生物行为对生物大辐射事件的响应模式, 揭示重大生物事件过程中生物行为与环境协同演化的方式。
遗迹多样性(ichnodiversity)的概念相对古生物学中的生物多样性来说意义更局限。广义生物多样性包含遗传多样性、物种多样性和生态系统多样性3个部分, 是一个描述自然界多样性程度的内容广泛的概念(马克平, 1993)。遗迹多样性仅指遗迹属和种的数量, 与物种多样性的含义接近, 为生物和底质相互作用的类型(Buatois and Má ngano, 2011, 2013; 刘梦瑶和张立军, 2018)。
遗迹化石是生命活动在沉积物和其他底质中留下的痕迹, 因此, 对遗迹化石而言, 形态学分类占据重要地位。Gould(1991)于Wonderful Life一书中首次提出了歧异度(disparity)的概念, 用以描述形态结构的差异。近年来, 遗迹学家将歧异度这一概念应用到遗迹化石的研究中, 对遗迹化石的形态构筑类型进行鉴别, 即遗迹歧异度(ichnodisparity)(Buatois and Má ngano, 2011, 2013)。将遗迹多样性和遗迹歧异度相结合, 能够从分类学和形态学2种角度更科学地体现遗迹学在演化古生物学中的价值(Má ngano and Buatois, 2014)。
遗迹丰度(ichnoabundance)是另一个遗迹化石半定量分析的重要指标, 可用于对比某一生境内生物对底质的扰动程度, 弥补了遗迹多样性和歧异度仅能表征遗迹化石类型多寡的片面性(Knaust et al., 2014)。同一个遗迹属或种, 其在1个样本中出现还是在多个样本中出现, 以及其在不同样本中出现的频率, 都是多样性统计和评估的因素。
遗迹化石和现代生物遗迹又被称作生物成因构造(biogenic structures), 该概念包含生物扰动构造(bioturbation structures)和生物侵蚀构造(bioerosion structures)2类。生物扰动构造指生物对未完全固结的沉积物进行破坏和改造留下的痕迹, 而生物侵蚀构造是指生物在坚硬底质(如石质硬底、木头、生物壳体、骨骼等)上机械破坏或化学、生物化学侵蚀留下的痕迹(杨式溥, 1990; Má ngano and Buatois, 2016a)。部分生物扰动构造反映了生物对其栖息地改造和维护的行为过程, 能够改变沉积物的结构和性质, 影响水体物质交换和生态系统功能, 即所谓的生态系统工程(ecosystem engineering)。能够完成这一过程的生物, 被称作生态系统工程师(ecosystem engineers)(Jones et al., 1994)。
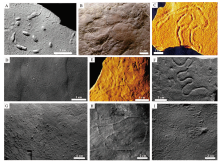
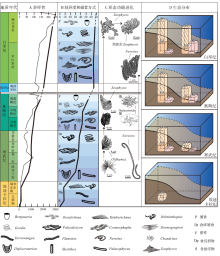
前寒武纪是寒武纪生命大爆发的序章, 将近90%的时间里都不存在宏体动物的生命迹象。直到埃迪卡拉纪, 大量后生动物在极短的时间内骤然出现, 但该次生物爆发和辐射的时间(Tweedt and Erwin, 2015; Wray, 2015; Luzhnaya and Ivantsov, 2019; Moczydł owska et al., 2021)、地点(MacGabhann et al., 2019; McMahon et al., 2020)、进程(Grazhdankin, 2014; Pu et al., 2016; Yang et al., 2016; Wood and Erwin, 2018)、模式(Erwin, 2020a; Evans et al., 2021)仍存在不少争议。埃迪卡拉纪的后生动物大多数都不具硬壳(Sprigg, 1947; Xiao and Laflamme, 2009; Droser and Gehling, 2015; Zhu et al., 2017), 而动物的软躯体难以保存, 从而造成遗迹化石是这一时期生物演化的重要实证材料, 也为前寒武纪两侧对称动物的存在提供了关键性证据(Weber et al., 2007; Rogov et al., 2012; Chen et al., 2013, 2019)。埃迪卡拉纪遗迹化石的多样性、形态类型、分布范围都极其有限, 其仅分布于沉积物表面, 不具有垂向结构和构造, 迄今只报道了9属, 包括Archaeonassa, Epibaion, Gordia, Helminthoidichnites, Helminthopsis, Kimberichnus, Palaeophycus, Torrowangea和Treptichnus (Má ngano and Buatois, 2014; Buatois et al., 2016b)(图1)。
 | 图1 埃迪卡拉纪的遗迹化石记录 A— Treptichnus isp., 采自纳米比亚元古界顶部Huns组, 现存于开普敦南非博物馆(Jensen et al., 2000; Má ngano and Buatois, 2016a); B— Epibaion costatus; C— Archaeonassa fossulata; D— Gordia marina; E— Palaeophycus tubularis; F— Helminthopsis, 来自中国云南东部梅树村剖面石岩头组(Weber et al., 2007); G— Kimberichnus teruzzii, 来自俄罗斯Arkhangel’ sk(Ivantsov, 2013); H— Torrowangea rosei; I— Helminthoidichnites tenuis。其中B, C和E均来自澳大利亚南部Flinders山脉Ediacara段Rawnsley Quartzite(Má ngano and Buatois, 2016a); D, H和I均来自伊比利亚中部地区Rí o Huso剖面Domo Extremeñ o群上部(Jensen and Palacios, 2016)Fig.1 Ichnogenera in the Ediacaran |
埃迪卡拉纪造迹生物最典型的特征是其仅生活于沉积物表面, 形成浅表扰动。仅发现6种生物扰动的构筑类型, 包括简单水平拖迹、被动充填水平或倾斜潜穴、简单主动充填水平或倾斜潜穴、椭圆形印痕和抓痕及具有水平或垂直分支的水平潜穴。生物侵蚀构造包括圆柱形垂直或倾斜钻孔、圆孔状或凹坑状钻孔、球形钻孔(Buatois et al., 2016b; Má ngano and Buatois, 2016a), 最大扰动深度只有1 cm(Má ngano and Buatois, 2014), 最大生物扰动指数为1(Taylor and Goldring, 1993; Má ngano and Buatois, 2014)。埃迪卡拉纪广泛分布坚固、密实的微生物席底质, 在席底的限制下, 该时期的动物仅具有4种生活方式: (1)席底固着者(mat encrusters)— — 附着在微生物席表面, 以滤食为生; (2)席底刮擦者(mat scratchers)— — 在微生物席表面刮擦, 并不完全破坏微生物席, 目前仅存在Kimberella一种代表生物; (3)席底黏着者(mat stickers)— — 以圆锥形外壳附着于微生物席, 并随微生物生长而迁移, 以固定在微生物席表面; (4)席下矿工(undermat miners)— — 在微生物席下部挖掘水平潜穴, 以微生物席最下层腐烂的生物为食(Seilacher, 1999)。觅食策略包含食悬浮物、食碎屑物和食沉积物(Seilacher et al., 2005)。造迹生物大都聚集于滨、浅海环境中, 存在极大的生态位重合现象, 深海环境中遗迹属数量极少且形态简单, 生态分布范围集中在滨岸带和近岸带(Má ngano and Buatois, 2016a)。
寒武纪生命大爆发在地球环境与生命过程的演替中具有划时代意义, 关于这次事件的全貌和意义目前仍存在争议(Maloof et al., 2010; Erwin et al., 2011; Erwin, 2015; Má ngano and Buatois, 2016a)。遗迹化石作为后生动物(特别是两侧对称动物)行为的记录, 为寒武纪生命大爆发事件的进程提供了重要参考。结合遗迹化石已有报道来看, 此次生命大爆发是真实存在的(Zhang and Shu, 2021)。寒武纪遗迹化石表现出属级多样性和歧异度爆炸性增长, 出现了垂向构造, 实现了从二维到三维的突破。生物扰动强度、扰动深度都大幅增加, 对生态环境产生了重大影响。其在垂向和横向分布空间上得到显著拓展, 体现出生态分布立体性扩张的特点。
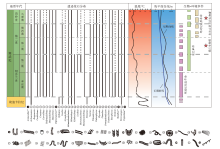
寒武纪遗迹化石的多样性增长分为2个阶段(图2)。(1)第1阶段是指从寒武纪幸运期到第四期(538.8± 0.2 Ma~509.0 Ma), 该阶段遗迹化石的属级多样性快速增长, 增长速率由陡到缓。其中幸运期— 第二期共记录了59个遗迹属, 比埃迪卡拉纪增长了490%; 归为27种形态构筑类型, 比埃迪卡拉纪增长了350%。第三期— 第四期多样性增长速率降低, 共75个遗迹属, 比第二期增长了27%; 归为32种形态构筑类型, 比第二期增长了18%。(2)第2阶段是指从寒武纪乌溜期到第十期(509.0 Ma~485.5± 1.9 Ma), 该阶段遗迹化石的属级多样性和歧异度变化趋于平稳。苗岭世遗迹属降低到72个, 芙蓉世恢复了增加的趋势, 芙蓉世末期全球共记录77个遗迹属, 较第二世增长不足3%; 共33种形态构筑类型, 较第二世仅增加了1种(Buatois et al., 2016b)。寒武纪生物侵蚀构造属级多样性和歧异度数量变化平缓, 没有表现出爆炸性增长。
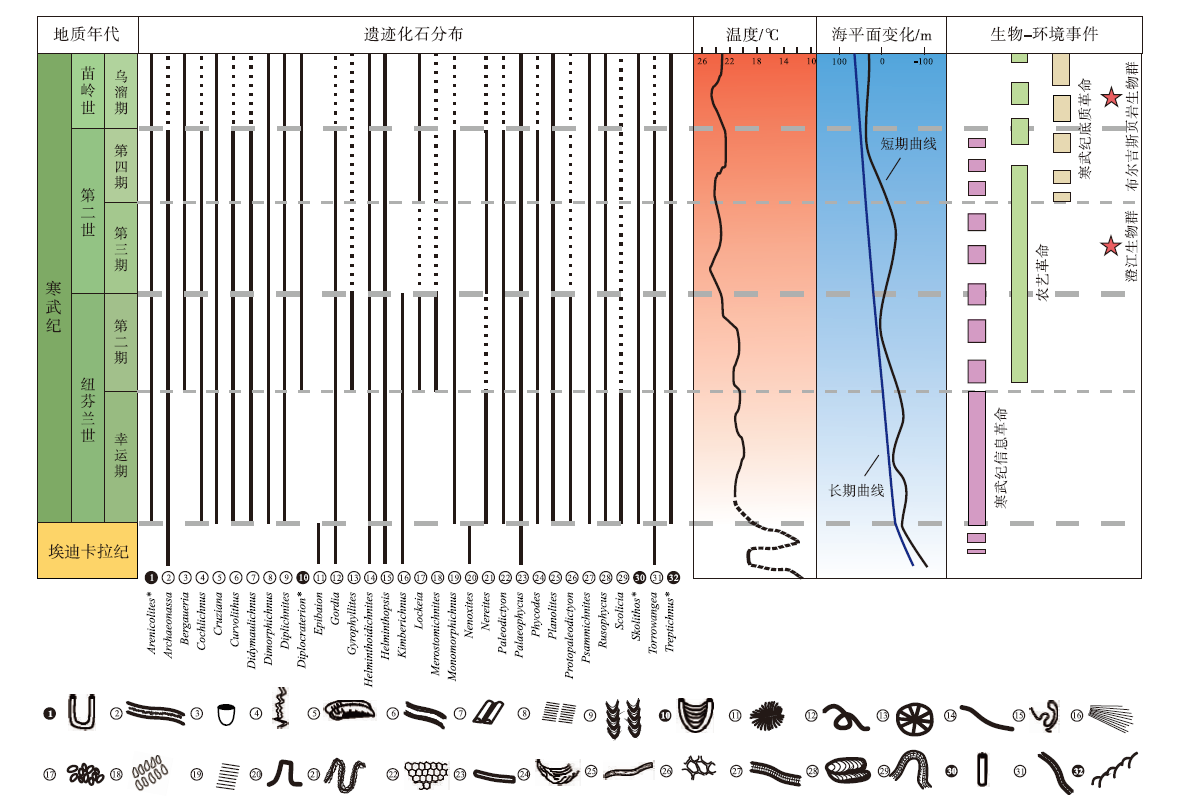 | 图2 埃迪卡拉纪— 寒武纪之交遗迹化石分布与温度、海平面变化及其他生物— 环境事件的关系(据Má ngano and Buatois, 2017等改编) 第1栏为埃迪卡拉纪— 寒武纪典型的遗迹化石分布格局, “ * ” , “ ❶” 表示垂向结构或具垂向分量的遗迹属(数据来源于Crimes and Anderson, 1985; Elicki et al., 2013; Pandey et al., 2014; Singh et al., 2014; Wang et al., 2015; Jensen and Palacios, 2016; Shahkarami et al., 2017); 第2栏为埃迪卡拉纪到寒武纪乌溜期温度曲线(修改自Scotese et al., 2021); 第3栏为埃迪卡拉纪到寒武纪乌溜期海平面曲线(修改自Haq and Al-Qahtani, 2005); 第4栏展示了寒武纪典型生物— 环境事件, 包括寒武纪信息革命、农艺革命和寒武纪底质革命, 并标明了特异埋藏生物群(修改自Má ngano and Buatois, 2017; Erwin, 2020b)。时间标尺参照国际年代地层表2022/10版Fig.2 Sketch showing distribution of trace fossils, and its’ relation with temperature, sea-level change and typical bio-environmental events through the Ediacaran-Cambrian transition(modified from Má ngano and Buatois, 2017 etc.) |
寒武纪发生了3次有代表性的“ 生物革命” 事件, 分别是寒武纪信息革命、农艺革命和寒武纪底质革命(图2)。生态环境的多样性、复杂性和不均一性显著增加, 可能导致动物进化出了更复杂的运动行为、更复杂的感知器官和更强的感知能力, 这一过程被称作寒武纪信息革命(Cambrian Information Revolution)。寒武纪信息革命发生在幸运期(Plotnick, 2007, Plotnick et al., 2010), 直接表现为水平潜穴的分支增多、潜穴深度增大、形态趋于复杂、出现生物钻孔和垂向生物遗迹等(Zhu, 1997; Kobluk et al., 2016; Má ngano and Buatois, 2017)。寒武纪首次出现了内生水平潜穴Cruziana(Parcha and Pandey, 2011)。遗迹属Treptichnus在寒武纪初期就迅速增加至8个种, 种级多样性、形态复杂程度和化石数量都远高于前寒武纪, 其中具垂向分量的Treptichnus pedum被用以划分寒武纪和前寒武纪界线(Zhu, 1997; Jensen et al., 2000; Droser et al., 2002; Parcha and Singh, 2010; Wilson et al., 2012)。Psammichnites这样高度复杂的牧食迹出现, 也体现出了生物更复杂、更具有创新性的觅食策略, 表明底栖生物已具备在更多不同类型的海洋环境中获得食物资源的能力(Plotnick, 2007; Plotnick et al., 2010; Má ngano and Buatois, 2016a)。海生动物出现了矿化的硬壳, 大型生物壳体钻孔类生物遗迹得以形成, 反映出更复杂的种间关系(Kobluk et al., 2016)。生物的垂向掘穴行为形成了较深阶层、具有垂向分量的遗迹化石, 包括Arenicolites, Diplocraterion, Syringomorpha (图2)(Má ngano and Buatois, 2017)。生物扰动强度增加和深度增大, 最大生物扰动指数为6, 潜穴深度大多在6 cm以内, 少量最极端的记录来自垂向潜穴Skolithos, 尽管它通常被构造断裂破坏, 很难测量完整的长度, 但它所体现的扰动深度仍然突破了历史记录(Hallam and Swett, 1966; Taylor and Goldring, 1993; Má ngano and Buatois, 2014, 2017; Buatois et al., 2020)。不仅仅是垂向遗迹化石, 水平压痕状生物遗迹如Rusophycus, 也出现扰动加深(Má ngano and Buatois, 2017)。
农艺革命和寒武纪底质革命, 都可以泛指生物对环境产生改造而使前寒武纪坚固的微生物席底消减的过程(Seilacher and Pflü ger, 1994; Bottjer et al., 2000), 但二者侧重点不同。农艺革命(Agronomic Revolution)强调寒武纪的混合底取代前寒武纪的微生物席底这一过程:在纽芬兰世第二期, 中、深层食悬浮物者通过掘穴行为, 破坏了质密的微生物席底, 加强了水柱和沉积物之间的物质交换; 食悬浮物者可以通过水柱更轻易地获取有机质颗粒并通过生物沉积作用来富集有机物, 从而进一步促进海底食沉积物动物多样化(Hily, 1991; Erwin and Tweedt, 2011; Má ngano and Buatois, 2014)。寒武纪第二世第四期发生的寒武纪底质革命(Cambrian Substrate Revolution), 则更强调底栖生物群落在底质改变过程中的响应, 既受到农艺革命的影响, 也是生物在自然选择下进化的结果(Seilacher et al., 2005; Má ngano and Buatois, 2017)。始于强烈的生物扰动, 2次革命相继出现, 对后生动物面貌和底栖生态产生了深远影响。
寒武纪生物扰动的阶层结构始终相对简单。浅层食沉积物者和食碎屑物者占据浅海和边缘海低能环境, 深层食碎屑物者只占据低能海洋环境, 食悬浮物者则占据高能潮下带, 它们各自为营、分层清晰, 体现出了生物对垂向生态空间的利用仍然有限(Má ngano et al., 2013; Buatois et al., 2020)。
从环境分布上看, 遗迹化石表现出扩张趋势。寒武纪浅海遗迹化石极为丰富, 寒武纪纽芬兰世— 第二世浅海最大属级α 遗迹多样性(单一相带或单一环境带内的遗迹多样性)为28种, 包含Skolithos, Arenicolites, Diplocraterion, Cruziana, Asterosoma, Rhizocorallium, Teichichnus等多个Skolithos遗迹相和Cruziana遗迹相的特征遗迹属(Crimes and Anderson, 1985; Parcha and Pandey, 2011; Elicki et al., 2013; Wang et al., 2015)。相比之下, 深海α 遗迹多样性只有8种, 以Oldhamia广泛分布为特征(Jensen et al., 2013)。特别值得注意的是, 在寒武纪浅海沉积物内观察到了主要分布于深海的遗迹属Oldhamia(Seilacher et al., 2005; Jensen et al., 2013), 甚至观察到了更新的地层中常见于深海的雕画迹Protopaleodictyon, Paleodictyon(Morgan et al., 2019; Malekzadeh and Wetzel, 2020)。此外, 通常认为代表远洋深海的Nereites遗迹相主要由牧食迹、耕作迹和觅食迹组成, 而寒武纪时牧食迹、耕作迹和觅食迹鲜见于深海, 反而多见于浅海, 因此只有在寒武纪后才能将Nereites遗迹相看作深海沉积环境的特征遗迹相(Crimes and Anderson, 1985; Jensen and Mens, 1999; Orr, 2001; Jensen et al., 2013)。这可能与深海遗迹化石起源于浅海有关, 即浅海遗迹化石从寒武纪开始向远岸、深海迁移(Crimes and Fedonkin, 1994; Buatois et al., 2020)。寒武纪也首次出现遗迹化石向半咸水、陆表海等环境扩张的趋势(Má ngano and Buatois, 2004; Minter et al., 2017; Buatois et al., 2020), 半咸水环境出现了节肢动物遗迹Cruziana, Rusophycus, Diplichnites和垂直潜穴Skolithos, 陆表海环境则以Skolithos和Cruziana遗迹相为代表(Buatois, 2005; Davies et al., 2009)。
奥陶纪生物大辐射是显生宙历史上规模最大的一次生物辐射事件, 几乎贯穿了整个奥陶纪, 突出表现是物种数在门以下级别, 特别是属、种级别出现显著增长, 全球海洋属级生物增量超过3倍(Sepkoski, 1997; Buatois et al., 2016b; Fan et al., 2020; Deng et al., 2021)。奥陶纪遗迹化石的属级多样性和歧异度有显著增加, 尤其是深海生物遗迹。遗迹化石生态分布由浅海扩展到深海, 形态由简单到复杂、由浅表阶层扩展到中深阶层, 生物觅食行为模式更多样, 生态系统结构更复杂。
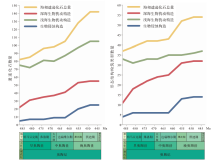
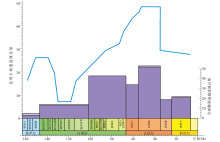
奥陶纪生物扰动和生物侵蚀构造的多样性总量增长至142种, 比寒武纪末增加了73%, 歧异度总量达54类, 比寒武纪增长了46%, 以深海遗迹多样性和歧异度大幅增长而浅海增幅相对较小为特征(Buatois et al., 2016b)(图3)。与大多数实体化石的3幕式辐射不同(Servais et al., 2010; Stigall et al., 2020), 深海遗迹属的增长分为2幕: 第1幕为早奥陶世— 中奥陶世大坪期, 深海遗迹属从22属增长至37属, 生物侵蚀构造仅增加4个遗迹属; 第2幕发生在中奥陶世达瑞维尔期— 晚奥陶世, 深海遗迹属总量达55属, 相较于晚寒武世共增长150%, 生物侵蚀构造遗迹属总量达25属, 相较于晚寒武世增加了400%(Buatois et al., 2016b)。
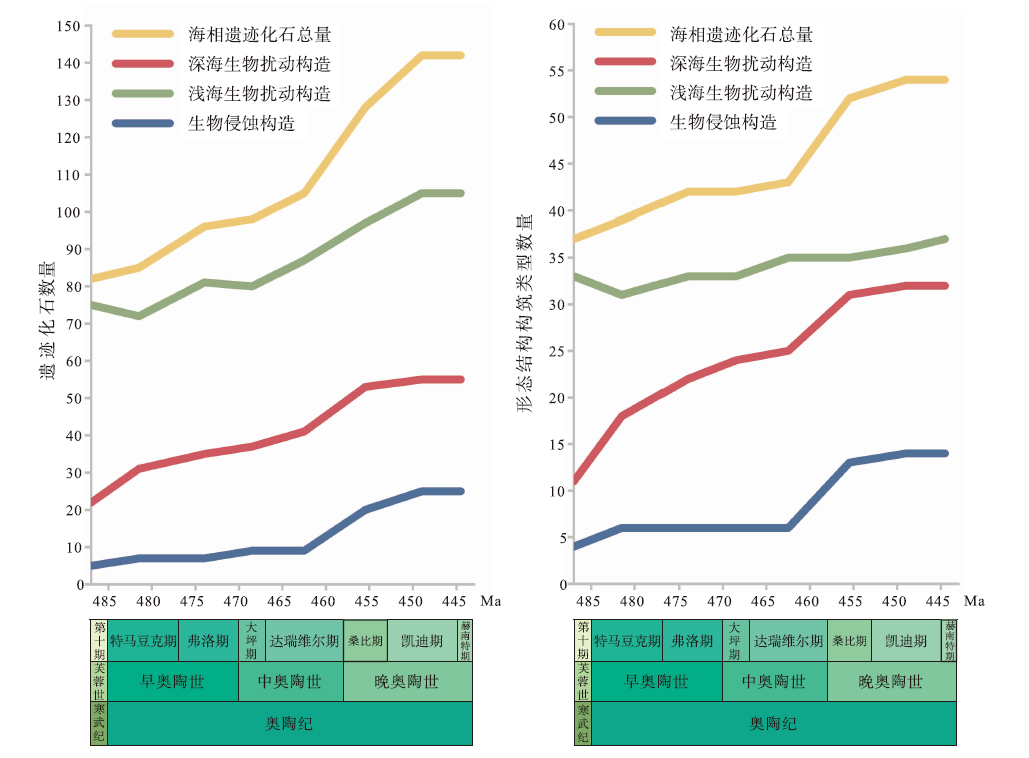 | 图3 奥陶纪遗迹化石多样性和歧异度变化(修改自Buatois et al., 2016b) 左图为遗迹多样性变化趋势, 右图为遗迹歧异度变化趋势。时间标尺参照国际年代地层表2022/10版Fig.3 The trend of ichnodiversity and ichnodisparity through the Ordovician(modified from Buatois et al., 2016b) |
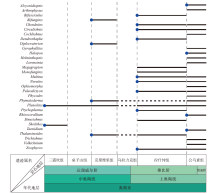
遗迹歧异度的增加同样分为2幕, 与多样性的增长趋势几乎一致(图3)。其中浅海遗迹歧异度几乎没有明显增长, 仅比寒武纪增加了4种。深海遗迹化石歧异度则大幅增长, 总量达32种, 比寒武纪增加了21种, 以形态较复杂的深海雕画迹出现为显著特征(Crimes et al., 1992; Buatois et al., 2009)。生物侵蚀构造的形态构筑类型从4种增加至14种, 奥陶纪最早出现的新增生物侵蚀构造是棒状钻孔Gastrochaenolites和放射状钻孔Podichnus (Buatois et al., 2016b)。笔者的研究资料表明, 华北板块西缘在奥陶纪大辐射期遗迹化石的多样性和歧异度增幅明显, 从中奥陶世初期的三道坎组的2属2种构型, 到晚奥陶世晚桑比期的公乌素组增加为21属16种构型(图4)。
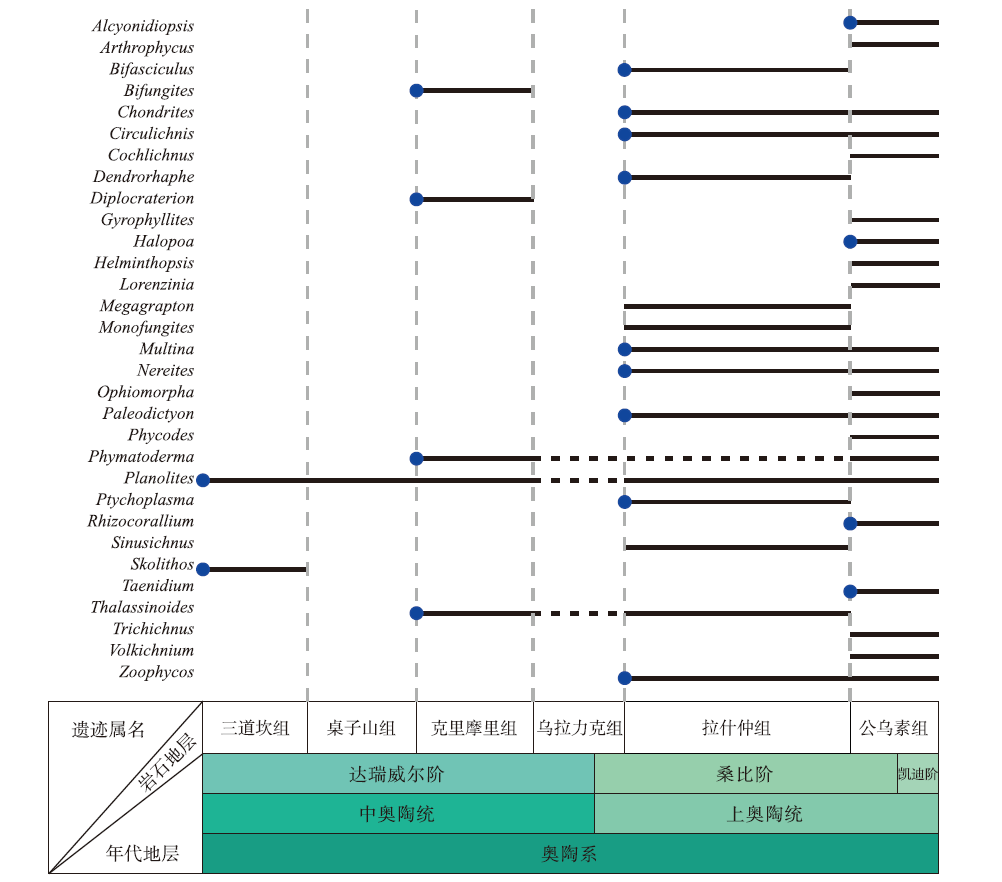 | 图4 华北板块西缘奥陶纪遗迹化石的产出和分布 蓝色圆点表示在该地区首次出现的形态构筑类型, 其中Megagrapton和Paleodictyon属于同一种形态类型, 且在同一时间段出现, 因此, 仅选取一个标明。该图根据该地区现有报道(李日辉, 1993; 费安玮, 2000, 2001; 费安玮等, 2004; Fan et al., 2021)绘制, 遗迹属未延续仅表示更新的地层中尚未报道, 并非已灭绝。时间标尺参照国际年代地层表2022/10版Fig.4 Ichnofossils from the Ordovician of western Inner Mongolia, North China |
奥陶纪潜穴深度由浅到深。Thalassinoides是体现奥陶纪生物扰动行为变化的重要代表性遗迹属, 它在奥陶纪出现了2种类型: 一种是简单迷宫形态(图5-a), 在早、中奥陶世占据主要地位, 生物扰动深度不超过6 cm; 另一种是具有复杂三维结构的迷宫到箱状形态(图5-b), 在晚奥陶世大量出现(尽管此时也存在简单迷宫状Thalassinoides), 扰动深度可达30 cm(Droser, 1987; Droser and Bottjer, 1989)。这导致底质内的阶层结构更复杂, 觅食行为由以食沉积物为主转变为食沉积物、食悬浮物、主动捕食、耕作等多种方式并存(Má ngano and Buatois, 2011; Buatois et al., 2020)。
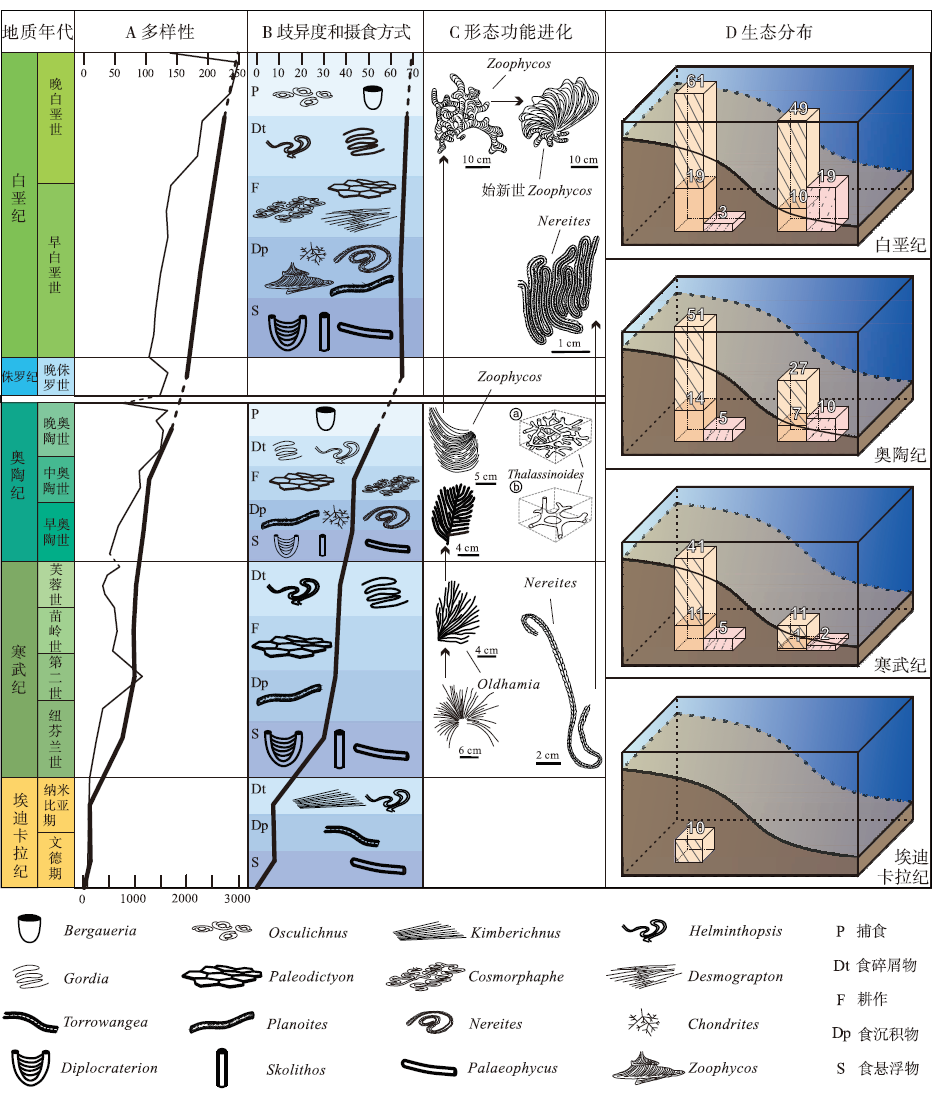 | 图5 海相遗迹化石对显生宙3次生物大辐射进程的响应模式对比 A— 粗线条为海相生物扰动构造多样性曲线, 参照表格上部比例尺(修改自Buatois et al., 2016b; Má ngano and Buatois, 2016b), 细线条为海相生物多样性曲线, 参照表格底部比例尺(修改自Sepkoski, 1996; Erwin, 2020b); B— 海相生物扰动构造歧异度曲线(修改自Buatois et al., 2016b; Má ngano and Buatois, 2016b), 及各时期不同生物觅食方式遗迹化石代表; C— 遗迹形态功能进化示意图, 以遗迹属Oldhamia, Nereites, Zoophycos, Thalassinoides为例, 其中Thalassinoides仅为立体形态示意图, 无比例尺(Seilacher, 1974; Droser, 1987; Zhang et al., 2015); D— 各时期海相遗迹化石环境分布示意图, 选取了各年代中延限较长的遗迹属作为统计对象, 左侧为浅海环境, 右侧为深海环境, 其中橙色柱状图为遗迹属总量, 深橙色部分表示其中垂向或具有垂向分量的遗迹属, 浅粉色柱状图为其中的雕画迹(Buatois et al., 2016b; Uchman, 2003)Fig.5 Summary of marine ichnofossil responses during three major Phanerozoic biodiversification events |
奥陶纪近岸浅海和远岸深海2种环境的遗迹化石都表现出复杂化。奥陶纪浅海生物的行为体现出极大的创新性, 如出现了仅仅分布于奥陶纪— 志留纪地层中的“ J” 型遗迹化石Daedalus (Seilacher, 2000); 始于寒武纪的遗迹属Rosselia在奥陶纪出现了向上运动形成的纺锤形结构, 推测是为了避免被单向流动的沉积物埋藏而产生的行为, 但在寒武纪没有同类型结构的记录(Desjardins et al., 2010; Buatois et al., 2016a); 原本在寒武纪浅海密集分布的Skolithos在奥陶纪明显减少(Droser, 1991); Thalassinoides由简单迷宫结构逐渐转变为复杂箱状结构占主导(Droser and Bottjer, 1989; Má ngano and Buatois, 2016a)。浅海环境中属于双壳类的钻孔生物, 属于头足和腹足类的肉食性动物及腕足类、珊瑚、层孔虫、海绵、苔藓虫等底栖固着生物大量出现, 导致生物侵蚀构造多样性和歧异度同步增加(Servais et al., 2010; Servais and Harper, 2018; Stigall et al., 2019)。如出现双壳动物造迹形成的棒状钻孔Gastrochaenolites、深层槽型钻孔Petroxestes, 腹足动物捕食形成的生物壳体的裂口Caedichnus, 苔藓虫形成的不规则洞穴状钻孔Sanctum, 层孔虫、腕足类形成的深层钻孔痕迹Trypanites和Palaeosabella, 海绵形成的放射状钻孔Cicatricula, 这些钻孔对奥陶纪碳酸盐岩沉积地层的侵蚀作用显著增强(Tapanila and Copper, 2002; Benner et al., 2004; Wilson and Palmer, 2006; Stafford et al., 2015; Vinn and Wilson, 2015; Vinn et al., 2019; Erickson, 2020)。
奥陶纪深海遗迹化石以具有多种几何形态、复杂精美的深海雕画迹为特征。雕画迹是寒武纪以后深海Nereites遗迹相的典型遗迹化石, 记录了复杂的生物觅食、牧食和耕作(培植微生物)过程(Uchman and Wetzel, 2012)。典型遗迹属包括网状雕画迹Megagrapton、Protopaleodictyon和Paleodictyon, 螺旋状雕画迹Spirorhaphe, 规则蛇曲形雕画迹Cosmorhaphe, 放射状雕画迹Lorenzinia等(Crimes et al., 1992; Buatois et al., 2009; Fan et al., 2021)。寒武纪深海广泛分布的Oldhamia, 在奥陶纪出现了生物行为优化的现象, 分支更加紧密, 排列更加规则(Seilacher, 1974)(图5-Oldhamia)。
遗迹化石分布范围扩张表现为深海遗迹化石多样性和歧异度激增。在寒武纪乃至前寒武纪出现并且分布于浅海的遗迹属Nereites, Helminthopsis, Helminthoidichnites, Glockerichnus, Paleodictyon等, 在奥陶纪已迁移至深海, 说明部分奥陶纪深海遗迹化石起源于浅海, 造迹生物从寒武纪晚期由近岸向远岸扩散(Crimes et al., 1992; Crimes and Fedonkin, 1994)。
奥陶纪生物大辐射后, 生物几乎覆盖了整个海洋生态空间。此后, 自志留纪直到晚古生代末期, 生物遗迹面貌的变化包括遗迹化石从海洋扩展到陆相环境中和造迹生物对生态空间的高效利用; 海洋环境遗迹多样性高, 形态构筑类型丰富多样。中生代, 不同进食习惯的类群相互抑制作用加强, 底栖固着生物减少, 食悬浮物者遗迹丰度降低。
自早古生代后期到晚古生代, 遗迹化石展现出大量向非海洋环境扩张的潜力。志留纪后期, 非海洋环境遗迹化石开始出现。如Marriott(2009)等在潮汐间歇性入侵形成的半咸水湖泊中发现了多种节肢动物、多足类动物停息、爬行、觅食的痕迹, 这些可能的造迹生物此前大多栖居于咸水环境。即使迁移本身可能由偶发性事件导致, 遗迹化石仍体现了造迹生物生态范围向陆地扩散的趋势(Buatois et al., 2005; Minter et al., 2017)。志留纪长期受淡水影响的半咸水环境中也出现了脊椎动物遗迹, 包括鱼类游泳迹Undichna, 吻捕迹Osculichnus(Knaust and Minter, 2018; Fan et al., 2019), 目前发现的最早的鱼类停息迹则产自志留纪的浅海相地层中(Ayranci et al., 2022), 体现了志留纪遗迹化石生态分布之广泛, 类型之多样。泥盆纪形成了完善的陆生脊椎动物和无脊椎动物生态系统, 到泥盆纪后期非海洋环境遗迹化石大量出现。石炭纪, 淡水鱼类、节肢动物、环节动物和软体动物大幅多样化, 遗迹化石定居非海洋环境在中生代到来前已成定局(Maples and Archer, 1989; Minter et al., 2017)。
海洋环境遗迹多样性延续了奥陶纪晚期的盛况(Toom et al., 2019)。滨岸、浅海有大量形态各异、功能多样的常见生物遗迹, 如垂直潜穴Skolithos, U形和Y形潜穴Diplocraterion, Arenicolites, Polarichnus, 哑铃型和箭镞状潜穴Arthraria, 塞子状潜穴Bergaueria, 具有成对沟槽的移迹Cruziana, 水平弹簧状潜穴Helicodromites, 单层蹼状构造Rhizocorallium, 足迹和抓痕Monomorphichnus, 树枝状潜穴Chondrites等(Uchman, 2004; Sedorko et al., 2019; Toom et al., 2019)。同种遗迹化石的生态分布发生扩张, Má ngano等(2020)报道了西班牙北部中泥盆世滨海相地层中非三叶虫的真节肢动物在沉积物中蜕皮以躲避天敌的痕迹, 而此前仅报道过浅海相地层中三叶虫的蜕皮行为。深海遗迹化石群在这一阶段的大部分时间中保持相对稳定。Uchman(2004)对全球深水相剖面的深海遗迹属统计表明, 自奥陶纪末到石炭纪末, 深海遗迹化石的多样性、歧异度都保持非常稳定的水平, 单个剖面中最多能观察到27个遗迹属。Wetzel等(2007)在阿曼三叠纪深海相剖面中报道了32个遗迹属, 是石炭纪末到侏罗纪已知多样性最高的深海遗迹化石群落。
中生代, 以腕足动物为主的动物群被以软体动物为主的动物群所取代, 觅食方式从固着底栖悬浮式觅食变为移动悬浮式觅食和食沉积物, 主动捕食行为增加, 是“ 中生代海洋革命” 的表现(Vermeij, 1977; Harper and Skelton, 1993; Baumiller et al., 2010)。这一时期放牧行为加剧, 底栖生物群落受到影响, 如生物内栖化加重, 或是因固着生物被剥离后无法再次接合, 导致固着生物减少(Vermeij, 1977)。
白垩纪延续了中生代海洋革命的进程, 使中生代生物辐射达到顶峰, 遗迹化石类型丰富多样, 尤以深海雕画迹的繁盛为特点。白垩纪是深海遗迹化石多样性、歧异度陡增的时期。浅海遗迹多样性始终保持高值, 形态多样, 直至K-E大灭绝前夕才降低。生物遗迹形态进一步优化, 造迹生物行为集约性进一步优化。
白垩纪全球深海遗迹属总量达侏罗纪的2倍, 其中深海雕画迹属级多样性增长尤为显著(Uchman, 2003; Uchman and Wetzel, 2012)(图6)。晚侏罗世提塘期到早白垩世阿普第期深海遗迹属级多样性增加, 但在早白垩世末阿尔必期跌落到极低的水平。自晚白垩世初, 深海遗迹多样性、歧异度持续增加。尽管此前的奥陶纪曾出现过深海遗迹属的小幅度辐射, 但其群落数量、辐射规模、形态丰富程度、同种环境下遗迹化石群落组成的稳定性远不及白垩纪晚期至新生代水平(Uchman, 2003)。与此同时, 雕画迹在深海复理石相遗迹中的占比逐渐增大(Uchman and Wetzel, 2012), 说明了雕画迹化石组合对于深海环境的表征意义。
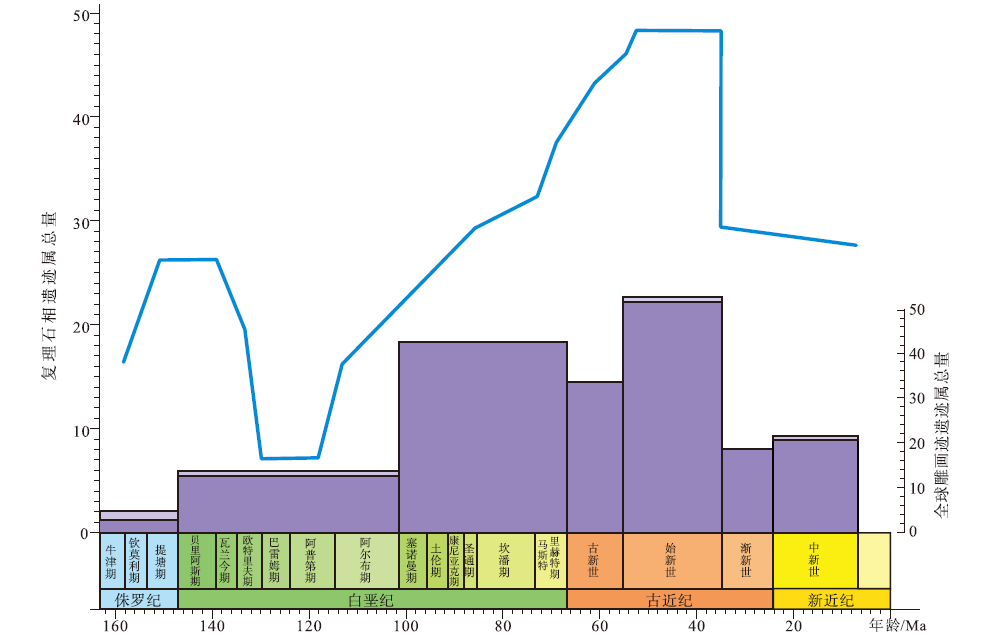 | 图6 侏罗纪至新近纪复理石相遗迹化石与全球雕画迹属级多样性变化趋势图(修改自Uchman and Wetzel, 2012) 折线图为复理石相的遗迹属总量变化图; 柱状图为全球雕画迹遗迹属总量图, 上部浅色部分表示暂时被归入了雕画迹的遗迹属。时间标尺参照国际年代地层表2022/10版Fig.6 Trends of ichnogeneric diversity in flysch formations and the number of global graphoglyptid ichnogenera from the Jurassic to Neogene(modified from Uchman and Wetzel, 2012) |
浅水环境遗迹属多样性在K-E界线之下保持高值。Frey和Howard(1990)在美国犹他州晚白垩世近滨— 远滨相地层中, 统计到包括Ancorichnus, Chondrites, Ophiomorpha, Planolites, Rosselia, Skolithos, Thalassinoides, Teichichnus等共17个遗迹属和12种形态构筑类型。形态丰富, 含具复杂垂向结构的遗迹化石, 浅、中、深层扰动共存。临近K-E界线遗迹多样性降低, 常见遗迹属仅为Chondrites, Zoophycos, Planolites, Thalassinoides, Ophiomorpha和Alcyonidiopsis, 但大灭绝事件前后遗迹化石面貌几乎不变(Má ngano and Buatois, 2016b)。
白垩纪的遗迹化石形态优化现象尤其体现在复杂的深海遗迹化石中。如蛇曲形觅食迹Nereites在寒武纪蛇曲形态简单, 排列稀疏; 随着时间推移, 直至白垩纪, Nereites蛇曲形态明显更规则、排列更紧密。蹼状遗迹化石Zoophycos古生代时仅有单个或双个蹼层, 并且蹼层形态小而简单, 无边缘管; 中生代Zoophycos的蹼层数增多, 出现多级蹼纹; 新生代演化为多蹼层、大而复杂的形态, 并出现了叶状边缘, 结构由简到繁, 整体形态由单一粗糙变为复杂精美(Olivero, 2003; Zhang et al., 2015)(图6-Nereites, Zoophycos)。
自古近纪开始, 全球生物在经历了白垩纪生物大灭绝后进入了新生代动物全面辐射阶段(Archibald and Deutschman, 2001)。新生代海相遗迹类群没有发生大的改变或突破, 不管是在深海还是浅海环境, 遗迹多样性、歧异度、遗迹化石群落组成、底栖生物生活方式、生态群落结构等方面都保持着相对稳定, 遗迹属数量丰富(Carmona et al., 2008)。
海相遗迹多样性、歧异度在此时期保持相对稳定。如Rodrí guez-Tovar等(2016)在西班牙南部的深海生物遗迹群落中共发现20个遗迹属, 其中雕画迹有12属: Belorhaphe, Cosmorhaphe, Desmograpton, Glockerichnus, Helicolithus, Helminthorhaphe, Lorenzinia, Megagrapton, Paleodictyon, Paleomeandron, Spirorhaphe, Urohelminthoida, 该遗迹组合属于典型的深海Nereites遗迹相。始新世以后, 仅有少数几个新的雕画迹属出现(Uchman, 2003)。
浅海环境遗迹化石报道较少。新生代浅海遗迹化石组合与中生代晚期的浅海生物遗迹群落相似, 以典型的Skolithos和Cruziana遗迹相为代表(Buatois et al., 2002; Uchman and Gazdzicki, 2006; Singh et al., 2008)。
深海遗迹多样性在古新世— 始新世达到顶峰, 此时深海雕画迹占深海遗迹属的比例达到最高值, 全球深海遗迹化石多样性从始新世开始下降(Uchman, 2004)。目前缺少中新世以后深海生物遗迹的研究报道, 这主要和上新世— 全新世这些更新的地层还深埋在海底之下、未经构造运动抬升并出露于地表有关。
生物大辐射具有复杂的形成机制, 但环境和生物之间的正向促进始终是其主旋律。通过对前寒武纪与显生宙之交海相遗迹化石的整体梳理可以看出, 遗迹化石记录对3次生物大辐射事件的响应特征可概括为以下4个方面(图5)。
大辐射事件都伴随着遗迹多样性的增加, 但不同时期辐射的侧重点不同, 歧异度变化与多样性变化趋势基本一致。前寒武纪遗迹化石仅有10个遗迹属, 6种形态构筑类型, 寒武纪生命大爆发后全球遗迹属增长至77个, 形态构筑类型增长至33个, 其中浅海生物扰动构造遗迹属和形态构筑类型增量最大。奥陶纪生物大辐射后全球遗迹属增长至117种, 形态构筑类型增加到40种, 其中生物侵蚀构造多样性从5种增加至25种, 形态构筑类型从4种增加至14种; 奥陶纪生物大辐射期间全球遗迹属和形态构筑类型的增量主要由深海遗迹化石类群和生物侵蚀构造贡献。白垩纪是中生代生物大辐射的尾声, 浅海遗迹多样性一直保持高值, 直至大灭绝前开始降低; 深海遗迹多样性在白垩纪阿尔必期后开始大幅增长, 一直持续到新生代渐新世才趋于平缓。
生物大辐射过程中遗迹化石形态构筑类型出现多样化和复杂化, 同类生物的行为习性发生优化, 形态、构筑和功能逐渐缜密、集约、系统、高效, 觅食格局清晰而规则, 体现出造迹生物形态功能的进化。寒武纪, 潜穴的分支增多, 深度增大, 出现了如Cruziana的内生水平潜穴; 前寒武纪就存在的遗迹属Treptichnus在寒武纪演化出更多遗迹种, 化石数量增加, 形态更为多样; 雕画迹首次“ 登场” , 反映了造迹生物对沉积物中有机质利用方式的进步和优化。奥陶纪, 形态结构复杂的雕画迹进一步繁盛, 造迹生物出现了创新性行为, 如仅仅分布于奥陶纪的“ J” 型遗迹属Daedalus, 始于寒武纪的遗迹属Rosselia出现了向上运动的逃逸行为, 三维分支潜穴Thalassinoides先后出现简单迷宫状、复杂箱状2种形态, Oldhamia出现了生物行为优化等等。到了白垩纪, 近岸浅海遗迹化石深受几次大灭绝事件的影响始终未能有突破性的发展, 但复杂深海遗迹化石的多样性达到了显生宙的巅峰, Nereites和Zoophycos先后出现了生物行为优化的现象。
随着生物大辐射进程发展, 造迹生物觅食策略更复杂多样。在埃迪卡拉纪, 微生物席底质广泛分布, 生物觅食策略包含食悬浮物、食碎屑物、食沉积物, 但均未能深入沉积物内部。
寒武纪, 遗迹歧异度增加, 以生物出现垂向掘穴行为为特征, 实现了从二维平面到三维立体空间的突破, 最大生物扰动指数可达6, 挖掘深度大大增加。这一时期底栖生物觅食策略为食沉积物、食碎屑物、食悬浮物、耕作相结合的方式, 并且突破了仅在沉积物表层觅食的限制。奥陶纪, 生物遗迹开始辐射至深海, 深海雕画迹的增多表明食沉积物觅食策略的地位上升, 复杂的耕作行为增多。同时, 生物钻孔大幅增加, 一方面源于大量出现的食肉动物的捕食行为, 另一方面生物对沉积物和其他底质的侵蚀能力显著增强; 放射虫、笔石、有孔虫等浮游生物大量出现, 水柱中颗粒有机质大量增加, 使得食悬浮物者在这一时期大幅增加。奥陶纪海洋底栖动物的觅食策略受环境和环境内动物群类型控制明显, 近岸浅海环境以食悬浮物、主动捕食为主, 深海、半深海、远洋环境则以食沉积物、食碎屑物、耕作更占优势。白垩纪深海雕画迹的大量出现说明了食沉积物者和生物耕作行为的主导地位。此时造迹生物已扩散至非海洋环境, 浅海造迹生物既面临捕食者压力, 又面临抢夺生存资源的竞争压力, 浅海环境觅食策略完备性提高, 食沉积物、食悬浮物、主动捕食等多种觅食方式相结合, 生物生存的策略性和集约性更强。深海环境则仍然以食悬浮物、食碎屑物和耕作为主导。
遗迹化石的空间分布范围扩大, 体现出造迹生物定居范围扩张, 阶层结构复杂化。前寒武纪— 寒武纪之交, 造迹生物的生态空间出现从二维到三维的突破, 底栖生物由仅生存在沉积物表层和浅层扩展至沉积物内部, 并表现出向陆表海、半咸水和深海多个方向迁移的趋势。生物行为直接导致寒武纪底栖生态环境显著改变, 食悬浮物者出现形成“ 农艺革命” , 建立了类似现代沉积物的分层结构, 生物扰动程度增强。因生物扰动增强形成的“ 寒武纪底质革命” , 使得微生物席底逐渐转变为混合底(Má ngano and Buatois, 2017)。
从寒武纪到奥陶纪, 浅海环境在奥陶纪生物大辐射期间被全面开发, 同时造迹生物开始向深海迁移, 这是显生宙第1次大规模底栖造迹生物迁移。复杂精美的雕画迹在深海出现, 是造迹生物占领新生存空间的一次成功尝试。奥陶纪生物大辐射期间浮游生物的大量出现, 表明海洋生态结构立体化, 几乎占据了整个水柱(Servais et al., 2008, 2010), 食悬浮物者增强了底栖生物与水体之间的物质交换, 底栖动物对海洋生态空间有机质的利用变得更为全面和多元。
白垩纪以前, 造迹生物就占领了淡水和陆地。白垩纪浅海的垂向潜穴加强了底栖生物与水体有机质的交互, 浅、中、深层潜穴共存(Ł aska et al., 2017)。深海遗迹化石群落繁盛, 丰富的深海雕画迹体现了生物对深海沉积物中有机质的充分利用和沉积物表层复杂的耕作行为。不同阶层的底栖生物互相牵制影响, 生态结构复杂。
由于遗迹化石指代的是生物和不同底质间相互作用的方式, 反映生物的行为习性(Buatois and Má ngano, 2011), 因此, 虽然生物大辐射期间遗迹化石和实体化石的多样性都快速增加, 但二者地质意义不能完全等同。通常认为, 生物大辐射事件源于获取生态机会, 包含4种途径: (1)出现新资源, 减少相似种之间的竞争; (2)原始优势物种减少或灭绝时, 抢占复苏期的原始生态位; (3)形成关键创新性演化, 使生物具备新能力, 利用曾经无法利用的资源; (4)殖居于未被利用的生态位(Stroud and Losos, 2016)。这4种方法对形成遗迹化石响应模式的影响价值不同。
新资源使得相似种之间的竞争关系减弱, 无法迫使生物形成获取资源的新方法。简言之, 其原理在于生成更多的养料来供给更多的生物, 但生物汲取养料的方式与此前类似。它常是生物大辐射的必要条件之一, 或是辐射形成的多米诺效应的结果, 但不是影响全球生物遗迹面貌的主控因素。
原住民在灭绝后大幅消亡, 可形成空缺生态位, 其后的复苏过程确实可以被看作一次辐射事件, 旨在生物多样性和生态系统恢复至灭绝前水平。但这一生态机会对遗迹化石多样性的意义相对较低, 根本原因是遗迹化石本身“ 多物一迹和一物多迹” 的特点使得大规模“ 灭绝” 在属级尺度很难真正发生, 仅有少量遗迹属真正灭绝了, 如Oldhamia。遗迹化石表现为在局限的时间段中遗迹多样性和歧异度降低, 在复苏期后又会重新出现, 其他指标如丰度、潜穴深度、直径、生态分布范围也能逐渐恢复至灭绝前水平。因此, 灭绝形成空缺生态位无法刺激全球遗迹化石面貌出现里程碑式变化。
生物形成关键创新性演化对于改变全球生物遗迹面貌的意义较大。创新性演化是指生物演化出新器官或组织(Stroud and Losos, 2016), 如鱼类在志留纪早期形成了颌。新器官或组织使生物得以完成新动作, 或具备更精密准确的行为, 如寒武纪生物的运动能力和感知器官进化, 导致寒武纪信息革命发生(Plotnick, 2007; Plotnick et al., 2010), 出现硬壳以在生物壳体形成钻孔(Kobluk et al., 2016)。奥陶纪底栖生物幼虫的游泳能力增强, 使得遗迹化石生态分布扩散至深海。因形成雕画迹的生物行为还是一种推测, 这一生态机会对白垩纪遗迹化石面貌的影响意义尚不明确。中生代海洋革命事件后, 激烈的竞争关系使得整个海洋底栖群落大规模重组, 海洋空间已经被充分占领(水平和垂向尺度), 内生动物分层结构的复杂性和捕食强度都大幅增加。雕画迹在此时繁盛(Uchman and Wetzel, 2012), 同属生物遗迹的行为优化现象, 高超的趋避技巧, 也在此时出现。推测可能是生物形成了更敏锐的感知能力, 更智能, 可以完成更精密的动作, 即形成了创新性演化, 如同现代生物的群集行为, 种群内的个体能够感知周围环境信息, 进行信息的交换和共享, 互相趋避, 是一种高度智能化的行为。但雕画迹本身的行为意义还是一种推测, 关于其成因是否受益于创新性演化, 还有待进一步研究。
生物占据空缺生态位是改变全球生物遗迹面貌、促进生物遗迹发展的主要原因, 它的影响在寒武纪和奥陶纪的生物遗迹响应中显得尤为重要。前寒武纪已有生命痕迹, 但生物群落还未形成足够规模, 且集中在滨海、浅海的沉积物表面。随着雪球地球融化, 生态环境改善, 寒武纪的海洋存在空缺生态位, 且有机质、氧气含量充足, 浅海环境内生物群落得以迅速发展(Zhang and Shu, 2014)。对空缺生态位的开发表现为浅海生物遗迹多样性和歧异度显著增加, 第二世遗迹化石在出现垂向分量的基础上, 出现了垂直和“ U” 型形态, 即生物开始向沉积物深部掘穴。必须强调的是, 空缺生态位并不是形成这一响应面貌的唯一“ 功臣” , 构造作用、雪球地球融化使输入海洋的可利用养分增多, 含氧量增加、钙含量增加、初级生产力提升等是促成此面貌的必需环境因素(Zhang and Shu, 2014)。生物的垂向掘穴, 破坏了原本坚实致密的微生物席底, 更多沉积物中保存的有机质随着水柱输送到海洋中, 增强了营养物质交换(Má ngano and Buatois, 2017)。即在占领生态位的过程中能动地创造了新资源, 形成正反馈, 指向了前文提到的第1种获取生态机会方法。
继寒武纪首次出现生物向深海迁移的趋势后, 这一趋势在奥陶纪被加强和放大。生物占据浅海生态空间, 但深海仍然未被完全开发, 存在明显空缺生态位, 造迹生物开始向远洋深海探索。与此同时, 奥陶纪古板块向赤道聚合, 具备显生宙最广泛的热带— 亚热带大陆架。奥陶纪频繁的火山活动一方面使海侵和海平面上升加剧, 另一方面形成了大量短时存在但具有众多群岛的边缘盆地, 浅海和深海都出现了新的生态位(Barnes, 2004; Servais et al., 2010), 表现为浅海遗迹多样性和歧异度进一步增加, 深海遗迹得以蓬勃发展, 大量此前仅在浅海分布的遗迹化石在早奥陶世弗洛期开始向深海扩散。且海洋垂向生态结构复杂化、觅食策略多样、生物扰动强度增加, 都能够说明生物开发新生态位的过程。无独有偶, 和寒武纪一样, 空缺生态位是主控致因却不是唯一致因, 地幔柱事件使得CO2含量升高形成温室效应, 温度升高使初级生产力提升, 为海洋充入新资源(Servais et al., 2010)。深海雕画迹出现和生物行为的集约性, 反应了生物自身的感知和运动能力增强(Uchman, 2003)。这些条件协助形成了奥陶纪遗迹化石面貌, 也再次印证了生物宏演化规律和适宜的环境背景这2个方面的条件在生物辐射中缺一不可(詹仁斌等, 2013)。
目前大量的研究都强调外因对生物行为的影响, 诚然, 环境和生物表型的变化是最直接能够为人们所观察到的现象, 因此, 外因在辐射进程中重要性常常更容易被关注。随着基因组学和分子钟研究的深入, 生物内因对辐射的调控也逐渐被人们关注。如对于寒武纪生物基因组的研究, 能够说明在寒武纪生命大爆发之前, 基因组已经具备一定的调控能力(Zhang and Shu, 2014)。虽然化石数据库相对滞后于分子钟的推算结果, 但对于大部分具有完整骨骼的类群来说, 分子钟证据和化石证据都基本能够相互印证(Erwin and Tweedt, 2011; Erwin, 2020b)。有趣的是, 在对于物种起源的研究中, 分子钟推算的后生动物起源时间和现有化石数据库相去甚远, 时间差甚至超过了200 Ma(Erwin, 2020b), 甚至更多。这一争论在遗迹化石面貌中表现为生物某种创新性运动能力的出现时间和原因, 因缺乏更多的证据和高精度统计, 目前更像是一场关于“ 先有鸡还是先有蛋” 的辩论。
总体而言, 生态机会的出现仍是被广泛承认的形成遗迹化石辐射面貌的重要致因, 同时辐射进程也是一个致因, 有主有次、多重因素交织影响而成的复杂过程。
1)海相遗迹化石对生物大辐射进程的响应包括多样性和歧异度、造迹生物形态功能、造迹生物觅食策略和生态分布4个方面, 表现为多样性和歧异度显著增加、生物行为集约化、底栖生物的觅食策略和行为习性更为多样、遗迹化石的空间分布范围扩大和阶层结构逐渐复杂化。
2)遗迹化石对显生宙3个主要生物大辐射过程的响应模式各有特点: 寒武纪遗迹化石出现垂向构造, 突破二维平面结构变为三维; 奥陶纪遗迹化石向深海迁移, 且形态更加复杂、规则; 白垩纪深海雕画迹多样性大幅增长。
3)对辐射事件的响应致因源于获取生态机会的过程, 其中对遗迹化石面貌影响最大的是在未被利用的生境定居的过程, 其次是造迹生物本身形成关键创新型演化。除此以外, 还有发生灭绝事件和出现新资源这2种方式, 但并不是影响遗迹化石面貌的主控因素, 通常是辅助因素或多米诺效应的结果。
致谢 感谢在鄂尔多斯盆地西缘野外工作和遗迹化石采集过程中中国地质大学(武汉)DIG团队老师和同学的大力支持!
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|