







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
生物岩的分类*
[吴亚生 ]
]
]
|
|

作者简介 吴亚生,男,1963年生,中国科学院地质与地球物理研究所副研究员,中国科学院大学岗位教授,主要从事碳酸盐岩地层古生物和生物岩研究。E-mail: wys@mail.igcas.ac.cn。
生物岩是新兴交叉学科生物岩石学的主要研究对象,是生物作用形成的岩石,主要包括生物礁岩、非造礁生物骨骼和泥晶组成的岩石、微生物骨骼组成的岩石、微生物作用形成的岩石。以往的生物礁和微生物岩分类系统只包括了一部分生物礁岩和微生物岩,尚有一部分宏体生物形成的岩石类型和微生物形成的岩石类型没有准确命名。为了促进生物岩的研究,作者提出一个包括生物礁岩、非造礁生物骨骼碳酸盐岩、微生物岩和其他微生物成因岩石的统一的生物岩分类系统,包括4个层级24种基本岩石类型。新增的基本岩石类型包括生物泥晶岩、微礁岩、微屑岩、包壳石、微骨架岩、微绑结岩、丝状岩、微泥晶岩等。作者还对部分岩石例如骨架岩等的定义进行了补充修正与完善,讨论了生物岩研究中存在的4个问题。
About the author WU Yasheng,born in 1963,is an associate professor at Institute of Geology and Geophysics,Chinese Academy of Sciences,and a professor at University of Chinese Academy of Sciences. He is mainly engaged in research of biopetrology and paleontology of carbonates. E-mail: wys@mail.igcas.ac.cn.
Bioliths are the main research object of the new interdisciplinary biopetrology,are rocks formed by biological processes,including reef rocks,rocks composed of non-reef-building organisms and micrite,and rocks formed by microbial processes. Previously,reef rocks and microbialites were studied separately and not included in the same discipline. The classical classification system of reef rocks and microbialites only includes a part of reef rocks and microbialites,and some biolith types are not recognized and included. The incomplete classification of bioliths limited the research of bioliths. In this paper,a unified classification system of bioliths including reef rocks,non-reef-building skeletal carbonates,microbialites and other microbiogenic rocks is proposed. The new basic bioliths include biomicrite,microlithite,microbialite,microallolite,micromicrite,framelite,crustolite,wrapolite,and filalite. This classification of bioliths into 24 basic rock types in 4 levels may facilitate the study of bioliths. The author also modified and improved the definition of some rocks such as framestone and discussed some problems in the study of biolith.
生物成因的碳酸盐岩以及生物成因的其他岩类, 称为生物岩(Wu, 2022a), 生物礁岩是典型的生物岩, 微生物岩也是典型的生物岩。除了生物礁岩之外, 非造礁生物也形成碳酸盐岩。除了微生物岩之外, 微生物还形成其他类型的岩石, 例如由经过风浪作用破坏的微生物骨骼组成的岩石。可见, 和生物相关的岩石是一个十分庞大的岩石群体, 近年来引起了地质学家的广泛关注。
前人对生物礁岩和微生物岩进行了分类和命名。在Folk(1959, 1962)的灰岩分类系统中, 灰岩被划分为异化颗粒灰岩、非异化颗粒灰岩和礁岩(biolithite)三大类; 并对前两者做了进一步划分, 但对礁岩未做进一步划分。在Dunham(1962)的碳酸盐岩分类系统中, 非礁相碳酸盐岩被划分为颗粒岩、泥粒岩、粒泥岩、灰泥岩(mudstone)、结晶碳酸盐岩, 生物礁岩被统称为“boundstone” 。Embry和Klovan(1971)首次对生物礁岩作了进一步分类, 划分为骨架岩、障积岩和绑结岩(bindstone); 此外, 在Dunham(1962)划分的非礁碳酸盐岩类型基础上, 新增加2种含砾(直径大于 2 mm)的岩石类型: 砾屑岩(rudstone)和漂砾屑岩(floatstone)。Wright(1992)增加了5种成岩成因的岩石: 胶结岩(cementstone)、亮晶岩(sparstone)、微亮晶岩(microsparstone)、致密颗粒岩(condensed grainstone: 很多颗粒之间是缝合线接触)、缝合颗粒岩(fitted grainstone: 颗粒之间是缝合线接触)。吴亚生(Wu, 1991, 1992)根据对中国南方广泛分布的中、上二叠统钙质海绵生物礁的研究, 定义了一种以具有稀疏的原地造礁型海绵动物骨骼为特征的生物礁岩—潜障积岩(prebafflestone)。金振奎和邵冠铭(2014)将生物礁岩划分为“礁石灰岩” 和“礁砾屑石灰岩” 。Lokier和Junaibi(2016)将生物礁岩划分为绑结岩(bindstone)、骨架岩和似绑结岩(boundstone: 指绑结方式无法识别的绑结岩)。
最初识别的微生物岩类型是叠层石(Kalkowsky, 1908), 而微生物岩的第2种类型——凝块石, 则是直到1967年才被识别(Aitken, 1967)。微生物岩这一术语, 直到1987年才被提出(Burne and Moore, 1987)。Burne和 Moore(1987)将微生物岩分为叠层石、凝块石、核形石、隐微生物岩(cryptic microbialites)、球形微生物岩(spherulitic microbialites)。Riding(2000, 2011)将微生物碳酸盐岩分为叠层石、凝块石、树形石、均一石。
虽然前人在生物礁岩和微生物岩分类方面取得了很多进展, 但是还存在一些问题: (1)生物礁岩被放在沉积岩中, 但是, 微生物岩尚无明确的归属; (2)有一些由生物形成的岩石, 包括宏体生物形成的和微生物形成的, 尚未被包括到现有的生物礁岩分类系统或微生物岩分类系统中; (3)缺乏一个包括所有生物成因岩石的分类系统。作者此文的目标是建立一个包括所有生物岩的分类系统, 并对前人分类系统中尚未包括的生物岩进行定义和命名, 抛砖引玉, 以期能够有助于生物岩石学的研究与发展。
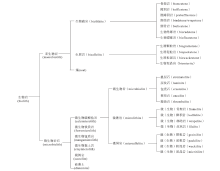
生物岩(biolith)指由生物作用形成的岩石。在前人的文献中, “biolith” 一词指由生物遗骸或生物成因的矿物组成的岩石, 吴亚生(Wu, 2022b)用它指所有由生物骨骼或生物作用形成的岩石。生物岩主要是碳酸盐岩。由于矿物成分和化学成分比较单一, 生物岩的分类主要根据结构、结构组成和形成机制。结构组成指岩石由哪些结构组分构成的、含量各是多少。生物岩常见的结构组分包括生物骨骼、微生物成因的泥晶纹层、微生物成因的微凝块、宏体生物或微生物成因的泥晶等。生物岩的结构, 指结构组分的排列方式。生物岩的形成机制包括生物控制矿化作用、微生物诱导结晶作用(以前称微生物诱导矿化作用)、障积作用、捕集作用(trapping)、风浪和流水的破坏作用等。作者首先根据形成生物岩的生物的大小, 把生物岩分为由宏体生物形成的宏生物岩(macrobiolith)和由微生物形成的微生物岩石(microbiolith; Wu, 2022b), 然后根据成分对宏生物岩和微生物岩石做进一步分类, 再根据结构组分、结构、形成机制进行第三级、第四级分类(图1)。
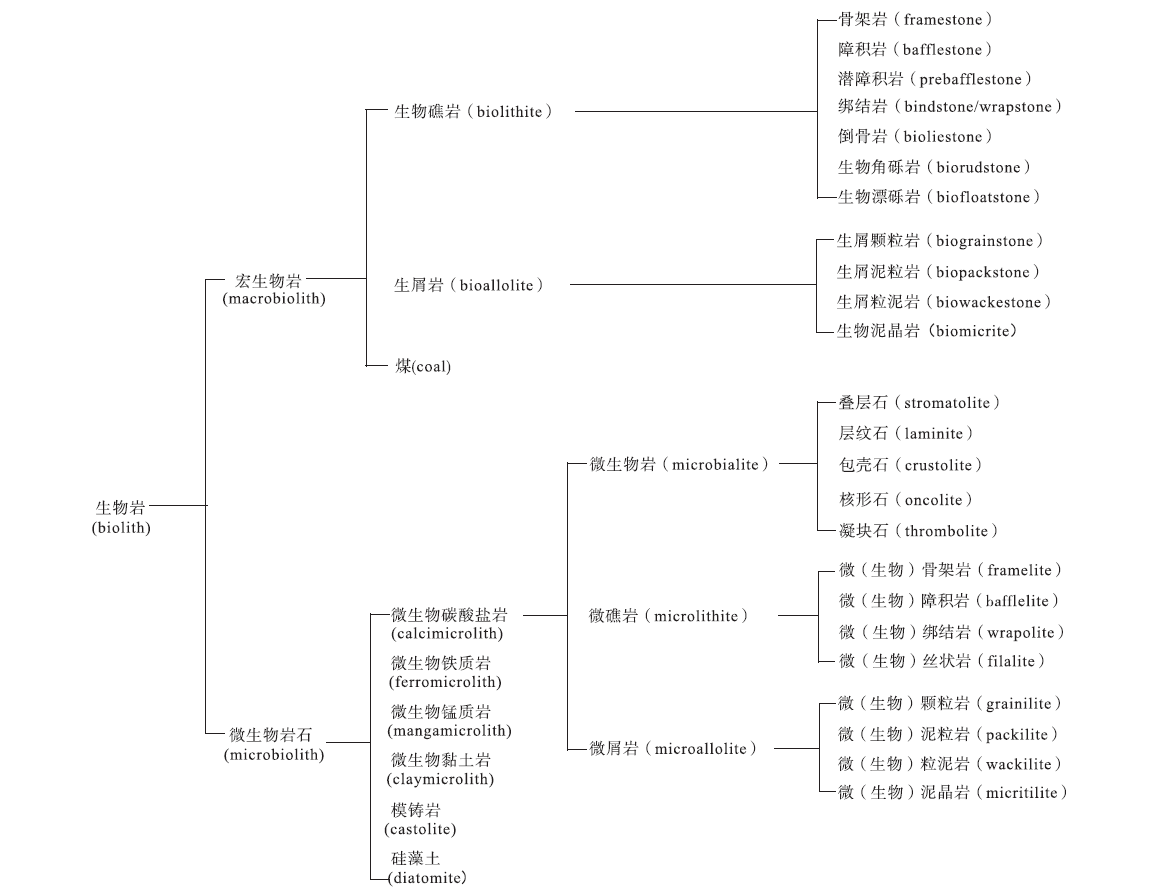 | 图1 生物岩的分类Fig.1 Classification of bioliths |
根据成分和结构可以把宏生物岩分为生物礁岩、生屑岩、煤。主要由宏体造礁生物的骨骼组成的岩石属于生物礁岩; 由宏体非造礁生物的骨骼组成的岩石属于生屑岩。由植物体堆积、并且经过降解、压实、热演化形成的碳质为主的岩石属于煤。作者主要聚焦于生物成因的碳酸盐岩, 所以下文重点介绍生物礁岩和生屑岩的分类。
2.1.1 生物礁岩(biolithite) 生物礁岩(biolithite)是Folk(1962)提出的一个术语, 指“未扰动的生物礁岩” , 由原地的宏体生物骨骼构成。生物礁岩的分类主要根据岩石的形成机制和结构。根据迄今的认识, 生物礁岩至少包括骨架岩、障积岩、潜障积岩、绑结岩、倒骨岩、生物角砾岩、生物漂砾岩这7种类型。
Embry和Klovan(1971)首次提出的生物礁岩分类包括3种岩石类型: 骨架岩、障积岩和绑结岩(“绑结岩” 在此前文献中多译作粘结岩)。
骨架岩的原始定义(Embry and Klovan, 1971)是具由原地块状造礁生物的骨骼构成的岩石骨架的岩石, 例如泥盆纪的块状层孔虫构成的骨架岩。这一定义的限定为块状造礁生物。但是, 地质时期的很多生物礁不一定由块状造礁生物的骨骼构成, 例如, 二叠纪生物礁主要由钙质海绵的骨骼构成, 而钙质海绵的骨骼一般是柱状的; 寒武纪的古杯动物礁主要由锥状的古杯动物骨骼构成; 奥陶纪的托盘海绵礁主要由锥状的托盘海绵骨骼构成。只要造礁生物的数量丰富, 以前的学者把这些柱状或锥状造礁生物骨骼构成的岩石也都归入骨架岩。这里涉及一个问题: 柱状和锥状的造礁生物是否能够形成岩石骨架, 什么情况下它们可以形成岩石的骨架?作者认为, 如果宏体造礁生物是柱状或锥状, 它们需要相互联结, 才能构成岩石的骨架; 联结的含义是观察到它们相互接触, 或者有皮壳状的钙质生物骨骼把它们缠绕起来。所以, 作者将骨架岩的定义修改为: 具有相互联结的造礁生物骨骼形成的骨架的岩石, 相互联结的形式包括骨骼附着在下面的骨骼之上生长或者皮壳状生物把柱状、锥状生物骨骼缠结到一起。骨架岩的关键鉴别特征, 不在于宏体造礁生物骨骼的数量, 而在于它们是否相互联结构成岩石的骨架。如果宏体造礁生物骨骼不相互联结的话, 即使数量丰富, 也不能归入骨架岩, 只能归入障积岩。
宏体造礁生物的骨骼一般都超过薄片的大小, 需要在露头或光面上观察岩石的结构、确定岩石的类型。需要在不同方向的露头上对造礁生物骨骼是否相互联结作出判断。新生代珊瑚礁系统中, 往往有块状珊瑚构成岩石骨架的典型骨架岩。在湖南慈利的二叠纪生物礁中, 有相互接触的丛状四射珊瑚骨骼构成的骨架岩(图2)。
 | 图2 湖南慈利二叠系丛状四射珊瑚骨架岩Fig.2 Photograph of an outcrop of coral framestone in the Permian reef in Cili, Hunan Province |
障积岩以具有能够起障积作用的造礁生物为特征。障积生物指在生物礁中起到阻挡水流、使其携带的沉积物沉积的生物(Embry and Klovan, 1971)。Embry和Klovan(1971)认为障积岩不常见, 障积作用是一种主观的解释; 他们归入障积岩的是一种具有分枝状珊瑚骨骼的岩石。Lokier和Junaibi(2016)主张废除障积岩这个术语。但是, 根据作者的研究, 障积岩在二叠纪钙质海绵生物礁中很常见, 所以此术语应予保留。造礁生物能否起障积作用应当取决于其密度: 越是密集, 越是能起障积作用。据此推理, 造礁生物稀少的情况下不能起障积作用。但是, 生物礁中造礁生物稀少的岩石也得有个名称, 因此, 吴亚生定义了“潜障积岩” 这一术语(Wu, 1991, 1992; 吴亚生, 1997; 李阳等, 2018)。究竟造礁生物的密度达到多少, 才会起障积作用?目前尚无实验数据, 作者根据经验给出1个大致的标准: 当岩石中造礁生物的间距小于其直径的3倍, 就可归入障积岩中。
绑结岩(bindstone)由板状生物骨骼绑结沉积物形成。绑结岩这一术语的认识过程比较复杂。在Dunham(1962)的碳酸盐岩分类体系中, 所有的生物礁岩被统称为boundstone, 所以, 这个英文术语应该翻译为“生物礁岩” ; 而在Embry 和 Klovan(1971)的分类系统中, bindstone被定义为与骨架岩和障积岩并列的一类, 指由生物骨骼的包裹作用和binding 作用形成的岩石。
Wright(1992)用boundstone代替Embry 和 Klovan(1971)的bindstone。Lokier和Junaibi(2016)把礁岩分为骨架岩、bindstone和boundstone, 用bindstone指生物binding沉积物形成的岩石, 用boundstone指binding作用难以识别的bindstone。作者认为: boundstone作为reef rocks的代名词不一定有存在的必要; Embry和Klovan(1971)的bindstone的图版是层孔虫礁岩; 层孔虫不存在所谓的“粘结作用” ——层孔虫原来属于腔肠动物, 现在属于多孔动物, 它们活着的时候不会“粘结” 沉积物、死了也不会“粘结” 沉积物, 当然, 如果是包在海底的骨骼碎块上生长, 的确有防止它们被风浪搅动的作用, 可以夸大一点儿说它们起了“绑结” 作用, 所以, 作者把这种岩石叫做绑结岩。以前的文献中多把bindstone翻译成粘结岩。这种翻译是根据英语词汇的意思, 而不是根据术语提出者实际要表达的意思。作者主张避免叫粘结岩, 因为叫粘结岩容易让人误解为生物用黏液粘结沉积物。到目前为止, 没有任何关于生物的黏液可以粘结沉积物形成沉积岩或生物岩的实例。绑结岩在中国存在的实例较多: 南海新生代生物礁(图3)和浙江的中二叠世生物礁都有钙质红藻绑结岩(图4), 新疆、陕西的晚奥陶世生物礁都有层孔虫绑结岩。
 | 图3 南海流花构造新生代生物礁钙质红藻绑结岩 皮壳状钙质红藻匍匐包裹沉积物; 南海东沙群岛, LH4-1-1井岩心光面照片Fig.3 Bindstone photograph of polished core sample from the Cenozoic reef sequence in Liuhua structure, South China Sea |
 | 图4 浙江桐庐冷坞剖面中二叠统钙质红藻绑结岩 皮壳状钙质红藻石叶藻属(re)包裹在钙质海绵(sp)和苔藓虫 (br)外面。薄片透射光显微照片Fig.4 Photomicrograph of red algal bindstone from the Middle Permian reef at Lengwu outcrop, Tonglu, Zhejiang Province |
在二叠纪钙质海绵生物礁中, 常见皮壳状生物骨骼层层包裹在柱状钙质海绵外面的情况。这是绑结岩与骨架岩的过渡类型。当绑结生物骨骼的体积大于柱状生物骨骼时, 叫骨架—绑结岩(图5); 反之, 叫绑结—骨架岩。
 | 图5 湖南慈利高峰剖面上二叠统长兴组骨架—绑结岩 A—纹层状的古石孔藻(bin)包裹在钙质海绵骨骼(ske)外, 剩余孔隙被方解石胶结物(cem)充填; B—是A的中部的局部放大Fig.5 Photograph of frame-bindstone in the Upper Permian Changxing Formation reef in Cili Gaofeng outcrop, Hunan Province |
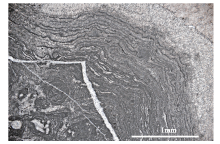
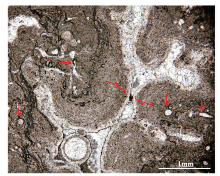
吴亚生等(Wu, 1991, 1992; 吴亚生, 1998; 李阳等, 2018)还定义了一种主要由倒伏的造礁型海绵动物的骨骼组成的岩石, 称为倒骨岩(bioliestone)。倒骨岩的主要特征是大多数造礁生物骨骼已被波浪作用打倒。在既有倒伏造礁生物骨骼又有原地造礁生物骨骼的情况下, 将倒伏造礁生物骨骼含量大于原地造礁生物骨骼含量的岩石叫倒骨岩。湖北利川柏杨见天坝和湖南慈利高峰乡上二叠统钙质海绵礁剖面都有钙质海绵倒骨岩(图6)。此外, 湖南慈利高峰乡上二叠统有丛状四射珊瑚形成的骨架岩和倒骨岩(图7)。
 | 图6 湖北利川柏杨镇见天坝村剖面上二叠统生物礁中的钙质海绵倒骨岩(据Wu, 2022b) A—礁体上部的一处露头照片, 红色箭头: 与层理平行的裂缝; 白色箭头: 岩层顶面朝向照片的右上角; 黄色箭头: 与层理平行的4个柱状钙质海绵骨骼(红色箭头); B—来自礁体底部的一薄片(JTBN-8-3A)的显微照片, 红色箭头: 示顶底构造, 注意与层理平行的白色胶结物和暗色泥晶之间的界面; a、b、d: 柱状纤维海绵动物骨骼; c、e: 柱状串管海绵动物骨骼; a、e朝向我们; b、d与照片平行; c倒转Fig.6 Bioliestones in the Upper Permian reef at Baiyangzhen Jiantianba outcrop, Lichuan, Hubei Province(after Wu, 2022b) |
 | 图7 湖南慈利高峰剖面上二叠统丛状四射珊瑚倒骨岩 倒伏和翻转的四射珊瑚(rugose coral)体积大于岩石体积的50%Fig.7 Photograph of bioliestone in the Upper Permian reef in Gaofeng outcrop of Cili, Hunan Province |
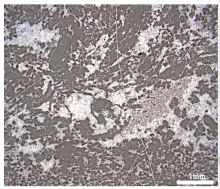
Embry和Klovan(1971)定义了砾屑岩(rudstone)和漂砾屑岩(floatstone)2种非礁岩石, 分别指砾级颗粒相互接触和不相互接触的岩石。风化成因的角砾组成的岩石属于普通沉积岩, 不属于本文讨论的范畴。碳酸盐岩分类一般限于盆内沉积、无远距离搬运形成的颗粒和泥晶, 颗粒包括内碎屑、鲕粒、球粒、生屑等。除了生物骨骼有较大者外, 其余种类的颗粒大小一般都小于0.5 cm。相反, 造礁生物骨骼一般大于1 cm。砾级的生物骨骼需要区分非礁相的与礁相的。因为, rudstone和floatstone被定义为非礁相的岩石类型, 所以, 需要专门为生物礁相中主要由非原地的砾级造礁生物骨骼组成的岩石定义名称。吴亚生(1997)提出将主要由造礁生物骨骼的角砾组成的岩石称为生物角砾岩(biorudstone; 砾相互接触)或生物漂砾岩(biofloatstone; 砾不相互接触)。生物角砾岩和生物漂砾岩都可以根据生物骨骼的类型和大小进行进一步描述, 例如, 珊瑚砾岩、层孔虫漂砾岩, 珊瑚巨砾岩、珊瑚中砾岩、珊瑚细砾岩。中国南海的一些现代珊瑚礁的礁脊相由5~50 cm的珊瑚砾块组成(王国忠, 2001), 它们在未来被埋藏和胶结后, 就是典型的生物角砾岩。目前尚未在二叠纪钙质海绵生物礁中见到生物角砾岩, 但是, 常见到含有折断的海绵骨骼的钙质海绵漂砾岩(图8)。
 | 图8 湖北利川柏杨镇见天坝村剖面上二叠统生物礁的钙质海绵漂砾岩 见到2个折断的、平行于层面的柱状钙质海绵骨骼(SP); 红色箭头垂直于层面, 指向上方Fig.8 Photograph of floatstone in the Upper Permian reef at Baiyangzhen Jiantianba outcrop, Lichuan, Hunan Province |
生物角砾岩与倒骨岩的区别是, 前者由相互接触的大型块状造礁生物骨骼的碎块组成, 而后者由相互接触的、倒伏的、比较完整的柱状造礁生物骨骼组成。倒骨岩与生物角砾岩的差别是不同时代、不同环境下不同造礁生物、遭受不同强度的风浪作用造成的。
礁岩的类型主要取决于造礁生物的类型和古水深这2个因素。在浪基面以下的环境中, 如果条件适宜, 就会有珊瑚、钙质海绵等造礁生物生长; 初期, 造礁生物稀疏, 如果因为环境原因, 造礁生物死亡, 形成的岩石叫潜障积岩。在造礁生物稀疏生长的情况下, 如果海水变浅, 风浪将沉积物搅动, 造礁生物被埋葬、死亡, 形成的岩石叫倒骨漂砾岩。如果造礁生物密集生长, 被风浪搅动、埋葬, 或者由于风浪不断搅动, 使之倒伏和滚动、无法继续生长而死亡, 形成的岩石叫倒骨岩; 如果因为风浪作用, 密集生长的造礁生物被打碎, 形成的岩石叫角砾岩。根据对大量二叠纪钙质海绵礁的观察, 海绵倒骨漂砾岩、海绵倒骨岩都比较常见。六射珊瑚礁可以生长到海平面附近, 遭受强烈风浪作用, 在礁脊形成大型角砾的堆积, 叫珊瑚角砾岩; 大型的珊瑚角砾也可以滚落到礁前的沉积物中, 形成珊瑚漂砾岩。在现代珊瑚礁的礁脊和礁后潟湖相中, 珊瑚角砾是主要沉积物; 它们被埋藏、成岩之后, 就是生物角砾岩。
2.1.2 生屑岩(bioallolite) 生屑岩(bioallolite)指主要由非造礁宏体生物的骨骼或骨骼碎屑和/或宏体藻类形成的泥晶组成的非礁相碳酸盐岩。已经广泛使用的颗粒岩、泥粒岩、粒泥岩和泥晶灰岩(又叫灰泥岩)等术语是根据组构来定义的, 而不是根据它们是否由生物成因的骨骼和泥晶组成(Dunham, 1962)。区分生物成因与非生物成因的碳酸盐岩组分是科学研究进步的必然, 是生物岩石学的基本思想之一。因此, Wu(2022b)定义了生物颗粒岩、生物泥粒岩、生物粒泥岩和生物泥晶岩(biomicrite: Wu, 2022b)4个术语, 来指宏体生物的骨骼、宏体生物形成的泥晶为主要成分的岩石。生物颗粒岩指非造礁生物的骨骼或其碎屑相互接触、无泥晶基质的岩石; 生物泥粒岩指非造礁生物骨骼或其碎屑相互接触、具有泥晶基质的岩石; 生物粒泥岩指非造礁生物的骨骼或其碎屑分散在泥晶中、且其含量大于10%的岩石; 生物泥晶岩指生物泥晶含量大于90%的岩石。生物泥晶主要来自不完全钙化的宏体钙化绿藻死亡后的分解。可以根据骨骼的生物类别和大小, 对生物颗粒岩、生物泥粒岩、生物粒泥岩作进一步描述, 例如腕足类砾屑颗粒岩、三叶虫砾屑颗粒岩。
主要由生物颗粒岩、生物泥粒岩组成的丘状岩体称为生物滩; 主要由生物粒泥岩、生物泥晶岩组成的丘状岩体称为生物泥晶丘或生物灰泥丘。如果岩石中泥晶的来源不确定, 则不能归入生物岩中。
碳酸盐泥晶包括4种成因: 不完全钙化绿藻死亡分解形成的、微生物成因的、无机成因的、大碳酸盐颗粒遭受生物破坏或机械磨蚀形成的。今后的研究应尽可能区分这4种不同成因的泥晶。
微生物岩石(microbiolith)指由微生物作用形成的岩石(Wu, 2022b)。由于中文微生物岩一词已经被(microbialite)占用, 包括microbialite的microbiolith只好用微生物岩石一词做名称。根据矿物组成, 可把微生物岩石分为微生物碳酸盐岩、微生物铁质岩、微生物锰质岩和微生物黏土岩; 它们分别由微生物形成的碳酸盐矿物、铁矿物、锰矿物和黏土矿物组成。根据其成因和组构, 可以把微生物碳酸盐岩分为微生物岩、微生物礁岩和微生物屑岩。
2.2.1 微生物岩(microbialite) 根据形态和内部结构, 将微生物岩分为叠层石(Kalkowsky, 1908)、层纹石(Aitken, 1967; Monty, 1976; Li et al., 2021)、包壳石(crustolite; Wu, 2022b)、核形石、凝块石(Aitken, 1967)。
叠层石是最早识别的微生物岩石类型, 也是最容易识别的微生物岩类型, 以具有柱状、丘状、分枝状的宏观外形和微生物席成因的纹层状内部显微结构为特征。根据成因, 可以把叠层石分为2类: 诱导钙化型和捕集型。前者由微生物席的诱导钙化形成的暗色泥晶纹层构成岩石骨架, 其间可以为沉积物、胶结物或孔隙。后者由非钙化微生物席与沉积物的交替形成, 其中的微生物席一般在死亡后降解掉, 留下层状的孔洞。这种叠层石的形成需要微生物席和沉积物早期固结2种作用: 没有微生物席就不能形成特有的纹层结构; 没有早期固结, 纹层结构就无法保存下来。根据研究, 与捕集型叠层石相比, 诱导钙化型叠层石数量更多、更重要。
层纹石(laminite)指微生物席形成的层状碳酸盐岩(Aitken, 1967)。层纹石与叠层石的区别仅仅在于前者的宏观形态呈层状, 后者呈柱状、丘状、分枝状等形状。层纹石(laminite)这一术语在国外文献有较多使用, 国内学者对这一术语不够了解, 往往把层纹石作为一种层状的叠层石。根据研究, 层纹石的数量远比叠层石多(Li et al., 2021; Wu et al., 2021)。
包壳石(crustolite)指主要由包裹在其他生物骨骼、或碎屑、或任何物体上的微生物膜形成的泥晶纹层组成的岩石(Wu, 2022b), 包壳状纹层的体积需要大于被包裹的物质。贵州紫云松山谈陆寨、湖北利川柏杨镇见天坝村上二叠统长兴组钙质海绵生物礁中常见由微生物成因的暗色泥晶碳酸盐纹层包裹在钙质海绵骨骼或生物骨骼碎屑上形成的包壳石(图9)。陕西耀县桃曲坡、岐县烂泥沟中奥陶统常见主要由包裹在单体珊瑚骨骼上的微生物膜形成的泥晶碳酸盐纹层组成的包壳石。青海花石沟新生代生物礁中有微生物膜形成的泥晶纹层包裹在细小凝块石外面形成的包壳石(图10)。
 | 图9 贵州紫云松山谈陆寨上二叠统叠层石—包壳石 钙质海绵生物礁中的包壳石, 主要由微生物席形成的暗色泥晶纹层组成Fig.9 Photomicrograph of thin section of crustolite in the Upper Permian reef at Tanluzai, Songshan of Ziyun, Guizhou Province |
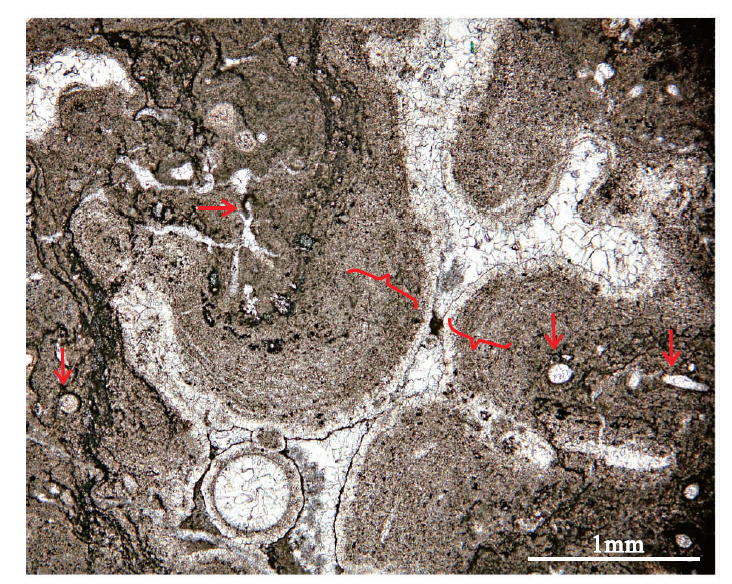 | 图10 青海花石沟新生代泥晶灰质包壳石 微生物成因的泥晶纹层(红色大括号)包裹在分枝丝状蓝细菌模孔(红色箭头)的壳之外(杭州地质研究院朱超提供薄片)Fig.10 Photomicrograph of crustolite in the Cenozoic reef at Huashigou, Qinghai Province |
核形石(Oncolite)是一种具有微观同心层状内部结构的球形结构(Pia, 1927; Peryt, 1981; Burne and Moore, 1987; Han et al., 2015; 吴亚生等, 2018), 由微生物形成的泥晶纹层层层包裹在某种核心(如岩石碎屑、半固结沉积物的碎屑、生物骨骼的碎屑)上形成。
凝块石被定义为具有凝块结构的微生物岩(Aitken, 1967; Riding, 2011)。但什么是凝块结构, Aitken(1967)没有给出明确的定义, 这导致其他学者不易把握。很多学者用宏观或中观的“凝块结构” 来识别凝块石, 例如Riding(2011)。但这种宏观或中观的“凝块结构” 不一定是微生物作用形成的; 水流冲洗作用、溶蚀作用、不均匀重结晶作用都可以造成类似凝块结构的假凝块结构。凝块石的识别必须根据真正的凝块结构。直接由微生物作用形成的凝块结构才是真正的凝块结构。如果凝块石具有几毫米大小的溶洞, 会形成漂亮的假凝块结构。新疆下寒武统肖尔布拉克组有漂亮的假凝块结构。生物控制钙化作用形成的具有特定形状的生物骨骼或微生物骨骼, 不叫凝块。因此, 一部分学者把具有Epiphyton、Renalcis构成的骨架的岩石叫做凝块石是不妥的。
微生物诱导矿化指微生物生理活动导致纳米到微米级碳酸盐矿物在其周围结晶。凝块的特征包括: 由泥晶组成、厚度和形状不规则、围绕模孔分布、在薄片中呈暗色。真正的凝块是微米级的; 厘米和亚厘米级的“凝块” 其实是由凝块和其间的微亮晶组成的岩石。Wu(2022b)将凝块石定义为具有微生物作用形成的深色显微泥晶凝块组成的骨架的岩石。根据形状, 可以把显微凝块结构划分为斑点状、蠕虫状、网状等类型(Li et al., 2021)。陕西耀县桃曲坡水库附近中奥陶世凝块石具有网状显微凝块结构(图11)。
 | 图11 陕西耀县桃曲坡中奥陶世网状凝块石 薄片(照片号0C9A7220), 偏光显微镜透射光照片; 暗色的是泥晶凝块, 孔隙被胶结物(白色)和沉积物充填Fig.11 Photomicrograph of reticular thrombolite in the Middle Ordovician at Taoqupo, Yaoxian, Shaanxi Province |
尽管leiolite(“均一石” )被定义为内部结构均一的微生物岩(Braga et al., 1995; Riding, 2011), 但因为缺乏识别的标志和实例, 建议废除它。一些中国学者报道了一些均一石, 但是, 其照片特征不符合均一石的定义。例如, 肖恩照等(2020)报道的河北涞源祁家峪剖面寒武系长山组的“均一石” 具有相互接触的原地附枝藻骨骼, 应属于微骨架岩。
2.2.2 微(生物)礁岩(microlithite) 微生物礁岩以具有丰富的原地微生物骨骼, 且其中的微生物骨骼具有规则的形状、可以进行生物学或古生物学的分类鉴定为特征(Wu, 2022b)。微生物骨骼即钙化微生物的钙化部分, 是具有特定形状和大小的矿物结构, 一般与微生物本身的形状和大小相似。就蓝细菌而言, 奥陶纪的钙化蓝细菌, 至少有20个属(Liu et al., 2016), 未钙化的蓝细菌数量未知。
微生物矿化分为微生物控制矿化和微生物诱导矿化; 微生物控制矿化指在微生物胶鞘内或细胞内进行矿物结晶、从而形成特定形状和结构的骨骼的作用。微生物诱导矿化指形成厚度不均匀、表面不平坦的矿物壳包在微生物体外面的矿物结晶作用(吴亚生等, 2021)。微生物形成骨骼和矿物壳都是普遍现象。主要由原地微生物骨骼组成的岩石叫微生物礁岩; 主要由原地微生物矿物壳组成的岩石叫微生物岩。除了矿物壳组成的微生物岩, 还有微生物席捕集作用形成的叠层石。无论是微生物礁岩, 还是微生物岩, 只要形成丘状的结构体, 就可以叫微生物礁。
微生物礁岩的进一步分类主要根据微生物骨骼的数量和形态。目前, 至少包括4种类型: 微(生物)骨架岩、微(生物)障积岩、微(生物)绑结岩(wrapolite)、微(生物)丝状岩。微骨架岩(framelite)指具有由底栖微生物的原地骨骼构成的骨架的岩石(Wu, 2022b), 其典型代表是Epiphyton微骨架岩、Renalcis微骨架岩。Epiphyton、Renalcis是古生代碳酸盐岩地层中常见的钙化微生物化石。新疆塔中上奥陶统礁滩相常见蓝细菌骨骼组成的微骨架岩(Liu et al., 2017: Fig.12-A; Fig.13-A, 13-B, 13-C, 13-G, 13-L)。
 | 图12 新疆阿克苏西南苏盖特布拉克下寒武统玉尔吐斯组中的微丝状岩(据Wu, 2022b) Girvanella微丝状岩的显微照片, 岩石骨架由原地的丝状微生物骨骼(可能是Girvanella)构成Fig.12 Girvanella filalite in the Cambrian Yuertusi Formation at Sugaitblake, southwest of Aksu, Xinjiang(after Wu, 2022b) |
 | 图13 层纹石—颗粒岩(A)和微生物骨屑颗粒岩(B)的显微照片(据Wu, 2022b) A—新疆阿克苏西南苏盖特布拉克村附近埃迪卡拉系奇格布拉克组的层纹石—层纹石屑颗粒岩, 由微生物席成因的暗色泥晶薄纹层(黑色箭头)和以层纹石屑为主的浅色厚纹层(紫色箭头)的交互组成; 部分层纹石屑内部由于发生重结晶而呈白色; B—新疆阿克苏西南苏盖特布拉克村附近寒武系第三阶云质肾形藻屑颗粒岩, 已经重结晶成粉晶白云岩, 由肾形藻屑(红色箭头)组成Fig.13 Laminite-grainilite(a)and light-colored thick laminae of laminitic debris, and skeletal grainilite(b)(after Wu, 2022b) |
在此前的文献中, 微生物骨架岩一般被归在微生物岩中。但根据定义, 微生物岩是微生物诱导矿化作用(microbially-induced biomineralization)和捕集作用(trapping)形成的, 而微生物礁岩由微生物的控制矿化作用(microbially-controlled biomineralization)形成。此前学者定义的枝状岩(dendrolite; Riding, 1988, 1991), 以在中观尺度上(厘米级)具有树枝状外观、由Epiphyton组成为特征。根据作者的分类, 应将它归入微生物骨架岩。
山东济南寒武系第三阶张夏组有Epiphyton构成骨架的岩石。Riding(2000: 图13)将其归入枝状岩(dendrolite), 但Lee等(2014: 图9)将其归入骨架岩(framestone), 而Yan等(2017)将其归入凝块石。由于这种岩石不是宏体生物形成的, 不应该归入骨架岩; 又因其不符合凝块石的定义, 不应该将它归入凝块石, 作者认为而应当归入微生物骨架岩。
微(生物)障积岩(bafflelite)的特点是在泥晶为主的基质中有丰富的(体积含量大于15%, 个体的间距小于个体直径的5倍)底栖微生物的原地骨骼(Wu, 2022b)。
微(生物)绑结岩(wrapolite)指由皮壳状钙化微生物骨骼层层包裹在物体表面形成的岩石。例如, 新疆上奥陶统良里塔格组礁滩相地层常见到皮壳状蓝细菌骨骼Wetheredella和 Rothpletzella的包壳组成的岩石(Liu et al., 2017: Fig.11-G, 11-H, 11-I)。
微(生物)丝状岩(filalite)指具有由丝状微生物的骨骼构成骨架的岩石(Wu, 2022b)。丝状岩在新疆上奥陶统和下寒武统玉尔吐斯组(图12)有发现。新疆塔中上奥陶统礁滩相常见蓝细菌骨骼组成的骨架岩(Liu et al., 2017: Fig.12-G, 12-J, 12-K)。
2.2.3 微(生物)屑岩(microallolite) 微(生物)屑岩(microallolite)指主要由微生物岩的碎屑、非原位微生物骨骼、微生物骨骼的碎屑、微生物形成的泥晶组成的岩石(Wu, 2022b)。
仅由微生物成因的颗粒(包括微生物岩的碎屑、非原位微生物骨骼、微生物骨骼的碎屑)和胶结物组成的岩石, 叫微生物颗粒岩。根据颗粒的成分, 分为微生物岩屑颗粒岩、微生物骨屑颗粒岩(图13-B)。根据颗粒成分, 可以将微生物岩屑颗粒岩分为叠层石屑颗粒岩、层纹石屑颗粒岩、凝块石屑颗粒岩等。微生物成因的颗粒相互接触、填隙物为微生物泥晶的岩石叫微生物泥粒岩。微生物颗粒分散存在于微生物泥晶中, 且微生物颗粒含量大于5%的岩石, 叫微生物粒泥岩。微生物泥晶含量超过95%的岩石叫微生物泥晶岩。若为微生物颗粒岩与微生物岩或微生物礁岩过渡的情况, 其命名根据主要成分而定(图13-A)。
微生物铁质岩、微生物锰质岩和微生物黏土岩可参照微生物碳酸盐岩的分类方法进行分类。
一般来说, 生物岩最初是由方解石或/和文石矿物组成的。 然而, 许多古老的生物岩已变成白云岩。 那么, 这些岩石怎么命名呢? 这里建议, 根据白云石的含量, 在图1中的岩石名称前面添加“云质” 、 “灰云质” 、 “云灰质” 这样的修饰词, 分别指白云石含量>95%、 >50%、 <50%的生物岩。 还可以在“云质” 、 “灰云质” 、 “云灰质” 的前面加上表示晶体大小的修饰语, 如“粉晶云质” 、 “细晶云质” 。 粉晶云质肾形藻屑颗粒岩指由于成岩作用, 95%以上的矿物已经转变为粉晶白云石的肾形藻屑颗粒岩。
杂微生物岩石指由硅酸盐岩岩屑、碳酸盐胶结物、鲕粒、微生物成因纹层、骨骼颗粒等2种或2种以上成分组成的岩石。Riding(1988)、 Riding和Virgone(2020)等对杂微生物岩石的分类和命名进行了深入的分析。当遇到杂微生物岩石时, 他们的分类方案是有用的。
生物岩的识别需要野外露头和室内薄片的双重研究为基础。有些珊瑚的直径可达1 m以上, 其形成的生物礁岩的结构必须在露头上确定。大多数微生物岩石必须根据薄片上的微观结构来确定, 尽管前人是根据中观结构(厘米尺度的)来定义和识别微生物岩的(Riding, 2011)。
生物岩由生物作用形成。虽然很多生物岩的形成过程中有沉积作用的影响, 但是, 生物岩的形成不需要沉积作用介入; 从本质上讲, 沉积作用对生物岩的形成主要起破坏作用, 而不是建设作用。生物礁岩主要由原地生长的造礁生物骨骼组成。在没有风浪的情况下, 造礁生物可以原位完好保存。但是, 有风浪作用的话, 一些甚至全部的造礁生物可能会被打倒, 从而形成倒骨岩或生物角砾岩。微生物岩石中人们最熟悉的是叠层石。一部分叠层石的成因被认为与微生物对沉积物的“捕集作用” (trapping)有关, 但这种类型只占叠层石的一小部分; 大多数叠层石是微生物席诱导钙化作用形成的, 而且叠层石在微生物岩石中只占一小部分。根据对新疆下寒武统微生物岩石的研究, 数量最多的微生物岩石类型是凝块石(Li et al., 2021)。而凝块石的成因主要是微生物诱导的矿物结晶作用。所以, 微生物岩石的形成主要靠微生物的作用。正因为如此, 对叠层石的定义强调它们以具有微生物席矿化形成的泥晶微纹层组成的骨架为特征; 对凝块石的定义强调它们要有微生物诱导结晶作用形成的暗色泥晶微凝块组成的岩石骨架。对生屑岩而言, 非造礁生物骨骼也是生物控制矿化作用形成的。非造礁型生物骨骼是生屑岩的物质基础; 没有沉积作用, 它们一样能形成岩石。所以, 可以简单地说: “绝大多数生物岩是生物长出来的” ; 生物岩的形成机制与沉积岩的形成机制有根本的不同。
生物岩与沉积岩的形成机制不同, 导致研究方法的不同。所以, 作者认为需要将生物岩从沉积岩中分离出来, 用生物岩石学的方法去研究可能更有利于该类岩石的科学认识。有些学者认为, 生物岩的形成经历了成岩作用的改造, 所以, 应该放到沉积岩中。在遭受成岩作用改造之前, 已经自成岩石; 成岩改造不是生物岩形成岩石所必须的。所以, 生物岩还是应当自成一类。当然, 生物岩的研究离不开沉积学和沉积岩石学知识和方法的应用, 所以, 沉积学和沉积岩石学都是生物岩石学的相关学科。
将所有生物成因的岩石归集到生物岩名下不只是一个认识的进步, 而且使人认识到, 生物岩不仅数量巨大, 而且种类很多, 亦彰显生物岩研究的重要性和生物岩石学学科的重要性。一个全面、系统、科学的分类, 是生物岩研究所必需的理论基础之一。作者的新分类, 不仅包括了传统的生物礁岩和微生物岩类型, 而且增加了17种新类型和新厘定类型, 形成了一个比较完善的新分类系统。
在宏体生物形成的宏生物岩中, 除了包括以往研究较多的生物礁岩类型外, 还包括倒骨岩、生物角砾岩、生物漂砾岩等以往关注较少的类型; 而绑结岩与粘结岩2个中文术语的澄清, 有助于今后更准确地使用它们。生屑岩首次被归入生物岩中; 微生物岩与微生物岩石2个术语首次明确加以区分; 首次定义了微礁岩和微屑岩2个术语, 使微生物岩石的术语更加明晰。
新分类系统不仅可以避免以往一种岩石被不同学者归入不同类型的问题, 而且使今后的生物岩研究有了更科学的依据, 应当能对今后生物岩的研究起促进作用。
致谢 感谢时志强、田景春、关平3位教授的有益讨论, 感谢审稿人的宝贵修改意见。
(责任编辑 郑秀娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|