


{kind=link}
{kind=link}
{kind=link}
{kind=link}
华南埃迪卡拉纪—寒武纪宏体藻类化石的时空分布与多样性变化*
[李高榕1  , 魏凡
, 魏凡1, 2 , 丛培允1, 2 ]
, 魏凡, 丛培允|
|

第一作者简介 李高榕,男,1996年生,云南大学古生物研究院博士研究生。E-mail: ligaorong@mail.ynu.edu.cn。
华南地区众多埃迪卡拉纪—寒武纪特异埋藏化石库为解读生命和生态系统的演化提供了重要化石证据,其中宏体藻类产出数量多、分布广、形态多样,是该时期生态系统中不可忽视的组成部分。过去对华南宏体藻类的研究多集中于形态、分类和古生态方面,对于其不同时期的地理分布和多样性变化较少涉及。文中对华南埃迪卡拉纪—寒武纪特异埋藏化石库中的宏体藻类进行了总结和分析,结果发现:(1)至晚埃迪卡拉世,宏体藻类的古地理分布范围和属、种级多样性明显扩大和提高,在晚埃迪卡拉世第五期达到顶峰; (2)寒武纪纽芬兰世宏体藻类的地理分布范围和属、种级多样性均较低,至第二世则明显扩大和增加; (3)苗岭世之后,宏体藻类古地理分布大幅度缩小,但多样性显著升高并达到第二高峰,直到芙蓉世多样性又急剧下降。推测区域海水含氧量的增加可能是导致华南宏体藻类繁盛(属、种多样性高)的主要原因,适合宏体藻类保存的泥页岩的分布范围或出露面积则可能是影响宏体藻类古地理分布范围的重要因素。
About the first author LI Gaorong,born in 1996,is a Ph.D. candidate at Institute of Palaeontology,Yunnan University. E-mail: ligaorong@mail.ynu.edu.cn.
Numerous Ediacaran-Cambrian Lagerstätten in South China provide important fossil evidence for the understanding of early evolution of life and ecosystems. Macroalgae were abundant,widely distributed,diverse in morphology,and constituted an essential part of ecosystems during the Ediacaran-Cambrian. Previous investigations on the Ediacaran-Cambrian macroalgae in South China focused on morphology,classification and palaeoecology,while their geographical distribution and diversity changes have seldom been studied in detail. In this paper,we summarize the Ediacaran-Cambrian macroalgae from the Ediacaran-Cambrian Lagerstätten of South China. The palaeogeographic distribution and genus/species diversity of macroalgae increased significantly from early to late Ediacaran,and reached the peak in the Age 5. The palaeogeographic distribution and genus/species diversity were extremely low during the Terreneuvian(Cambrian),and increased during the Epoch 2. Thereafter,the palaeogeographic range of macroalgae decreased in the Miaolingian-Furongian,and the genus/species diversity increased significantly and reached the second peak. The increase of regional marine oxygen level,which is suggested to be closely related with the availability of nutrient elements such as N and P,may be the main reason for the flourishing of macroalgae(high genus and species diversity),and the wide distribution of mud shale suitable for the fossilization of soft-bodied organisms might be an important factor controlling the geographic extension of the Ediacaran-Cambrian macroalgae in South China.
藻类是生命演化史早期就已出现的主要生物类群之一, 也是最早出现的真核生物类型(Miao et al., 2019; 张凤廉等, 2021)。在真核生物出现(约16亿年前)之后的10亿年间, 藻类主要以微体化石的形式保存(袁训来等, 2023), 虽然部分地层中报道过宏体藻类化石(Han and Runnegar, 1992; Zhu et al., 2016), 但是因保存较差、数量较少且产出地层年代不清晰, 故而存在颇多争议(张凤廉等, 2021)。进入埃迪卡拉纪之后, 出现了多个以宏体藻类为主的特异埋藏化石库, 例如蓝田生物群、庙河生物群、瓮会生物群等, 宏体藻类的地理分布范围相比之前也有显著提高(陈孟莪和萧宗正, 1992; Yuan et al., 1999, 2011; Xiao et al., 2002; 王约等, 2007; 杜汝霖, 2017; Ye et al., 2019)。迄今, 在全国13个省(市)的埃迪卡拉系中已经发现20余个化石点产出宏体藻类化石(刘傲然, 2018), 其中华南地区的湖北、湖南、贵州、云南东部和安徽南部等地产出尤为丰富。寒武纪, 特异埋藏宏体藻类也产出丰富并广泛分布, 岩家河生物群、牛蹄塘生物群、澄江生物群、杷榔生物群和凯里生物群等化石库均保存了数量可观的宏体藻类化石(杨瑞东, 2006; 郭俊峰等, 2010, 2017; 伍孟银等, 2013; 傅晓平等, 2014; Hou et al., 2017)。另外, 相比埃迪卡拉纪之前, 全球埃迪卡拉纪宏体藻类的形态多样性也出现了大幅度增长, 在华南表现特别明显(丁莲芳等, 1996; Wang et al., 2016; 袁训来等, 2016; Ye et al., 2019)。埃迪卡拉纪, 枝状、扇状、丛状等多种形态的宏体藻类开始大量出现, 细胞功能分化更为明显, 很多化石的保存特征也开始能区分出主要藻类的门类, 这显示了埃迪卡拉纪宏体藻类在多样性方面出现了巨大变化(杨瑞东, 2006)。至寒武纪, 以澄江生物群和凯里生物群为代表的特异埋藏化石库中的宏体藻类属、种丰富, 并出现了确切的钙质宏体藻类(杨瑞东等, 1999b; Yang, 2000; Hou et al., 2017)。由此可见, 华南埃迪卡拉纪— 寒武纪的宏体藻类具有数量多、分布广、形态多样以及保存地层较为连续等特点, 为研究该时期宏体藻类的古地理分布和多样性变化提供了绝佳的材料。
埃迪卡拉纪— 寒武纪是元古宙和显生宙的转折, 是地球生命演化的关键时期, 也是生态系统演化的重要时期。宏体藻类是该时期主要的生命形式之一, 也是生态系统中不可或缺的重要一环。华南保存了埃迪卡拉纪— 寒武纪诸多特异埋藏化石群, 是研究生态系统演化的重要地区。藻类作为这些特异埋藏化石群落中的一员, 无疑是认识埃迪卡拉纪— 寒武纪之交这一重大地质转折期生命或生态系统演化的重要化石证据。前人的研究主要集中于这一时期特异埋藏保存生物群中宏体藻类的形态、分类和古生态(Xiao et al., 2002; Xiao and Dong, 2006; 王约等, 2007; Yuan et al., 2011, 2013; Ye et al., 2019), 对宏体藻类整体的古地理分布和多样性变化等涉及较少。文中全面总结了华南埃迪卡拉纪— 寒武纪特异埋藏宏体藻类化石的古地理分布和多样性特征, 并讨论了该时期宏体藻类的时空分布和多样性变化历程。
宏体藻类是指肉眼可见的真核藻类, 多为多细胞或多核细胞生物, 大小通常大于1 mm(Xiao and Dong, 2006)。与埃迪卡拉纪— 寒武纪后生动物相比, 宏体藻类的研究程度相对较低, 在分类方面尤为明显。后生动物化石具有复杂的解剖构型和多样化的保存形式, 但宏体藻类因缺乏硬体结构而完整保存的相对较少, 在埋藏过程中容易受到破坏, 所保存的生物学信息极为有限, 从而难以辨识。现生藻类通常以藻类的形态、色素成分与含量、光合作用产物、生殖器官与生殖方式、生活史等方面进行分类(刘涛, 2017), 但上述大多数可用来作为分类的特征极少能被石化和保存下来, 造成多数情况下藻类的形态是藻类化石分类最重要也是唯一可以仔细考量的依据, 从而对藻类化石的精确分类位置产生极大影响。同时, 遗迹化石(如粪便化石)等部分其他类型化石因外形同藻类相似, 也容易与藻类化石混淆(Steiner et al., 2005; Babcock et al., 2012)。正是由于现生藻类分类特征在化石中的缺失, 化石藻类的分类系统一直存在较大争议, 因此现今主要包括以现生藻类分类系统和形态为基础的2种分类方法(丁莲芳等, 1996; Xiao and Dong, 2006; 杨瑞东, 2006; Loduca et al., 2017)。这2种分类方法均集中在对藻类属、种的划定方面, 对科及以上更高阶的分类单元较少涉及。为了简要地表示藻类化石的不同形态特征, 文中主要依据杨瑞东(2006)提出的以形态为主的分类观点— — 化石若保存有同现生藻类特征相对应的结构, 则将其归入现生藻类分类系统, 同时也参考了Xiao和Dong(2006)、Loduca等(2017)提出的分类观点。
笔者对华南埃迪卡拉纪— 寒武纪已知的宏体藻类化石记录进行了统计(表1)。为了准确反映埃迪卡拉纪— 寒武纪宏体藻类在华南的时空分布和多样性变化, 文中遵循以下原则: (1)尽可能以属、种为研究对象来探讨宏体藻类的多样性变化; (2)只引用有明确照片和描述的宏体藻类化石数据, 且多名学者对其都进行过引用; (3)只引用有明确地质年代的化石数据; (4)为了方便讨论宏体藻类的时空分布, 以“ 点” 来表示因地图比例限制而无法在地图中识别出来的某个区域的多个剖面。
| 表1 华南埃迪卡拉纪— 寒武纪宏体藻类统计 Table1 Statistics of the Ediacaran-Cambrian macroalgae from the fossil record of South China |
目前, 华南埃迪卡拉纪— 寒武纪的地层划分仍处于研究之中。对于埃迪卡拉系, Narbonne等(2012)、Xiao等(2016)和周传明等(2019, 2021)均对其进行过探讨, 文中采用周传明等(2019, 2021)的观点作为划分依据。该观点主要基于对华南埃迪卡拉纪生物演化进程的认识, 综合化学地层学、事件地层学和同位素地质年代学成果, 将埃迪卡拉系划分为上、下2个统和6个阶。对于寒武系, 笔者参照国际地层年代表4统10阶的划分标准和朱茂炎等(2021)所总结的岩石地层进行划分。
据笔者统计, 华南埃迪卡拉纪— 寒武纪产出的特异埋藏保存化石群共计24个, 其中含有宏体藻类化石记录的共18个, 涉及安徽、湖南、贵州、陕西和云南等地, 共报道宏体藻类化石61属111种(包含17个未定种)(表1)。这充分表明宏体藻类在埃迪卡拉纪— 寒武纪已相当繁盛, 华南化石点呈区域聚集分布的情况较为普遍。虽然在统计中还存在少数未定种, 但是已能够反映出华南埃迪卡拉纪— 寒武纪宏体藻类的基本时空分布特征。
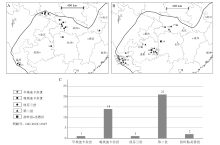
1)早埃迪卡拉世。共发现2处特异埋藏保存的化石库(蓝田生物群和瓮安生物群), 分别产于蓝田组下部和陡山沱组下部, 但只有蓝田生物群中产出确定的宏体藻类化石, 瓮安生物群中的藻类化石基本为磷酸盐化的微体藻类, 目前还未有确切的宏体藻类化石发现(袁训来等, 2002, 2016; Yuan et al., 2011; Xiao et al., 2014)。蓝田生物群主要产于安徽南部休宁县蓝田镇, 化石分布较为集中(图1-A, 1-C)。早埃迪卡拉世为Marinoan冰期之后的第1个地质时段, 此时海平面开始上升, 海洋中生命所需的营养元素也开始急剧增加, 这为宏体藻类的辐射和繁盛提供了适宜的条件(McFadden et al., 2008; Jiang et al., 2011; Knoll, 2017; 张凤廉等, 2021)。
 | 图1 华南埃迪卡拉纪— 寒武纪宏体藻类古地理分布 A— 埃迪卡拉纪宏体藻类分布; B— 寒武纪宏体藻类分布; C— 不同时期化石点数量统计图剖面位置编号: 1— 安徽休宁蓝田; 2— 湖北秭归庙河; 3— 秭归芝麻坪; 4— 兴山麻溪; 5— 神农架林区三里荒; 6— 秭归泗溪; 7— 秭归雾河; 8— 湖南桃映理公港; 9— 贵州江口瓮会; 10— 陕西宁强胡家坝; 11— 勉县鹁鸽洞; 12— 宁强宽川铺; 13— 宁强高家山; 14— 云南晋宁王家湾; 15— 玉溪江川; 16— 湖北秭归岩家河; 17— 湖北秭归罗家沟; 18— 宜昌长阳; 19— 京山石龙; 20— 贵州金沙岩孔; 21— 遵义松林; 22— 黔东南麻江; 23— 台江革一; 24— 剑河八郎; 25— 剑河交榜; 26— 镇远江古; 27— 云南安宁山口; 28— 昆明海口; 29— 昆明二街; 30— 昆明呈贡; 31— 玉溪澄江; 32— 昆明宜良; 33— 昆明高楼房; 34— 马龙乌龙箐; 35— 会泽大海; 36— 昆明岗头村; 37— 武定石将军; 38— 贵州剑河八郎; 39— 湖南花垣排碧村Fig.1 Palaeogeographic distribution of the Ediacaran-Cambrian macroalgae in South China |
2)晚埃迪卡拉世。共发现6个特异埋藏保存的化石库, 分别是庙河生物群、瓮会生物群、武陵山生物群、高家山生物群、石板滩生物群和江川生物群, 均产出确定的宏体藻类化石(陈孝红等, 1999; 华洪等, 2000; Xiao et al., 2002; 唐烽等, 2007; 王约等, 2007; 杨凡等, 2018)。主要分布于湖南西部、湖北西部、陕西南部、贵州东部以及云南东部, 目前已发现14个化石保存点, 且在湖北西部和陕西南部较为集中(图1-A, 1-C)。相比于早埃迪卡拉世, 晚埃迪卡拉世宏体藻类的分布范围明显扩大, 向华南地区西部扩散。
3)寒武纪纽芬兰世。共发现4个特异埋藏保存的化石库, 分别是岩家河生物群、梅树村小壳化石群、西乡生物群和宽川铺生物群。仅有岩家河生物群保存有确定的宏体藻类化石, 其主要分布于湖北省秭归市三斗坪(图1-B, 1-C)(罗惠麟等, 1985; 郑曦, 2010; 郭俊峰等, 2010, 2017; Liu et al., 2016)。其余3个化石库以微体化石为主, 暂无确凿的宏体藻类化石发现。同前一阶段相比, 宏体藻类的分布范围出现明显收缩。
4)寒武纪第二世。这一时期共有11个特异埋藏保存的化石库被报道, 分别是牛蹄塘生物群、澄江生物群、水井沱生物群、清江生物群、荷塘海绵生物群、石牌生物群、关山生物群、马龙动物群、杷榔生物群、剑河生物群及石龙洞生物群, 除荷塘生物群、石牌生物群和马龙动物群之外, 其余8个化石库均有确切的宏体藻类化石产出(Chen and Erdtmann, 1991; 杨瑞东等, 1999a; Chen et al., 2004; 罗惠麟等, 2008; 傅晓平等, 2010, 2014; Hu et al., 2010; 王平丽等, 2013; Hou et al., 2017; 刘璠等, 2017; Fu et al., 2019; Zhang et al., 2021; 赵小云, 2021; 潘晓强等, 2022)。这些特异埋藏化石库主要分布于云南东部、贵州东部和湖北西部, 共计发现21个化石保存点(图1-B, 1-C), 且在云南东部和贵州东部尤为集中。该时期华南宏体藻类的化石点从数量上看达到了最高, 但是晚埃迪卡拉世分布于陕南和湘西的宏体藻类在该时期同区域内并未发现, 因此总体分布范围相比于晚埃迪卡拉世略微收缩。
5)寒武纪苗岭世和芙蓉世。仅凯里生物群和花桥组2个特异埋藏保存的化石库被发现。它们均产出宏体藻类化石, 主要分布于贵州东部和湖南西部的2个化石点(图1-B, 1-C)(杨瑞东, 2006; Babcock et al., 2012), 化石分布范围出现了较大幅度的萎缩。
虽然宏体藻类化石在埃迪卡拉纪之前的地层中已有报道, 并在少数地点(例如河北怀来、蓟县和安徽淮南等地)呈聚集性产出, 但整体来看, 埃迪卡拉纪之前宏体藻类的形态以简单叶状体为优势形态类型, 较少出现分枝现象, 形态分异度较低, 主要类群以Chuaria、Tawuia、Grypania和Longfengshania等属为主(杜汝霖和田立富, 1982, 1985; 符俊辉, 1989; 牛绍武和朱士兴, 2002; 刘傲然, 2018)。同时, 由于化石解剖学证据不足或生物学特征不明确, 所以该时期多数宏体化石的生物学属性还存在争议(张凤廉等, 2021)。值得注意的是, 近年来在成冰纪地层中所报道的宋洛生物群不仅包括形态简单的宏体藻类(如Chuaria和Tawuia), 还发现部分属、种(如Konglingiphyton和Enteromorphites)具有固着器和分枝等形态特征(Ye et al., 2015), 这说明具有复杂形态和高度多样性的藻类出现时间可能早于埃迪卡拉纪。相比于埃迪卡拉纪之前, 埃迪卡拉纪— 寒武纪宏体藻类的整体形态结构变得更为复杂, 叶状体、拟茎和固着器等组织分化更为明显, 物种多样性和形态分异度都出现了较大幅度的增加(Xiao et al., 2002; Xiao and Dong, 2006; 王约等, 2007; Yuan et al., 2011), 已经具有了现生藻类的整体形态面貌。在埃迪卡拉纪之前占据优势的宏体藻类, 如Chuaria和Tawuia, 在该时期已被线状、分枝状和扇状藻类所取代, 如Vendotaenia、Sinocylindra、Doushantuophyton、Enteromophites和Flabellophyton等(图2)。在埃迪卡拉纪— 寒武纪, 宏体藻类虽大量发育, 但不同时期宏体藻类的多样性和形态面貌还是存在显著差异。
 | 图2 埃迪卡拉纪— 寒武纪代表性宏体藻类化石 A— Chuaria circularis; B— Sinocylindra yunnanensis, YKLP15252; C— Vendotaenia; D— Huangshanophyton fluticulosum; E— Longfengshania cordata; F— Doushantuophyton lineare; G— Marpolia spissa; H— Longifuniculum dissolutum; I— Enteromorphites siniansis。比例尺分别为5 mm(H), 3 mm(A), 2 mm(B, D, E, G, I), 1 mm(F)。其中A, F引自Ye等(2019), C引自郭俊峰等(2010), D, I引自袁训来等(2016), E引自 唐烽等(2006), G引自赵小云(2021), H引自伍孟银等(2013)Fig.2 Representative macroalgae types from the Ediacaran-Cambrian |
4.2.1 埃迪卡拉纪宏体藻类的多样性
早埃迪卡拉世, 仅有蓝田生物群中产出8属16种宏体藻类化石(图3; 图4), 以Flabellophyton lantianensis为主, Chuaria的产出也较为丰富, 还产出Doushantuophyton、Enteromphites和Morpolia等呈分枝状和丛枝的宏体藻类(袁训来等, 2002, 2016; Yuan et al., 2011, 2013)(图2)。蓝田生物群不仅具有Chuaria这样形态简单的藻类类群(该属也见于宋洛生物群), 还具有Flabellophyton和Enteromphites这样结构复杂的藻类属、种, 反映该时期宏体藻类开始出现由简单形态逐渐向复杂形态演化。
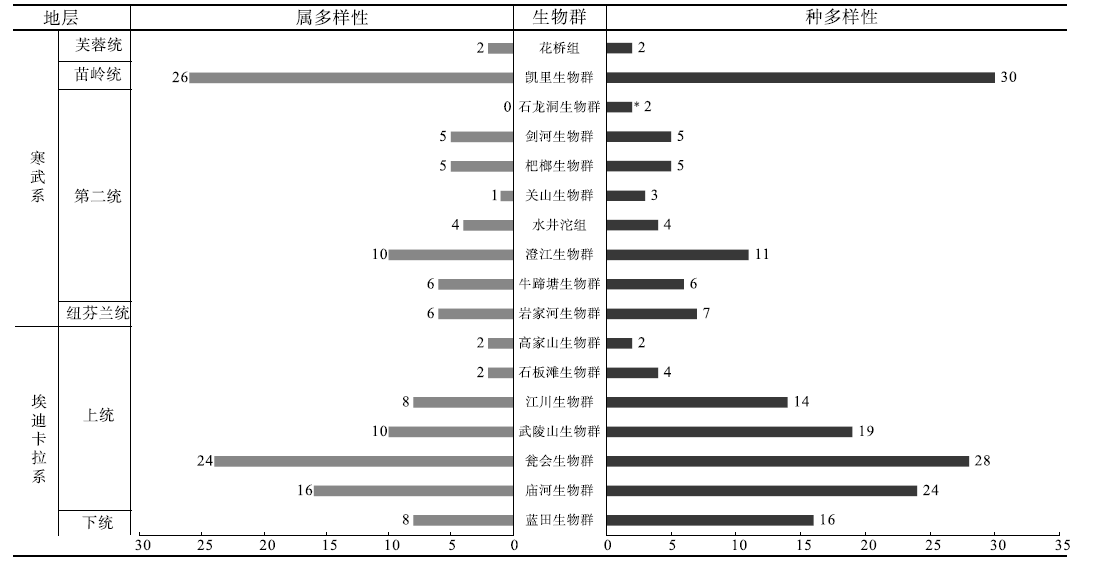 | 图3 华南埃迪卡拉纪— 寒武纪各特异埋藏保存生物群中宏体藻类的多样性(属和种)统计图 * 处为暂未确定属种, 但已经发现不同形态的宏体藻类化石Fig.3 Diversity of macroalgae from the Ediacaran-Cambrian Lagerstä tten of South China |
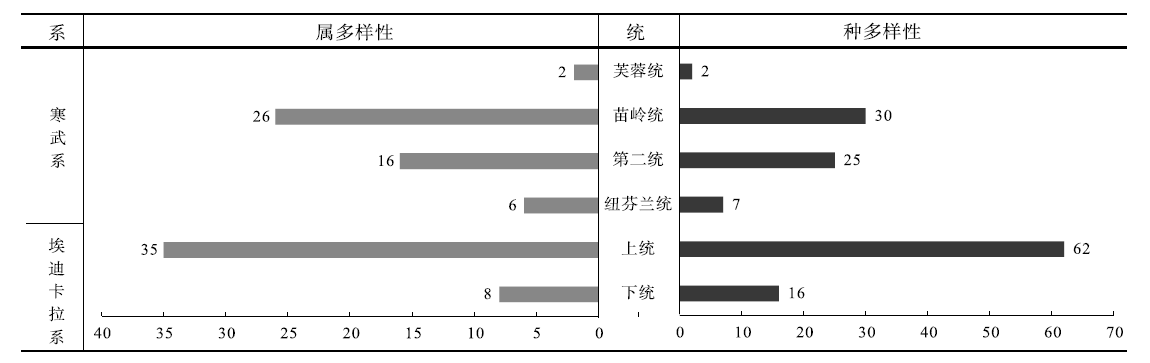 | 图4 华南埃迪卡拉纪— 寒武纪不同时段地层(统)中特异埋藏保存宏体藻类的多样性(属和种)统计图Fig.4 Diversity of macroalgae from different intervals of the Ediacaran-Cambrian in South China |
晚埃迪卡拉世, 共有6个特异埋藏化石库产出宏体藻类化石: 庙河生物群产出藻类化石16属24种, 瓮会生物群产出藻类化石24属28种, 武陵山生物群产出藻类化石10属19种, 江川生物群产出藻类8属14种, 高家山生物群产出藻类化石2属2种, 石板滩生物群产出藻类化石2属4种(图2)(陈孝红等, 1999; 华洪等, 2000; 陈孝红和汪啸风, 2002; Xiao et al., 2002; 袁训来等, 2002; 唐烽等, 2007; 王约等, 2007; Wang et al., 2015; 杨凡等, 2018; Ye et al., 2019)。该时期宏体藻类共产出35属62种(图3):庙河生物群和瓮会生物群宏体藻类化石产自陡山沱组上部, 产出年代大致相当, 暂时划归晚埃迪卡拉世第五期, 共产出宏体藻类24属32种(叶琴, 2018; 王野, 2019; 周传明等, 2021)。武陵山生物群产自留茶坡组中下部, 其余化石库均产自灯影组上部, 虽然留茶坡组和灯影组的具体层位对比暂时还不清楚, 但是可以确定的是这些化石库的产出年代均为晚埃迪卡拉世第六期, 共产出藻类17属33种(周传明等, 2021)。在晚埃迪卡拉世第五期, 宏体藻类整体多样性远高于早埃迪卡拉世。在此之前繁盛的形态简单的Chuaria等藻类业已被形态分异度高的线状、分枝状和丛状藻类所取代, 同时多数藻类的组织分化更为明显和复杂, 例如Baculiphyca和Gemmaphyton的固着器上具有明显的须根状结构(Wang et al., 2020)、Miaohephyton的藻体上具有确定的生殖窝(Xiao et al., 1998)、Wenghuiphyton和Globusphyton中存在平卧的匍匐状拟根(王约等, 2007; 王野, 2019)。值得注意的是, 此时的宏体藻类依然延续了早埃迪卡拉世的部分藻类属种, 如Doushantuophyton和Enteromphites(图2)(袁训来等, 2016; 王野, 2019; Ye et al., 2019)。晚埃迪卡拉世第六期的宏体藻类在江川生物群和武陵山生物群中基本延续着第五期以来宏体藻类繁盛的态势, 但是在多样性方面却有明显的降低, 总体以线状、棒状、叶状和条带状等不分枝形态为主, 扇状和丛状藻类的物种多样性远不如第五期。与江川生物群和武陵山生物群对比而言, 高家山生物群和石板滩生物群宏体藻类的多样性更低, 仅有少量属种出现, 且形态单一。总之, 在晚埃迪卡拉世第六期, 华南地区宏体藻类的多样性均出现不同程度降低, 且具有显著的地区差异。
总体而言, 埃迪卡拉纪是宏体藻类快速演化和辐射的时期, 宏体藻类在成冰纪及之前的基础上继续演化, 出现了多种新形态, 具有更为明显的细胞功能分化和复杂的结构构造, 形态分异度和属、种多样性都达到了更高的水平。但是高家山生物群和石板滩生物群中宏体藻类类群的稀少, 可能反映了埃迪卡拉纪末期华南部分地区宏体藻类多样性急剧降低, 或者是该时期能保存软躯体藻类的泥页岩等岩性分布范围急剧减少。
4.2.2 寒武纪宏体藻类的多样性
寒武纪纽芬兰世, 仅岩家河生物群中发现藻类6属7种(图3; 图4), 其产于岩家河组, 主要以带状、分枝状、叶状为主, 如Sinocylindra、Konglingiphyton、Longfengshania等, 具有明显的器官分化(郭俊峰等, 2010, 2017)。该生物群的宏体藻类和产于同一地区埃迪卡拉系上统的石板滩生物群相比, 物种多样性和形态分异度都具有明显提升, 分枝状和具有明显组织分化的藻类又一次出现, 藻类整体面貌具有显著的“ 庙河生物群” 特点(郭俊峰等, 2010)。但与产于滇东和湘西的埃迪卡拉系上统江川生物群和武陵山生物群相比, 宏体藻类的多样性依然较低。
寒武纪第二世, 共有8个特异埋藏保存化石库产出宏体藻类化石: 牛蹄塘生物群产出6属6种; 澄江生物群产出10属11种; 水井沱组(包含水井沱生物群和清江生物群)中可以确定的藻类化石4属4种, 还包含6个暂时无法确定的疑似藻类类群; 关山生物群产出至少3个种; 杷榔生物群产出5属5种; 剑河生物群产出5属5种; 石龙洞生物群产出2个疑似藻类类群(图3)(Chen and Erdtmann, 1991; 杨瑞东等, 1999a; Chen et al., 2004; 罗惠麟等, 2008; 傅晓平等, 2010, 2014; Hu et al., 2010; 王平丽等, 2013; Hou et al., 2017; 刘璠等, 2017; Fu et al., 2019; Zhang et al., 2021; 赵小云, 2021; 潘晓强等, 2022)。迄今报道的藻类化石共计16属25种(图4), 整体形态依然以条带状和线状为主(如Sinocylindra和Yuknessia), 也存在分枝状和丛状形态(如Doushantuophyton和Marpolia)(图2)。相比于寒武纪纽芬兰世, 第二世宏体藻类的多样性已经有了显著提升。但是, 该时期宏体藻类的多样性, 尤其是分枝状、扇状和丛状形态藻类的多样性, 远不及埃迪卡拉纪繁盛。值得注意的是, 该时期澄江生物群中产出的藻类化石的多样性较之其他地区显著偏高。
寒武纪苗岭世和芙蓉世, 共有2个特异埋藏保存化石库中产出宏体藻类化石, 凯里生物群和花桥组中分别产出藻类化石26属30种和2属2种(图3; 图4)(杨瑞东, 2006; 伍孟银等, 2010, 2012, 2013; Babcock et al., 2011; Wu et al., 2011, 2016)。凯里生物群中藻类形态分异度较高, 具有球状、带状、分枝状、丛状、束状等多个形态类型, 部分藻体还出现了确定的节状分支和钙化特征(Yang, 2000; 杨瑞东, 2006; Wu et al., 2011, 2016)。此时宏体藻类在形态分异度和多样性方面重新达到了相对较高的水平, 但是由于苗岭世之后华南地区的化石普遍稀少(朱茂炎等, 2021), 故而花桥组中2属2种的宏体藻类化石可能表明了此时湘西的生态环境并不适宜宏体藻类的生存, 或埋藏环境(岩性)不适宜宏体藻类的保存。
总之, 与埃迪卡拉纪相比, 寒武纪纽芬兰世的特异埋藏宏体藻类化石在物种多样性方面出现了较大萎缩, 形态分异度降低也较为显著。随后从寒武纪第二世开始, 直到苗岭世宏体藻类的物种多样性逐步回升, 但总体多样性还是低于埃迪卡拉纪。
华南埃迪卡拉系中共计产出宏体藻类38属72种。在延续埃迪卡拉纪之前部分宏体藻类属、种(以Chuaria、Grypania和Longfengshania为代表)的基础上, 埃迪卡拉纪宏体藻类演化出棒状、分枝状、丛状、扇状等多种形态类型(例如Anhuiphyton、Baculiphyca、Doushantuophyton和Flabellophyton等), 且部分属、种为埃迪卡拉纪所特有(表1)。该时期具有分枝状、束状、丛状和扇状等分枝形态的宏体藻类19属38种, 具有圆盘状、线状、带状、叶状和棒状等不分枝形态的宏体藻类19属34种。产生于埃迪卡拉纪之前的宏体藻类为5属7种, 呈圆盘状、带状和叶状形态, 显示了埃迪卡拉纪宏体藻类对之前时代宏体藻类属、种的延续性。埃迪卡拉纪之前, 宏体藻类主要以圆盘状和带状为主(叶琴, 2018), 暂时未见分枝状宏体藻类的报道。而埃迪卡拉纪, 分枝状宏体藻类不论属一级还是种一级均达到或超过了50%, 多样性方面整体快速发展。埃迪卡拉纪位于“ 雪球地球” 事件之后, 此时全球氧气增加、气候转暖、海平面上升和海域扩大, 都为宏体藻类的发展提供了充足空间(叶琴, 2018)。同时, 埃迪卡拉纪多种形态宏体藻类的出现与演化, 揭示了当时宏体藻类开始对生存空间与环境因子进行竞争, 如宏体藻类分枝状形态恰恰可以帮助宏体藻类获得充足的阳光、CO2和矿物质, 也是其形态分异的最初表现(袁训来等, 1995)。
华南寒武系共计产出宏体藻类35属51种(表1): 具有分枝状、束状、丛状和扇状等分枝形态宏体藻类17属21种, 其中6属7种为埃迪卡拉纪出现的类群; 具有圆盘状、线状、带状和叶状等不分枝形态的宏体藻类18属30种, 其中5属5种为埃迪卡拉纪出现的类群。相比于埃迪卡拉纪, 宏体藻类在种一级的多样性上有了明显的降低。虽然部分属、种自埃迪卡拉纪延续至寒武纪, 但是更多的属、种是自寒武纪才开始出现化石记录。暂时还无法排除化石鉴定上存在的问题, 例如是否存在同物异名现象, 但是不可否认的是, 寒武纪的宏体藻类与埃迪卡拉纪宏体藻类在属、种面貌上存在极大的不同, 埃迪卡拉纪— 寒武纪之交一系列环境和生物的演化很可能是促使宏体藻类属、种面貌变化的重要原因。
通过对特异埋藏宏体藻类的统计, 可以得知在埃迪卡拉纪— 寒武纪, 除苗岭世外, 其他各个世的宏体藻类的古地理分布范围和多样性呈现出高度相关性, 即古地理分布范围变大(缩小)的同时, 多样性也在提高(降低)。同时, 笔者还发现这些变化同岩相和海水含氧量存在明显相关性。
据统计(表1), 在埃迪卡拉纪— 寒武纪, 除了石板滩生物群外, 其余特异埋藏宏体藻类均产出于粉砂质泥岩、泥页岩和泥灰岩中, 在砾岩、砂岩、灰岩和白云岩等其他岩相中产出极少。其中产自粉砂质泥岩、泥页岩和泥灰岩中的宏体藻类59属107种, 产于灰岩中的宏体藻类仅有2属4种。在华南, 埃迪卡拉系上统、寒武系第二统均是粉砂质泥页岩相和泥灰岩相发育的地层(周传明等, 2021; 朱茂炎等, 2021), 此时特异埋藏宏体藻类地理分布范围均有所提高。埃迪卡拉系下统、寒武系芙蓉统灰岩、白云岩或砂岩相较为发育(周传明等, 2021; 朱茂炎等, 2021), 宏体藻类古地理分布在此时则明显降低。值得注意的是, 在部分区域纽芬兰统和芙蓉统存在大范围缺失, 这也会影响宏体藻类的古地理分布范围。而粉砂质泥岩、泥页岩出露的面积也影响对于宏体藻类古地理分布的判断, 例如苗岭世宏体藻类分布范围的急剧缩小, 可能与当时华南适宜藻类保存的岩相局限分布有关, 这有待于后续研究进行补充。由此可见, 岩相是影响特异埋藏宏体藻类产出的重要因素, 粒度较细的碎屑岩更有利于宏体藻类化石的保存。
大气— 海水含氧量和营养元素的供给被认为可能是藻类演化主要的环境控制因素(Anbar and Knoll, 2002; 张凤廉等, 2021)。海洋中的营养元素, 尤其是氮、磷, 对宏体藻类的分布、生长和繁殖起着显著的控制作用(Anbar and Knoll, 2002; Squire et al., 2006; Planavsky et al., 2010)。在埃迪卡拉纪— 寒武纪, 氮、磷等营养元素也更多地被大气— 海洋氧化程度所控制, 海水含氧量的提高使得营养元素的容量和可利用程度增大, 进而促使宏体藻类进一步演化并提升海洋初级生产力水平(Anbar and Knoll, 2002; Cremonese et al., 2014; Wang et al., 2018; 张凤廉等, 2021)。该时期宏体藻类多样性的变化, 恰恰也表明宏体藻类演化与海水含氧量的变化密切相关。胡修棉等(2001)、胡修棉和王成善(2001)认为碳同位素可以间接判别海洋氧化还原条件的改变, 是定性反映海水中溶解氧含量的手段之一, 还发现了碳同位素负偏移时海水中溶解氧含量往往偏高。Bristow and Kennedy(2008)、Xiao(2008)、Li等(2018)、Chen等(2022)也发现δ 13C的负偏移与海洋氧化事件存在密切关系。同时, 王新强等(2010)还发现埃迪卡拉生物群的发展多出现于δ 13C负偏移前后, 并在每次δ 13C负偏移之后呈现进一步多样化的趋势。笔者发现晚埃迪卡拉世和寒武纪第二世、苗岭世宏体藻类多样性的提高都在δ 13C出现负偏移之后, 例如庙河生物群和瓮会生物群便出现在陡山沱组近顶部碳同位素负偏移(EN3)之后, 寒武纪第二世宏体藻类多样性的提高出现于石岩头组碳同位素负偏移(SHICE)之后, 苗岭世宏体藻类多样性的提升也是在寒武系第二统顶部碳同位素负偏移(ROECE)之后(周传明等, 2019; 朱茂炎等, 2019)。此外, Sahoo等(2016)通过对华南埃迪卡拉— 寒武纪地层中铁元素、硫同位素和氧化还原敏感元素进行分析, 发现635 Ma、580 Ma、560 Ma、530 Ma和522 Ma都发生过海洋氧化事件, 而宏体藻类多样性的提高也与这些海洋氧化事件的发生时期大体一致。综上可知, 海水含氧量波动可能是影响这一时期宏体藻类多样性变化的主要因素, 海水含氧量或氧化水平的提高可能会提高营养元素如氮、磷的可获得性, 进而促进宏体藻类多样性的提高。另一方面, 宏体藻类的快速演化是一种正反馈效应, 会使得光合作用产出氧气的速率显著提高, 从而进一步促进海洋氧化(Campbell and Squire, 2010)。埃迪卡拉纪— 寒武纪宏体藻类多样性及海洋环境变化之间的耦合关系, 提供了生命与环境协同演化的又一案例, 当然这方面尚需开展更深入的研究。
1)统计表明, 华南埃迪卡拉纪宏体藻类的古地理分布和多样性均呈显著扩大和提高趋势, 在晚埃迪卡拉世达到最大, 而多样性在第五期达到顶峰, 在第六期末明显降低。寒武纪纽芬兰世— 第二世, 宏体藻类的古地理分布范围和属、种数量都增大。至苗岭世, 宏体藻类的多样性达到顶峰, 但地理分布范围则急剧缩小。芙蓉世宏体藻类的古地理分布和多样性均急剧降低。
2)除苗岭世外, 宏体藻类的古地理分布范围和多样性在埃迪卡拉纪— 寒武纪呈现出高度相关性, 即古地理分布范围变大(缩小)的同时多样性也显著提高(降低)。苗岭世宏体藻类分布范围的急剧缩小, 可能与当时华南适宜藻类保存的泥页岩相的局限分布有关。不同时期宏体藻类的产出显著受制于岩相, 其多产出于粉砂质泥岩、页岩或泥灰岩相中, 在灰岩和白云岩中产出较少。海水含氧量的升高, 可能是宏体藻类多样性提高的主要影响因素。
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|