



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
甘肃永昌乌拉尔世 Sphenopteris(楔羊齿)新记录: 兼论晚古生代 Sphenopteris多样性及古地理演化*
[王雪莲1  , 熊聪慧
, 熊聪慧2 , 孙柏年2 ]
, 熊聪慧, 孙柏年|
|

第一作者简介 王雪莲,女,1991年生,博士,特别研究助理,研究方向为古植物学和气候环境演变。E-mail: wangxuelian@nieer.ac.cn。
Sphenopteris(楔羊齿属)叶化石在全球晚古生代地层中广泛出现,但该属的系统分类位置及演化特征仍有待深入探讨,而且只有极少数种的表皮及气孔微细构造得到揭示。笔者在甘肃永昌乌拉尔统山西组中发现了2种形态保存较好且具有角质层的楔羊齿,分别是 Sphenopteris yongchangensis sp. nov.(永昌楔羊齿—新种)和 Sphenopteris nystroemii (弱楔羊齿)。前者小羽片呈扇形,基部连合,侧脉分叉1次,气孔作椭圆状且为两面生; 后者小羽片呈梭形且顶端钝圆,侧脉分叉1次并达缘,气孔分布于脉间区。经对比,文中报道的2种楔羊齿可能属皱羊齿目(Lyginopteridales)。此外,统计了 Sphenopteris在晚古生代的269个化石记录,结合该属的形态学、解剖学特征和分布记录,探讨了其多样性演化和古地理分布,结果表明: 宾夕法尼亚亚纪至乌拉尔世, Sphenopteris广泛分布于全球中低纬度地区,呈现出高度变异和分化; 自晚泥盆世至二叠纪, Sphenopteris型叶化石在中国自华南向华北迁移演化,说明其多样性特征不仅与湿润气候密切相关,而且与其古地理分布相耦合。
About the first author WANG Xuelian,born in 1991,is an assistant researcher of paleobotany and paleoenvironments,at Northwest Institute of Eco⁃Environment and Resources,Chinese Academy of Sciences. Her research focuses on paleobotany and climate changes. E-mail: wangxuelian@nieer.ac.cn.
Although Sphenopteris species were widespread in the Late Paleozoic around the world,the systematic position and evolutionary characteristics of this genus still need to be addressed,and microstructures such as epidermis and stomata have only been demonstrated in very limited species. We describe two well-preserved fossil species with microstructures from the Cisuralian Shanxi Formation(Early Permian)of Yongchang,Gansu,China,including Sphenopteris yongchangensis sp. nov. and Sphenopteris nystroemii. Pinnules of S. yongchangensis sp. nov. were fan-shaped,with their base connected,lateral veins bifurcate once,and elliptical stomata are amphistomatic. Pinnules of S. nystroemii were fusiform with round top,lateral veins bifurcate once and reach leaf margin,and its stomata were distributed in interveinal area. It is more reasonable to assign the present two species of Sphenopteris to the order Lyginopteridales by detailed comparative analyses. The diversity and palaeogeographic evolution of Sphenopteris species are then considered,based on 269 occurrence records of the genus from the Late Paleozoic,in combination with morphological and anatomical characteristics and biogeographic distribution pattern. The results show that the records of Sphenopteris were widely distributed in the middle-low latitudes worldwide from the Pennsylvania to Cisuralian,indicating high variation and differentiation of Sphenopteris-type leaves. It is speculated that Sphenopteris species migrated from South China to North China during Late Devonian to Permian,and the diversity pattern was not only closely related to humid climate,but also coupled with the palaeogeographic characteristics of this genus.
晚古生代地层中发现的蕨类植物化石大多情况下是破碎和不完整的(Gastaldo, 1988; Taylor et al., 2009), 因此产生了许多分类位置不明的类群, 其中就包括多样性较为丰富的Sphenopteris(楔羊齿属)。该属通常被认为是由叶形态保存较好的畸羊齿类(mariopterids)、莲座蕨类(marattialeans)或者皱羊齿类(lyginopteridaleans)植物组成的分类位置尚不明确的庞杂类群(Taylor et al., 2009), 大多数为种子蕨类, 但也有少部分为真蕨类(古和植, 1974)①。楔羊齿类(sphenopterids)植物主要分布于晚古生代, 其中石炭纪、二叠纪期间分异度最高。该类植物的小羽片均属于Sphenopteris型, 呈羽状或者扇状。分布于东亚二叠纪的楔羊齿类一般具有小羽片质薄、叶脉纤细、羽轴着生角度较小的特征, 在中国主要分布在乌拉尔世地层中, 并且很少发现其表皮特征。目前, 关于Sphenopteris的研究主要集中在宏观形态学分析阶段, 且由于缺乏解剖学的综合分析, 对其系统分类位置归属的研究还较为初步, 有价值的系统发育信息亦较为匮乏。
文中报道了甘肃永昌乌拉尔世地层中发现的2种Sphenopteris。在此基础上, 结合其宏观形态、微细构造特征及晚古生代时期的全球化石记录, 综合分析了Sphenopteris属的系统分类属性及其在晚古生代的多样性演化和古地理分布特征。
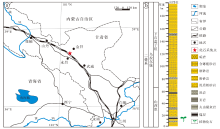
研究剖面位于甘肃省永昌县河西堡镇龙首山东段(GPS: 38° 28'33″N, 102° 101'32″E), 陆相二叠系发育完整, 产出植物化石(图1-a)。
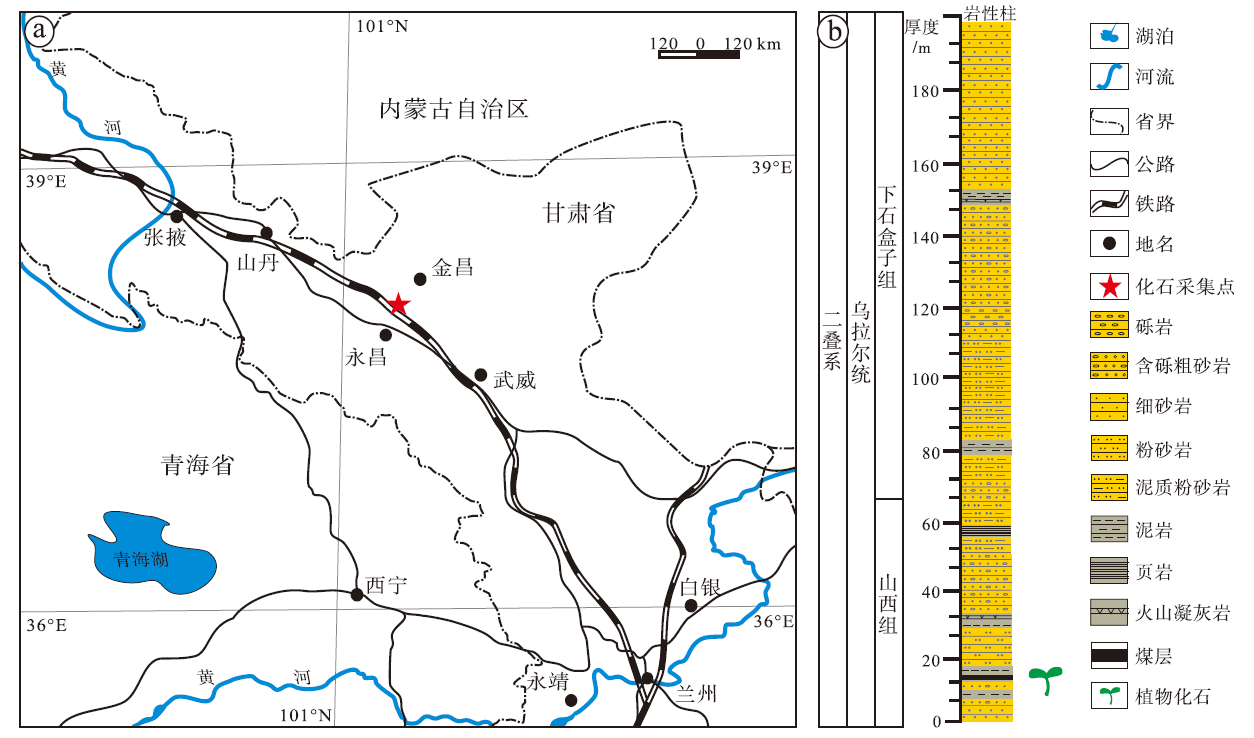 | 图1 甘肃永昌乌拉尔统地层柱状图及地理位置(据梁建德等, 1980) a— 化石采集点地理位置; b— 地层柱状图及化石产出层位Fig.1 Stratigraphic column of the Cisuralian and fossil sampling site in Yongchang, Gansu Province(after Liang et al., 1980) |
该剖面为一个向西抬起并封闭的向斜构造, 自上而下主要由石千峰组、上石盒子组、下石盒子组和山西组组成, 上覆地层是中三叠统的西大沟群, 下伏地层是乌拉尔统和宾夕法尼亚亚系的太原群(刘洪筹和沈光隆, 1978; 梁建德等, 1980)。其中乌拉尔统山西组厚约64.5 m, 分为7层(图1-b)(Wang et al., 2020, 2022; 王雪莲, 2020), 文中研究的植物化石材料均产自山西组第3层中。山西组自下而上岩性依次为浅黄褐色的薄层细砂岩, 灰白色、浅黄褐色含砾粗砂岩及石英砂砾岩, 夹灰黑色泥岩薄层; 第3层产大量的植物化石, 岩性为黑色泥岩; 第5至7层为灰黑色泥岩、粉砂岩, 局部夹砾岩, 最上部为黄绿色泥质粉砂岩(参见Wang et al., 2020)。
化石标本采用数码相机进行图像采集, 对标本的局部特征采用立体显微镜进行放大拍照。选择部分标本进行角质层微细构造分析, 分析过程中主要采用王水+次氯酸钠浸解法(王雪莲等, 2020)。对于一些碳膜保存较差但在体视镜下可观察到植物表面特征的化石, 采用扫基岩法获得化石表面的微细构造特征(徐小慧等, 2017), 而后在光学显微镜及扫描电镜下进行观察和拍照。所有的标本和薄片均保存于兰州大学地质科学与矿产资源学院古生物学与地层学研究所实验室。
楔羊齿类 Sphenopterids
楔羊齿属 Sphenopteris(Brongniart, 1822)Sternberg, 1825
永昌楔羊齿(新种)Sphenopteris yongchangensis Wang et Xiong sp. nov.
(图2-a; 图3; 图4)
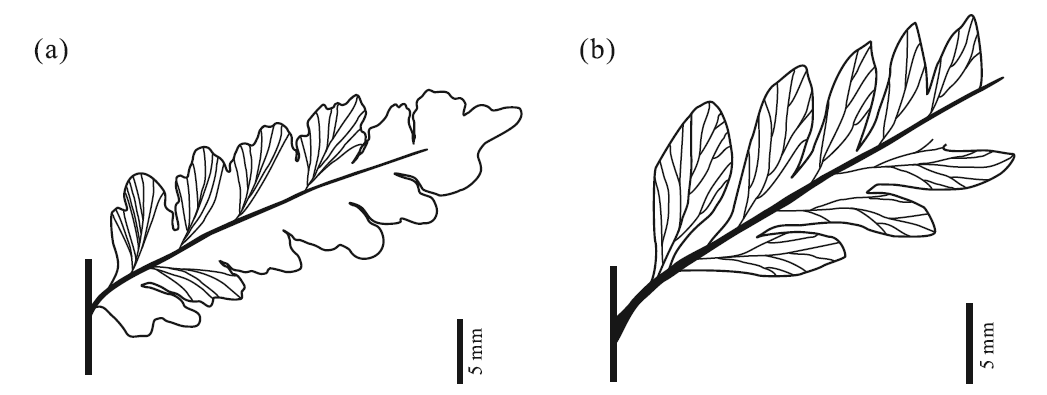 | 图2 甘肃永昌乌拉尔世2种Sphenopteris手标本素描 a— Sphenopteris yongchangensis sp. nov., 基于标本GSYC-LDGSW-2018-104(A, B)(图3-e); b— Sphenopteris nystroemii, 基于标本GSYC-LDGSW-2018-236(图5-g)Fig.2 Line drawing of two species of Sphenopteris from the Cisuralian of Yongchang, Gansu Province |
 | 图3 甘肃永昌乌拉尔世Sphenopteris yongchangensis sp. nov.形态特征 a, b, g— 末二次羽片的形态, 其中b为a中黄色方框的局部放大, 标本编号GSYC-LDGSW-2018-104(A, B); c, d— 标本编号GSYC-LDGSW-2018-164(A, B); e, f— 平行排列的末次羽片形态, 标本编号GSYC-LDGSW-2017-100(A, B); h-m— 小羽片形态, 标本编 号GSYC-LDGSW-2017-100(A, B)Fig.3 Morphological characteristics of Sphenopteris yongchangensis sp. nov. from the Cisuralian of Yongchang, Gansu Province |
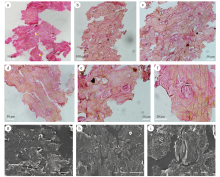
 | 图4 甘肃永昌乌拉尔世Sphenopteris yongchangensis sp. nov. 小羽片表皮微细构造特征 a— 上表皮在光学显微镜下的特征; b-e— 下表皮特征; g, h— 下表皮在扫描电镜下的特征; f, i— 气孔器形态。标本编号GSYC-LDGSW-2017-100(A, B)Fig.4 Pinnule cuticle micromorphology of Sphenopteris yongchangensis sp. nov. from the Cisuralian of Yongchang, Gansu Province |
正模标本: GSYC-LDGSW-2018-104(A, B)。
副模标本: GSYC-LDGSW-2018-164(A, B); GSYC-LDGSW-2017-100(A, B)。
词源: 种名 “ yongchangensis” 来源于当前化石的采集地点永昌县。
模式产地: 甘肃省永昌县河西堡镇。
地质时代和层位: 乌拉尔世/山西组。
鉴定特征: 末二级羽轴宽约5 mm, 末次羽片呈扇形, 互生于轴上。小羽片呈扇形, 边缘近乎全缘, 顶端较钝。小羽片大小约为12 mm× 7 mm, 基部彼此连合。中脉粗壮, 侧脉分叉1次。上表皮气孔作椭圆状, 其细胞呈矩形。下表皮气孔呈线形排列, 作椭圆状, 气孔器由2个半月形保卫细胞和6个副卫细胞组成, 下表皮细胞呈不规则的四边形。
标本描述: 当前标本保存了末二次羽轴和末次羽片的形态(图3-a至3-g)。末二级羽轴宽约5 mm, 轴表面特征不明(图3-g)。末次羽片呈扇形, 互生, 长约40 mm, 宽约15 mm, 末级羽轴宽约2 mm(图3-e至3-g, 3-l)。小羽片呈扇形, 边缘具齿, 基部与相邻小羽片完全相连, 顶端钝圆, 整个末级羽片上的小羽片基部几乎连合(图3-h至3-j)。小羽片中脉较粗, 自下而上逐渐变窄, 侧脉分叉1次, 小羽片长约12 mm, 最宽处约7 mm(图3-k, 3-l, 3-m)。
微细构造特征: 当前化石的气孔分布类型属于双生型。上表皮气孔分布较稀疏, 气孔器呈椭圆形, 直径约为10 μ m, 由2个呈半月形的保卫细胞组成, 表皮细胞呈规则的矩形(图4-a)。下表皮气孔分布较密集, 单列气孔呈线形排列, 列之间排列不规则(图4-b至4-d)。气孔器呈椭圆形, 直径约为15 μ m, 同样由2个呈半月形的保卫细胞组成, 由6个副卫细胞环绕而成(图4-e至4-i)。下表皮细胞形态不规则, 由长方形和不规则的四边形组成(图4-f)。
比较与讨论: 根据羽片特征将楔羊齿各个种进行详细对比(表1)。当前标本的小羽片形态与Sphenopteris neimongolensis、S. subsulcarta以及S. tingii较为相似: S. neimongolensis小羽片的侧脉分叉次数与羽轴夹角与当前化石一致, 但该种的小羽片叶缘全缘而区别于当前化石; S. subsulcarta的末二级羽轴较细而区别于当前化石; S. tingii的小羽片特征与当前化石非常相近, 但前者末次羽片与末二级羽轴夹角近似直角, 这一特征区别于当前化石的锐角。S. nystroemii末级羽轴与末二级枝夹角以及和小羽片的夹角都与当前化石较为接近, 但前者的小羽片基部与相邻小羽片分离且收缩、小羽片呈梭形而区别于当前化石。综上, 当前化石标本区别于楔羊齿属内的任何一个已知种, 因此将其定为一新种: Sphenopteris yongchangensis sp. nov.。
| 表1 Sphenopteris代表性种的特征对比(基于中国的化石记录) Table1 Morphological comparisons of representative Sphenopteris species(Based on fossil records from China) |
弱楔羊齿 Sphenopteris nystroemii Halle 1927
(图2-b; 图5)
 | 图5 甘肃永昌乌拉尔世Sphenopteris nystroemii形态特征 a, b— 末二次羽片的形态特征, 标本编号GSYC-LDGSW-065(A, B); c, d— 标本编号GSYC-LDGSW-2018-236; e— b中黄色方框局部放大的末次羽片的形态; f-i— 末次羽片和小羽片在立体显微镜下的特征, 标本编号GSYC-LDGSW-065(A, B); j, k— 小羽片在扫描电镜下 的表皮特征; l, m— j中黄色方框的局部放大, 可见气孔器特征(黄色箭头)Fig.5 Morphological characteristics of Sphenopteris nystroemii from the Cisuralian of Yongchang, Gansu Province |
1927 Sphenopteris nystroemii, Halle, 页57, 图版12, 图1-2
1974 Sphenopteris nystroemii, 古和植, 页77, 图版50, 图1-2
1987 Sphenopteris nystroemii, 赵修祜等, 页75, 图版10, 图1-1a
2010 Sphenopteris nystroemii, 孙克勤等, 页305, 图版178, 图2-3
化石标本: GSYC-LDGSW-065(A, B, C); GSYC-LDGSW-2018-236。
产地: 甘肃省永昌县河西堡镇。
地质时代和层位: 乌拉尔世/山西组。
标本描述: 标本保存了部分末二次羽片和完整的末次羽片(图5-a至5-d)。末二级羽轴宽约1.5 mm, 表面无纹饰(图5-f), 羽轴与末次羽片的夹角约为30° (5-e)。末次羽片互生于末二级羽轴的两侧, 呈披针形(图5-e)。末级羽轴宽约5 mm, 与小羽片的夹角约为45° (图5-g)。小羽片互生, 呈梭形, 整体纤细, 顶端钝尖, 长约3 mm, 最宽处约1 mm, 基部以窄的宽度着生(图5-h)。小羽片的中脉与侧脉等宽, 侧脉分叉1次, 且达缘(图5-i, 黄色箭头)。
微细构造特征描述: 当前标本角质层较薄, 因此采用扫基岩法获得表面部分的微细构造特征。仅观察到气孔分布在脉间区(图5-j, 5-k), 气孔几乎呈列分布(图5-l), 气孔器大小约为80 μ m× 110 μ m。
比较与讨论: 当前化石虽只保存了末二次羽片和末次羽片, 但化石整体纤细, 叶脉清晰, 化石的整体特征与产自山西太原下石盒子组中的S. nystroemii极为相似(表1)。赵修祜等(1987)和孙克勤(2010)对S. nystroemii进行了详细的描述, 当前化石材料均具备了与其一致的特征。因此将当前化石材料定为Sphenopteris nystroemii是合适的。
Sphenopteris代表了一个相对庞大且混乱的组合类群的叶片结构, 通常呈现为底部狭窄的羽状体。在晚古生代, 其形态上的主要特征为: 小羽片呈椭圆形轮廓, 几乎全缘, 侧脉分叉且延伸至边缘, 二级羽片呈明显的翅状外观(Taylor et al., 2009)。已有学者尝试对该属的分类位置进行划分, 依据的特征是整体或者茎杆部分的羽状结构, 但对于缺乏繁殖结构的情况还是较为困难的(van Amerom, 1975; Krings and Kerp, 2000)。目前研究认为, Sphenopteris可能归属于畸羊齿类(mariopterids)、莲座蕨类(marattialeans)或皱羊齿类(lyginopteridaleans)植物的叶结构部分(Gastaldo, 1988; Taylor et al., 2009)。由于无相关繁殖结构的对比材料, 因此笔者主要根据羽片的宏观和微观结构对其分类学属性进行分析讨论。
畸羊齿类最为代表的类群为Mariopteris和Karinopteris, 该类群地史时期的分布特征与Sphenopteris几乎相似, 均属于喜湿类植被(Boersma, 1972)。根据文中获得的Sphenopteris的微细结构特征, 畸羊齿类和Sphenopteris的气孔分布形式及形态大小均较为相似, 尤其是副卫细胞的排列结构, 但畸羊齿类羽状叶基部分裂呈耳状(Krings and Kerp, 2000), 与Sphenopteris小羽片结构形成强烈的对比。莲座蕨类属于真蕨植物的原始类群(刘璐等, 2020), 通常以生殖叶的形式保存, 且该类植物中、新生代占有绝对优势, 但目前化石记录显示莲座蕨目无论在个体数量还是属种数目上在晚古生代都不占优势(Lesnikowska and Galtier, 1992), 这与Sphenopteris丰富的化石记录似乎并不呼应。皱羊齿类是广泛分布于泥盆纪— 二叠纪的种子蕨植物, 是典型的攀爬类依生型类群(Taylor et al., 2009)。该类植物的茎分叉模式与目前发现的具分叉结构的Sphenopteris化石记录几乎一致(Meyer-Berthaud, 1990), 以及它们的小羽片形态和生长习性均具有相似性(Krings and Kerp, 2000), 因此将文中所描述的2种Sphenopteris暂时归为皱羊齿目(Lyginopteridales)更为合理, 后期还需要更多的化石证据进行补充和探讨。
3.2.1 Sphenopteris的物种丰富度变化规律
文中对Sphenopteris在全球晚古生代的地史分布做了详细的统计, 共计269个化石记录(见附加文件:晚古生代Sphenopteris分布统计表)并结合该属在全球范围内的分布对其多样性及古地理演化进行了详细的分析讨论。
全球范围内, Sphenopteris较为详实的记录最早出现于泥盆纪晚期(Sze, 1936; 赵修祜和吴秀元, 1986; 秦敏等, 2016), 石炭纪物种丰富度快速增加, 尤其在宾夕法尼亚亚纪达到了最高峰, 表明该类植物或具有楔羊齿型的化石叶片在石炭纪的高度分异(图6)。二叠纪中晚期, Sphenopteris物种丰富度明显降低, 至乐平世长兴期基本趋于灭绝。相比全球化石记录, 中国Sphenopteris自晚泥盆世始现后, 其物种丰富度在石炭纪有了一定的增长, 乌拉尔世达到了最高峰。
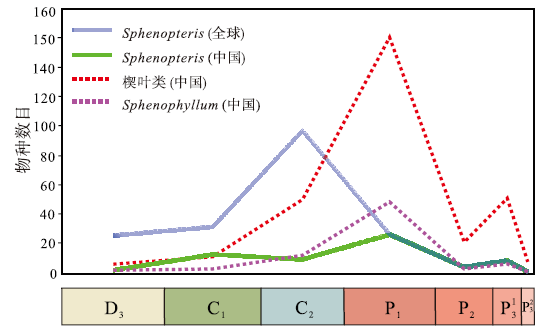 | 图6 晚古生代Sphenopteris的物种丰富度变化及与楔叶类植物的对比(其中楔叶类及楔叶属物种多样性数据引自邓珍珍等, 2021)Fig.6 Species richness of Sphenopteris during the Late Paleozoic and comparison with the diversity of sphenophytes(data of sphenophytes and Sphenophyllum based on Deng et al., 2021) |
楔叶类是晚古生代植物群的代表分子, 对它们在中国区域的多样性演化已有较为清楚的认识(邓珍珍等, 2021), 因此笔者将Sphenopteris和楔叶类的多样性演化模式进行对比。尽管两者的分类位置完全不同, 但仍可能会提供一些启示。该对比表明, 中国的Sphenopteris、Sphenophyllum与楔叶类整体的物种多样性变化趋势一致, 均于乌拉尔世最为繁盛。与晚古生代常以叶片形式保存的Sphenophyllum (楔叶属)对比发现, Sphenopteris和Sphenophyllum在中国均始现于晚泥盆世, 石炭纪二者的多样性增长幅度相似, 乌拉尔世多样性达到最高, 在乐平世晚期趋于灭绝(图6)。
Sphenopteris和楔叶类植物在晚石炭世至乌拉尔世的高度辐射, 代表了喜湿植物群对当时成煤期湿润环境的高度适应。这2类植物在晚古生代成煤期均为林下层植物, 其中楔叶类植物叶片在晚古生代显示了叶片面积逐渐增大及出现了叶镶嵌的特征(邓珍珍等, 2021), 而楔羊齿类植物同样显示了叶片形态的高度演化和分异。这可能指示当时这2大类植物的辐射演化受到了相似的环境驱动。
3.2.2 中国晚古生代Sphenopteris的地理分布和演化
结合文中永昌植物群中的新种, 目前中国所发现的Sphenopteris化石共有147个记录84个种(包含未定种和相似种), 在晚泥盆世至晚二叠世地层中均有报道, 其中泥盆纪和晚二叠世化石种数量较少, 石炭纪、早二叠世地层中最为常见, 尤其在石炭纪整体呈增长趋势, 乌拉尔世分异度最高且地理分布范围最广(图6; 图7)。Sphenopteris在中国晚古生代时期的分布特征基本与全球多样性演变趋于一致, 但欧美地区该属多样性在宾夕法尼亚亚纪达到最高。在晚泥盆世地层中, 共有3个种在华南地区被发现和报道(图7-a)(古和植, 1974; 秦敏等, 2016), 因而Sphenopteris在中国起源于晚泥盆世的华南地区。晚泥盆世华南广泛发育海陆过渡相地层(李星学等, 1995), 优越的气候条件有利于植物迁移和传播。
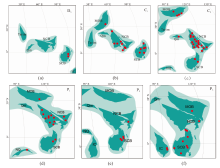
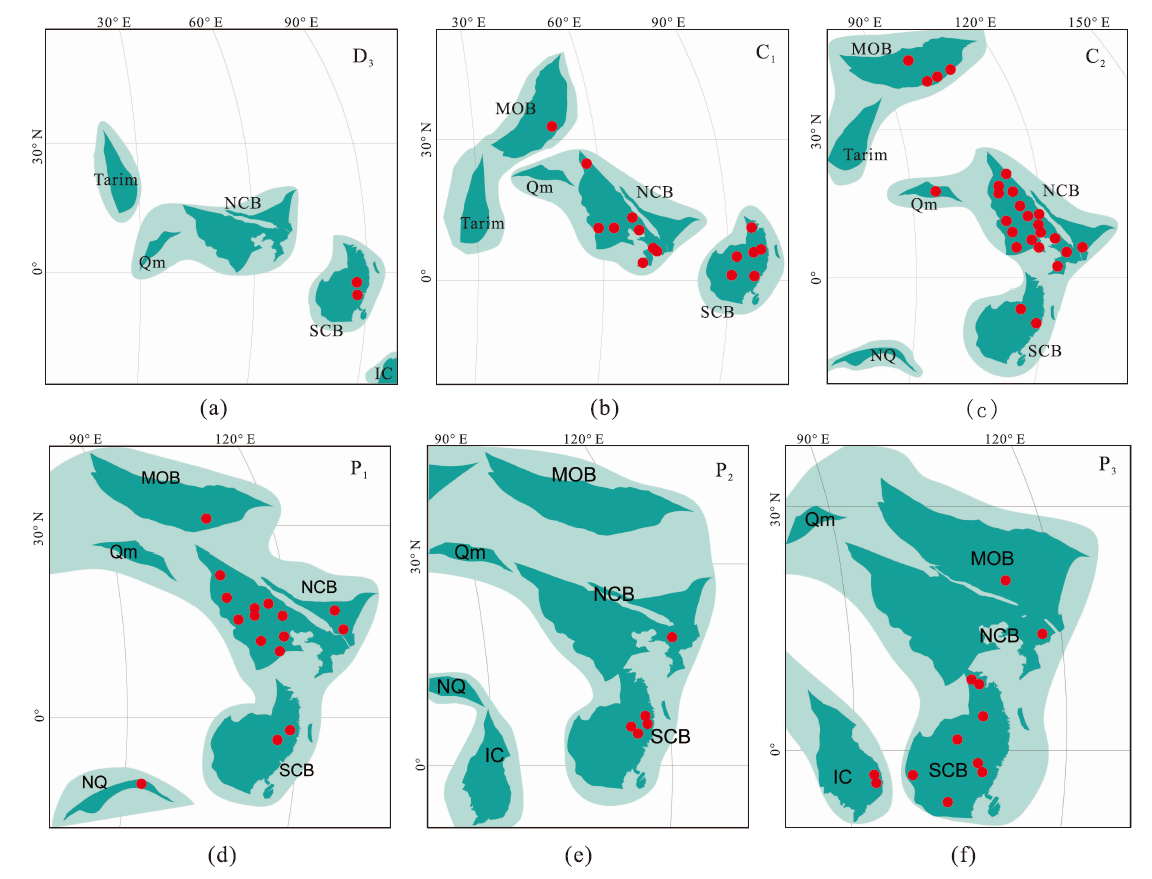 | 图7 晚古生代Sphenopteris在华南、华北及周缘块体的地理分布 a— 晚泥盆世(D3); b— 密西西比亚纪(C1); c— 宾夕法尼亚亚纪(C2); d— 乌拉尔世(P1); e— 瓜德鲁普世(P2); f— 乐平世(P3)。 古地理图依据Huang et al.(2018); NCB: 华北板块; SCB: 华南板块; Tarim: 塔里木板块; Qm: 柴达木地块; MOB: 蒙古板块; NQ: 北羌塘地 块; IC: 印支地块。 红点代表化石点, 数据来源详见附表1Fig.7 Geographic distribution of the Late Paleozoic Sphenopteris species across South China, North China, and other adjacent blocks |
在中国密西西比亚纪地层中, 约有16个Sphenopteris种被报道, 分别出现在江西、广西、宁夏和内蒙古等地(图7-b)(李汉民等, 1982; 赵修祜和吴秀元, 1982, 1986; 何锡麟等, 1990; 陈芬等, 1995)。密西西比亚纪植物群基本延续了晚泥盆世植物群的特征, 化石记录表明Sphenopteris在密西西比亚纪的华南地区有了进一步的演化; 宾夕法尼亚亚纪地层中, 共有25个种被报道, 分别在中国河北、内蒙古、辽宁、江西、甘肃和陕西等地(图7-c)(古和植, 1974; 冯少南等, 1977; 黄本宏, 1980; 赵修祜和吴秀元, 1986)。根据古地理资料, 整个宾夕法尼亚亚纪, 华南地区基本上被海水覆盖, 几乎无陆生植物的出现; 华北地区出现海陆交互相, 植物开始迅速繁衍, 为Sphenopteris在华北的分布提供了良好的条件(李星学等, 1995)。乌拉尔世地层中, 约有32个种被报道(图7-d), 瓜德鲁普世地层中约有6个种被报道(图7-e), 乐平世地层中约有12个种被报道(图7-f), 在二叠纪出现的Sphenopteris大多分布在华北地区, 化石产地分别位于山西、内蒙古、甘肃、河南等地, 只有少数几个种分布在华南和西南地区(冯少南等, 1977; 萧素珍和张恩鹏, 1985; 王德有, 1987; 赵修祜等, 1987; 何锡麟等, 1990; 孙克勤, 1992; 杨关秀, 2006)。华北二叠纪海陆交互地层最为发育, Sphenopteris的多样性在该时期最为丰富。三叠纪地层中, 共有5个种被报道, 分别在甘肃、辽宁和四川(郑少林和张武, 1981; 段淑英和陈晔, 1984; 蔡凯蒂, 1989), 之后出现的与之相似的极少数化石被认为是Sphenopteris, 文中不再纳入分析讨论。
综合以上的讨论分析, 关于Sphenopteris在中国的演化和迁徙路径可能为: 在晚泥盆世起源于华南, 至石炭纪向华北传播, 二叠纪该属在华北地区的多样性最为丰富。晚古生代, 华南、华北板块及其他块体处于古特提斯洋, 所处位置可能较为临近(图7), 为植物群的迁移、交流提供了可能。
角质层具有很高的保存潜力, 通常能保存叶片表皮的良好印迹, 为表皮微形态和解剖特征提供了良好的记录, 是分类学和生态学研究的重要资料(Taylor and Taylor, 1993; Psenicka et al., 2005; Barclay et al., 2007)。甘肃永昌乌拉尔世Sphenopteris的角质层均表现出叶近乎全缘、上表皮较下表皮厚且较为粗糙、下表皮薄、气孔器均无明显下陷的特征。通常全缘叶是现代热带雨林最典型的叶片类型, 且大型的叶片在成煤沼泽比在较干的雨林中常见, 且常常分布在林下层而非林冠层(Richards, 1952); 角质层上表皮较下表皮厚、气孔器无明显下陷均代表叶片长时期适应湿润环境(Tu et al. 1999, 2002; Kovar-Eder et al. 2001), 这也与石炭纪— 二叠纪时期大量喜湿的蕨类植物崛起(Wachtler and Perner, 2013)相吻合。
在古植物学的研究中, 分类位置不明的类群往往容易被忽视, 使其无法追根溯源, 但这些类群在古生代植被的研究中却屡见不鲜, 因此需要对其进行持续研究。中国西北地区乌拉尔世Sphenopteris的发现为进一步明确该属的系统分类位置提供了来自微观构造的证据。楔羊齿适应湿润环境的特征使得它们在石炭纪至乌拉尔世快速演化, 具有了极高的物种分异度和广泛的古地理分布特征。
1)记述了甘肃永昌乌拉尔统山西组中的2种楔羊齿: Sphenopteris yongchangensis sp. nov.和Sphenopteris nystroemii, 对其形态及微细构造特征进行了详细的系统描述; 结合其特征认为这2种楔羊齿归入皱羊齿目(Lyginopteridales)更为合理。
2)结合全球晚古生代楔羊齿的化石记录, 发现Sphenopteris的物种丰富度在石炭纪晚期最高。Sphenopteris在中国起源于晚泥盆世的华南地区, 而后向华北迁移演化; 此外, Sphenopteris化石角质层特征指示了该类植物具喜湿习性, 与晚古生代喜湿蕨类植物的兴衰以及它们的古地理分布特征相耦合。
(责任编辑 张西娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|