


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
黔西水城长兴期晚期生物与环境突变响应: 基于古生物学和地球化学证据*
[党志英1, 2  , 沈玉林
, 沈玉林1, 2 , 杨天洋1, 2 , 金军1, 2, 3 , 赵勇4 ]
, 沈玉林, 杨天洋|
|

第一作者简介 党志英,女,1994年生,博士研究生,研究方向为沉积学、古生物地层学。E-mail: zydanghpu@126.com。
微体化石和地球化学数据的综合应用在重建古环境的研究中发挥着重要的作用。黔西水城地区上二叠统长兴组可见钙质藻类、有孔虫、腕足类、腹足类、双壳类、海百合、介形虫和海胆等化石,以钙质藻类和浮游有孔虫最为丰富多样。基于古生物分布特征,结合全岩微量元素(Rb、Cu、Sr/Ba、V/(V+Ni)、Ni/Co、V/Cr、Zr/Rb、(Zr+Rb)/Sr和Rb/Sr)和生物壳体的微量元素(Mg/Ca、Sr/Ca、B/Ca、Fe/Mn和87Sr/86Sr)分析结果,发现研究区长兴期晚期生物对环境演化较为敏感,Hindeodus changxingensis-Clarkina zhejiangensis带(长兴期晚期,约251.9 Ma)的古气候和生物演化快速变化,短时间内发生了生物数量锐减至消失、水体盐度剧增、气温突然升高以及从海退转为快速海侵的一系列变化。推测在二叠纪末生物大灭绝事件中,短期的、快速的和极端的多重相互作用(盐度、温度和快速的海侵事件等)产生的环境突变可能是当时海洋生物遭受沉重打击的主要诱因。
About the first author DANG Zhiying,born in 1994,Ph.D.candidate, is currently engaged in sedimentology and paleobiostratigraphy. E-mail: zydanghpu@126.com.
The comprehensive application of microfossils and geochemistry plays an important role in the reconstruction of paleoenvironments. Abundant biological fossils,such as calcareous algae,foraminifers,brachiopods,gastropods,bivalves,crinoids,ostracods,and echinoids,were developed in the marine carbonates of the Late Permian Changhsingian in the Shuicheng area of western Guizhou,and calcareous algae and foraminifers are the most abundant species. In this paper,the distribution characteristics of fossils,whole-rock trace elements(Rb,Cu,Sr/Ba,V/(V+Ni),Ni/Co,V/Cr,Zr/Rb,(Zr+Rb)/Sr,and Rb/Sr)and trace element analysis of biogenic shells(Mg/Ca,Sr/Ca,B/Ca,Fe/Mn and87Sr/86Sr)were studied. It was found that biological evolution was sensitive to paleoclimate variation. During the Hindeodus changxingensis-Clarkina zhejiangensis( ca. 251.9 Ma),there was a rapid variation in paleoclimate and marine organisms,and biotic groups declined sharply to disappear in a short time. Meanwhile,a series of variations occurred,including a dramatic increase in water salinity,sudden warming,and a shift from regression to a rapid transgression process. It was speculated that the interactions among salinity,temperature and rapid transgression-regression events led to climate change,which was the main reason for the extinction of marine life in the Late Permian.
二叠纪发生了一系列全球性的重大生物演化与环境剧变事件。在二叠纪中期和二叠纪末期, 就分别发生了大规模的火山喷发(Zhou et al., 2002; Ivanov et al., 2013)、海退(Haq and Schutter, 2008; Qiu et al., 2014; Qu et al., 2021)和生物灭绝事件(Stanley and Yang, 1994; Wang and Sugiyama, 2000)等全球性生物—环境事件。目前, 二叠纪末生物大灭绝事件之前潜在的生物和环境变化也受到广泛的关注(Wu et al., 2007; Black et al., 2018; Schobben et al., 2020; Hü lse et al., 2021; Yang et al., 2022a), 学者们尝试从地壳活动、大气圈、水圈、沉积圈和生物圈(包括各类生物自身演化的因素)相互作用的视角出发, 探索二叠纪—三叠纪之交的生态系统演变(殷鸿福和宋海军, 2013)。
生物化石(实体化石和遗迹化石)是地层中最原始、最直接的产物, 为地层的对比和划分及古环境和古气候分析等提供了重要的依据(Leonova, 2018; Ding et al., 2021; 张立军等, 2021; Zheng et al., 2022)。全岩样品元素地球化学分析为古生代演替研究的有力工具, 特别是在生物事件层的古海洋和古气候研究中已引起了越来越多的关注(Marynowski et al., 2012; Kumpan et al., 2015; Carmichael et al., 2016, 2019; Yang et al., 2022b)。此外, 也可利用双壳类、头足类和腕足类等生物壳体的微量元素含量和相关参数(如,
黔西地区二叠纪—三叠纪陆相、海陆过渡相和海相地层完整连续, 含丰富的动植物化石, 是研究二叠纪—三叠纪地层的理想之地。从20世纪80年代起, 很多学者对此进行了探索且成果颇丰, 例如, 对黔西晚二叠世含煤地层的矿物学和地球化学特征及其异常富集层的成因机制(徐彬彬和何明德, 2003; 邵龙义等, 2016; Shen et al., 2016b, 2019; Qin et al., 2018; 赵勇, 2021)、古生物群落(范嘉松和吴亚生, 2002; 徐彬彬和何明德, 2003; Payne et al., 2011; 杨利蓉等, 2016; Jia and Song, 2018; Liu et al., 2020; 曹怡然等, 2022)的研究, 均为古中生代之交生物大灭绝事件的研究提供了重要的科学依据。在前人研究成果的基础上, 笔者综合运用黔西水城地区乐平统长兴组K576井古生物学和地球化学数据, 对长兴期晚期的区域构造背景、气候环境、海平面变化、海洋地球化学特征以及生物演替等进行一系列分析, 以从多个角度探索二叠纪末期海洋生物面对极端气候突变时的响应。
二叠纪正是世界性泛大陆解体的时期, 南方冈瓦纳大陆和北方大陆之间的特提斯构造发生了巨大的右旋剪切拉张活动(Scotese, 2009)。在该构造背景下, 中国西南地区广泛发育走滑断裂和拉张槽盆(陈焕疆, 1990)。
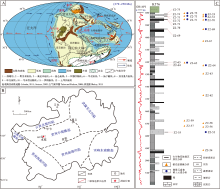
水城地区位于贵州省六盘水市, 晚古生代处于华南克拉通东南缘(图 1-A), 在大地构造上隶属于贵州中部隆起(图 1-B)。二叠纪水城地区发育紫云裂陷槽和水城—紫云断裂, 东吴运动后盆地基底构造进入相对稳定期。该时期沉积以海陆过渡相为主, 可识别出碳酸盐岩潮坪、障壁海岸和三角洲3种沉积体系(邵龙义等, 2016; Shen et al., 2016b, 2019; Qin et al., 2018)。长兴组主要以灰色、深灰色泥质粉砂岩、粉砂岩和细砂岩为主, 夹泥岩、钙质泥岩、泥灰岩和灰岩, 含煤6~20层(图 1-C), 并记录了较为丰富的植物化石及其碎片(徐彬彬和何明德, 2003)。
 | 图 1 黔西水城地区地质背景及长兴组岩性柱状图 A—全球二叠纪末期古地理图(据修改自李江海和姜洪福, 2013); B—黔西水城地区位置及区域构造单元划分(Luo et al., 2018); C—黔西水城地区K576井长兴组岩性柱状图和样品采集Fig.1 Geological background and petrological column of the Changxing Formation in in Shuicheng area, western Guizhou Province |
文中以251.9 Ma作为上二叠统顶界年龄, 将偏心率长周期(405 ka)滤波曲线作为天文谐调曲线, 建立了水城地区晚二叠世煤系天文年代标尺(赵勇, 2021)。
为揭示黔西水城地区长兴组的古生物学和地球化学特征, 笔者对水城地区K576井进行了取样(图 1-C)。样品分析和测试包括: (1)利用中国矿业大学资源与地球科学学院的OlympusBX53M显微镜对样品进行古生物学特征的观察与鉴定分析。(2)全岩微量元素含量测试在武汉上谱分析科技有限责任公司完成, 使用的仪器为电感耦合等离子体质谱仪(Agilent7700e)。(3)古生物壳体微区原位测试过程如下: 首先, 通过对光薄片磨制与显微结构观察, 选择含有古生物厚壳的区域进行微区原位分析, 其中矿物特征未经蚀变影响的厚壳部分(大于44 μm)(见图 2-R至2-T, 2-W, 绝大部分选自腕足类厚壳)可确保实验能够真实反映当时海水信息且不掺杂壳外因素的干扰; 其次, 将其置于多接收器电感耦合等离子质谱仪(MC-ICP-MSNeptunePlus)中, 在反射光下进行激光束(44 μm)剥蚀离子化测试。(4)碳酸盐岩锶同位素测试经历了溶解、分离、提纯、质谱的步骤, 选用的质量监控样品为NBS987(87Sr/86Sr=0.710241), 测试仪器为多接收器电感耦合等离子质谱仪(MC-ICP-MSNeptunePlus)(Thirlwall, 1991; Zhang and Hu, 2020)。后2项测试也均在武汉上谱分析科技有限责任公司完成。
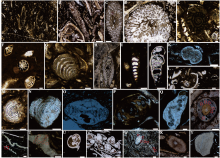
 | 图 2 黔西水城地区长兴组主要古生物类型 A-D—钙质藻类, A为 Gymnocodium bellerophontis, B和C为Permocalculus sp., D为Tauridium kurdistanensis; E-F—䗴类, E为Nanlingella sp., F为Reichelina sp.; G-Q—有孔虫类, G为 Pachyphloia schwageri, H为 Pachyphloia sp., I为 Geinitzina sp., J为 Nestellorella sp., K-L为 Howchinella sp., M和N为 Hemigordius aff. saranensis, O为 Hemigordius sp., P和Q为 Midiella sp.; R—长身贝类腕足动物棘刺; S-T—腕足碎片, 弯曲片状结构; U—腹足动物; V—双壳; W—介形虫; X—海胆刺; Y—海百合茎。比例尺: 500 μm(A, C, D, S, V, Y), 250 μm(T), 100 μm(R, T, U, W, X), 50 μm(B, E-Q)Fig.2 Main palaeontological types of the Changhsingian in Shuicheng area, western Guizhou Province |
测试样品均提前完成详细的薄片鉴定, 并进行相关的筛选, 以避免方解石脉等对实验结果产生干扰。以锶元素为例, 测试结果显示碳酸盐岩全岩样品中的锶元素平均含量为417 μg/g, 最高可至602 μg/g, 最低为285 μg/g; 古生物壳体微区原位样品中的锶元素平均含量为1088 μg/g, 最高为2990 μg/g, 最低为759 μg/g; 所有样品的锶含量均远超进行相关实验所需含量的最低值(200μg/g)(Korte et al., 2003)。相关性分析表明, 文中使用的碳酸盐岩全岩样品和微区原位样品的87Sr/86Sr值与Sr元素含量之间相关系数分别为0.0871和0.0209, 远小于0.3, 因而从锶元素含量的角度而言, 所选择的测试样品都可满足锶同位素分析的实验要求。
通过对贵州省水城地区长兴组剖面进行岩心采样、磨制薄片和镜下观察, 笔者发现了较为丰富的生物碎屑, 如钙质藻类、有孔虫、腕足类、腹足类、双壳类、海百合、介形虫、海胆(图 2)以及一些不确定的生物化石。
1)钙质藻类(图 2-A至2-D)。钙质藻类不仅是二叠纪浅海环境中的主要生产者, 而且也维持着浅层底栖生态系统的繁盛。主要成分是高镁方解石, 钙化强烈时以粒晶结构为主。形态保存完好的相对较多, 多呈梨形和卵形, 周围破碎的条带状藻类也极为丰富。
2)原生动物(图 2-E至2-Q)。常见的化石主要是有孔虫(包括䗴)。有孔虫个体微小, 多为0.05~1 mm, 因体小质轻, 不易破碎, 所以薄片中有孔虫形态大多保存完好。钙质壳, 主要成分是低镁方解石, 以隐粒和微粒结构为主。䗴类在研究区也有个别产出, 薄片中大小为0.5 mm左右, 可见纺锤状与椭圆状。
3)腕足类(图 2-R, 2-S, 2-T)。腕足类适应环境能力较强, 在研究区产出广泛。其多以碎屑形式产出, 形态不规则, 大多呈一定弧度的弯曲状。碎片长度多在0.5~4 mm之间, 主要呈弯曲状和波状, 镜下最常见的是弯曲片状结构。也可见腕足类棘刺, 其横切面呈椭圆形。
4)软体动物门(图 2-U, 2-V)。腹足纲、瓣鳃纲多见。广盐度的腹足类较丰富, 碎片大小为1~3 mm, 镜下多见粒状显微结构。瓣鳃纲又被称为双壳纲, 在研究区同样较为丰富, 碎片大小为0.5~2 mm, 多呈直片状、柱状, 且厚度一致, 显微结构为晶粒结构。通常, 瓣鳃类主要营底栖固着、爬行、钻穴和寄生等, 可与腕足类碎片共生。
5)棘皮动物(图 2-X, 2-Y)。见单晶结构的海百合茎和海胆。棘皮类生物为正常的窄盐度浅海生物, 镜下多观察到其茎的化石, 直径一般为0.3 mm左右, 呈圆形(横切面)与椭圆形(斜切面)。
6)节肢动物(图 2-W)。以介形虫为主, 粒径大小为0.5~1 mm, 壳体主要是低镁方解石, 具玻纤结构, 以圆形和椭圆形为主。研究区中介形虫的2个瓣壳多数是大小相等的, 同时也见双瓣壳周围存在压扁的单瓣壳。
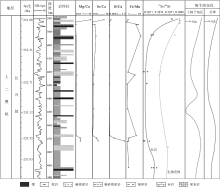
基于上述生物碎屑特征, 笔者进一步较为细致地整理了生物碎屑的分布特征、分布频率和粒径大小(仅针对产出较为连续的有孔虫、轮廓相对典型的腕足类和双壳类的壳体碎片)(图 3)。结果表明, 研究区以钙质藻类和浮游有孔虫最为丰富多样, 其中钙质藻类主要包括Gymnocodium和Permoclculus, 有孔虫则主要是Nestellorella, 其次是Reichelina(䗴类)。此外, 有孔虫、腕足类和双壳类生物壳体的出现频率相较其他古生物而言更具有延续性。晚二叠世早、中期物种丰度较大, 晚二叠世末期生物出现短暂的高分异度(样品Z2-70至Z2-71), 随之物种快速消减, 而介形虫数量增加、呈集群分布(样品Z2-75)(图 3)。生物的粒径大小在晚二叠世末期呈现出明显的波动下降趋势, 但粒径较小的双壳类则相对稳定, 且波动性远远小于粒径较大的有孔虫和腕足类(图 3)。
 | 图 3 黔西水城地区长兴组的古生物分布特征以及环境变化(火山活动、磁极性带、牙形石带、古气候、 |
文中测定了黔西水城地区长兴组碳酸盐岩中9个样品微量元素(表 1)的平均值, 并与上地壳微量元素含量进行了对比(图 4-A)。可以看出, Rb、Cs、Ba、Tl 、Pb相对亏损(CC< 0.5), Nb相对富集(2< CC< 5), Li、Be、Sc、V、Cr、Co、Ni、Cu、Zn、Ga、Sr、Zr、Sn、Hf、Ta、Th、U含量都相对接近于上地壳含量(0.5< CC< 2)。
| 表 1 黔西水城地区长兴组全岩微量元素浓度(μg/g) Table 1 Whole-rock trace element concentration (μg/g)of the Changxing Formation in Shuicheng area, western Guizhou Province |
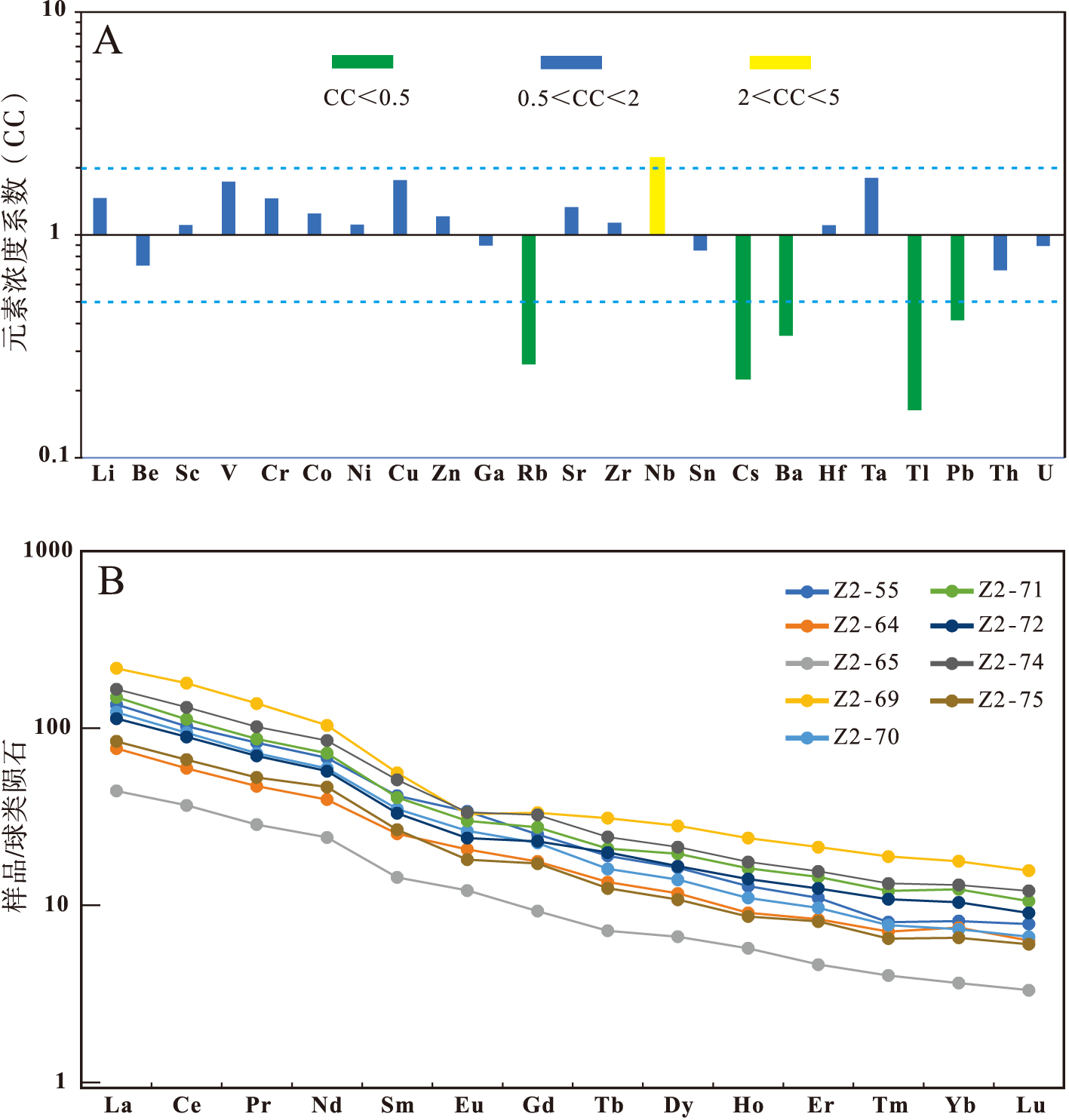 | 图 4 黔西水城地区长兴组微量元素富集系数(A)和稀土元素分布模式(B)Fig.4 Trace element concentration coefficients(A), and chondrite-normalized REE patterns(B)of samples of the Changxing Formation in Shuicheng area, western Guizhou Province |
黔西水城地区长兴组碳酸盐岩样品中的稀土元素总量为81.52~377.63 μg/g, 平均值为193.90 μg/g(表 2)。以球类陨石的测定值对样品稀土元素测试结果进行标准化, 如图 4-B所示, 稀土元素具现出LREE富集、HREE相对亏损、Eu微弱负异常或正常的特征。
| 表 2 黔西水城地区长兴组稀土微量元素浓度(μg/g) Table 2 REEs concentration(μg/g) of the Changxing Formation in Shuicheng area, western Guizhou Province |
笔者挑选了30处原生形态保存良好, 表面无明显蚀变的腕足类、有孔虫、介形虫等生物壳体进行微区原位微量元素分析, 此处选取了其中(腕足类介壳约占70%)蚀变程度最低的数据点(表 3)。可以看出:B含量急剧下降, 在长兴期早、中、晚期差异明显; Mg含量在长兴期早期和中期具2个较大的峰值, 在长兴期晚期急剧上升; Ca含量前期相对平稳, 在长兴期晚期出现峰值; Mn含量波动较单一, 仅在中期和晚期出现2个峰值; Fe和Sr含量最不稳定, 出现多个峰值。
| 表 3 黔西水城地区长兴组古生物壳体微量元素分析结果(CaCO3, wt.%; B, Mg, Mn, Fe, Ga和Sr, μg/g) Table 3 In-situ-LA-ICP-MS trace elemental analyses of biogenic shells of the Changxing Formation in Shuicheng area, western Guizhou Province(CaCO3, wt.%; B, Mg, Mn, Fe, Ga and Sr, μg/g) |
本次测试的锶同位素共获得20个数据, 平均值为0.707399, 其中小于0.7072的样品占40%(8个)、介于0.7072~0.7074之间的样品占25%(5个)、介于0.7074~0.7077之间的样品占10%(2个)、介于0.7077~0.7079之间的样品占10%(2个)、大于0.7079的样品占15%(3个)(表 4)。总体而言, 锶同位素在长兴期早期缓慢下降, 至长兴期中晚期达到最低值0.707053, 随后开始逐渐上升, 至长兴期晚期从0.707163迅速升至0.707611, 平均增长速率达0.0004/10 m。
| 表 4 黔西水城地区长兴组碳酸盐岩和古生物微区原位样品锶同位素组成统计 Table 4 Statistics of strontium isotopic composition of the studied samples in carbonate rock and biogenic shells of the Changxing Formation in Shuicheng area, western Guizhou Province |
沉积岩中的微量元素作为研究沉积环境的重要媒介, 经常使用敏感元素(即微量元素)之间的关系或比值进行比较。文中利用K576井长兴组全岩微量及稀土元素特征进行了一系列分析(图 5)。
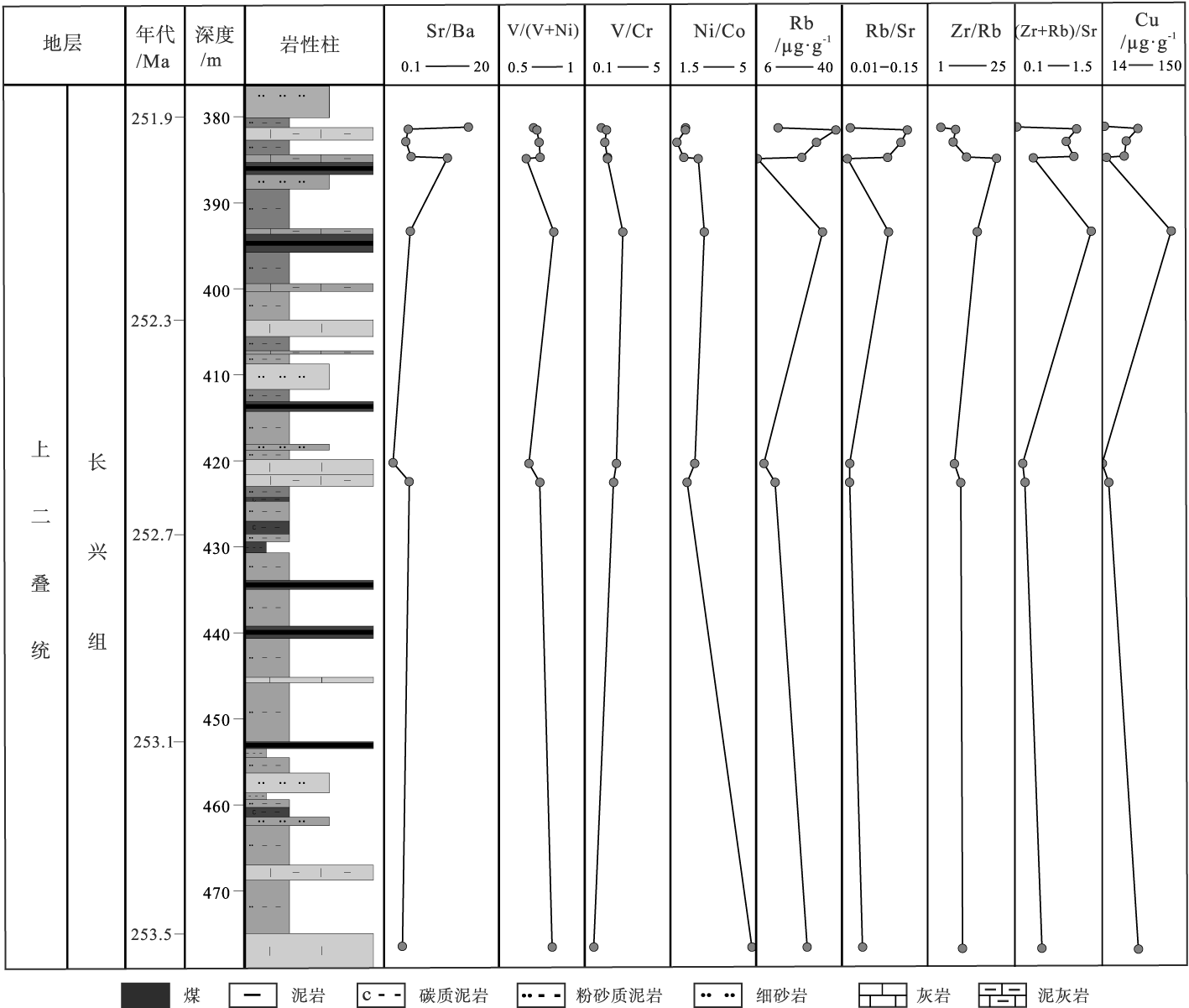 | 图 5 黔西水城地区K576井长兴组全岩微量及稀土元素特征垂向变化曲线Fig.5 Stratigraphic distribution of whole-rock trace elements and REEs of the Changxing Formation from Well K576 in Shuicheng area, western Guizhou Province |
Sr/Ba对水介质古盐度判识具有一定的指示意义, 其中Sr/Ba值大于1表示咸水环境, 0.6~1为半咸水环境, 小于0.6为淡水环境(Johnsson and Basu, 1993)。研究区Sr/Ba平均值4.69, 长兴组上部(样品Z2-75)高达14.32, 表明在长兴期晚期处于咸水环境, 且水体异常、存在盐度突然增加的现象。尤其值得关注的是样品Z2-75, 其位于灰岩层段, 表明该阶段也伴随着海侵事件。
V/(V+Ni)值小于0.45表征氧化环境, 0.45~0.6之间表征贫氧环境, 0.6~0.85之间表征缺氧环境, 大于0.85表征硫化环境(Hatch and Leventhal, 1992)。同时, V/(V+Ni)值也可用来判断沉积物沉积时底层水体分层的强弱, 大于0.84分层强, 0.6~0.84分层中等, 0.4~0.6分层弱(Hatch and Leventhal, 1992; 熊国庆等, 2008)。V/Cr值小于2表征氧化环境, 大于4.25表征还原环境, 2~4.25之间表征过渡环境(Jones and Manning, 1994)。Ni/Co值小于5表征氧化环境, 大于7表征还原环境, 5~7之间表征过渡环境(Jones and Manning, 1994)。研究区V/Cr值平均为1.40, 为氧化环境特征; Ni/Co值平均为2.53, 显示氧化环境特征; V/(V+Ni)平均值为0.73, 显示了分层中等的还原环境特征, 但其元素比值呈持续降低状态, 表明逐渐向氧化环境靠近, 因此其与上述2种微量元素比值指示的环境特征不符, 有可能是钒元素含量低造成的。总体分析认为, 长兴期研究区以较为稳定的氧化环境为主。
Zr/Rb值越低, 水动力越弱, 水位越高(Jin et al., 2006)。(Rb+Zr)/Sr值越低水体深度相对较大(Mü ller and Suess, 1979)。研究区Zr/Rb和(Zr+Rb)/Sr值分别分布在19.84~4.29之间和1.33~0.12之间, 变化趋势大致相同, 均呈现出波动式的下降, 在长兴期晚期(样品Z2-75)变化明显, 并快速达到最低值, 说明该时期研究区水体较深、水动力条件弱且海平面快速上升。
综合上述分析, 认为黔西水城地区长兴期整体上以稳定的氧化环境为主, 但是晚期环境发生显著的变化, 海平面快速上升, 海水咸化程度突增, 在样品Z2-75处高达14.32(表 1)。
Rb/Sr值是判别古气候环境的敏感指标(Ouyang et al., 2019)。Sr和Rb指示潮湿气候条件, Rb/Sr值高通常说明气候温暖湿润, 反之气候相对干旱炎热(Ouyang et al., 2019)。研究区Rb/Sr值分布在0.02~0.12之间, 二者的变化曲线基本趋于一致, 在长兴期晚期最低值出现之前, 数值波动相对较小(图 5), 反映了长兴期整体上以温暖潮湿的气候为主, 在长兴期晚期(样品Z2-74至Z2-75)气候突然快速炎热和干旱化。
从古生物的分布特征中也能发现气候变化端倪。在长兴组顶部生物多样性急剧下降, 仅残留了一定数量的介形虫和极少数量的腕足动物碎片(样品Z2-72至Z2-74), 其中介形虫呈集群分布、数量颇丰(样品Z2-75)(图 2-T; 图 3)。研究发现, 二叠纪晚期—三叠纪早期一些介形虫适应了缺氧的条件, 且多次在灭绝层位前后出现(Crasquin-Soleau and Kershaw, 2005; Wu et al., 2007)。Song等(2014)统计了关于缺氧和高温的文献资料, 对海洋生物进行实验评估, 发现介形虫的温度上限高于有孔虫、棘皮类、腹足类、非介形类甲壳类动物。因此, 在面临环境突变的时候, 不耐高温的浅水群落珊瑚、大型有孔虫、放射虫率先衰减, 而耐高温的介形虫则在缺氧环境中相对繁盛。此外, 相较于晚二叠世早、中期, 晚二叠世末期物种呈现短暂的高分异度(样品Z2-70至Z2-71)后快速消减, 直至消失不见, 壳体粒径趋于变小。这可能是生物对环境恶化作出的响应。
化石壳体微量元素和稳定同位素对恢复古环境和古气候等具有重要作用(Holmden and Hudson, 2003; Immenhauser et al., 2016)。在利用生物壳体重建古海洋环境中, Mg/Ca值用于重建底层海水温度, 因为壳体中的Mg/Ca值与海水温度呈正相关(Lear et al., 2000; Scheiner et al., 2018)。Sr/Ca值可当作海水古盐度的一个指标(Holmes, 1992), B/Ca值用于建立海水的碳酸盐化学信息(此处缺失 Δ[
研究区Mg/Ca、Sr/Ca、B/Ca和87Sr/86Sr值的变化趋势基本一致(图 6), 均在长兴期晚期呈明显的波动, 且急剧上升, 这表明在长兴期中晚期生物生活的海洋环境相对稳定, 直至长兴期晚期最后时段发生突变。例如, 长兴组顶部Z2-75样品记录了Mg/Ca值由之前的0.04~9.48突增至30.87, 指示了古海水温度的突然升高; Sr/Ca值由0.06~3.36急增至6.85, 可能指示了沉积水体的急剧咸化; 87Sr/86Sr值突增代表了海平面快速上升等。值得注意的是, 通常87Sr/86Sr值与海平面的升降呈现负相关关系(Butterfield et al., 2001; Das et al., 2006), 但长兴期晚期锶同位素比值与海平面呈正相关关系, 并与国际上公布的结果相符(McArthur et al., 2012; Ye et al., 2015)。Fe/Mn值曲线前段宽缓波动, 尾端快速下降, 表明长兴期早、中期生物经历了大规模的海退期, 而在长兴期晚期发生了快速海侵, 这与87Sr/86Sr比值的变化结果趋于一致。
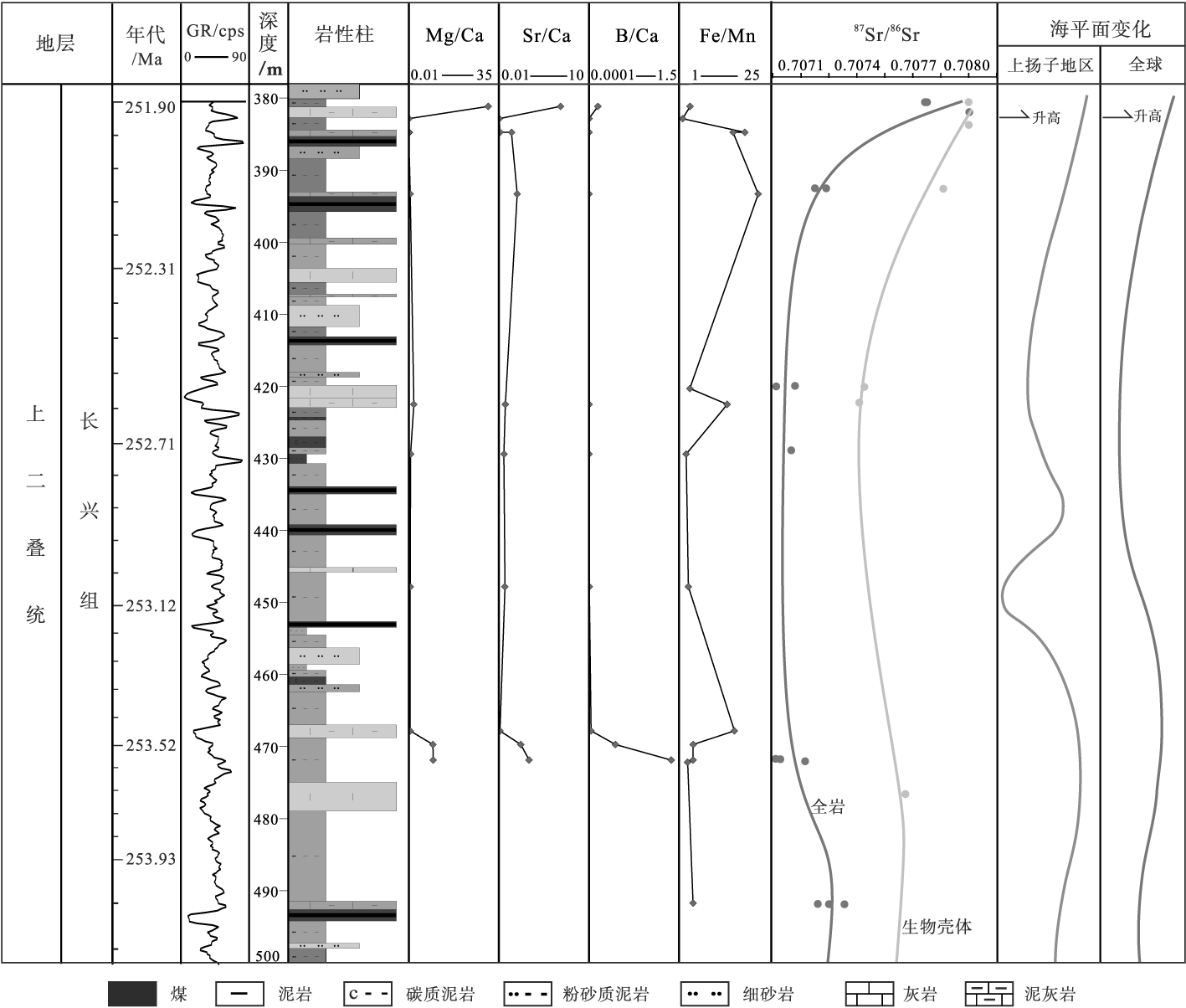 | 图 6 黔西水城地区K576井长兴组古生物壳体微量元素特征垂向变化曲线(Mg/Ca, Sr/Ca, B/Ca, and Fe/Mn, mmol/mol×10-2)Fig.6 In-situ LA-ICP-MS trace elemental analyses of biogenic shells of the Changxing Formation from Well 576 in Shuicheng area, weatern Guizhou Province(Mg/Ca, Sr/Ca, B/Ca, and Fe/Mn, mmol/mol×10-2) |
基于影响深远的二叠纪末期生物大灭绝事件背景, 二叠纪—三叠纪之交的全球变化和生物大灭绝被认为是地球各圈层相互作用的产物, 是一个从渐变到突变的长期过程(殷鸿福和宋海军, 2013)。中晚二叠世西伯利亚玄武岩和华南大规模的火山喷发导致地表环境发生剧烈变化, 在中国南方川滇黔地区以峨眉山玄武岩喷发为代表, 主要由大量的基性玄武质火山岩系和同时期的侵入体构成, 包括少量火山碎屑岩及超基性岩等(Xu et al., 2001; Xiao et al., 2004; Ali et al., 2010; Yang et al., 2021)。Zhang等(2021)在华南地区发现了极端的Cu和Hg富集, 且认为华南地块周围的大型长英质火山活动是其物质来源。火山爆发向平流层注入含硫气体是气候变化最重要的驱动因素之一(Hegerl et al., 2003; Sigl et al., 2015), 在气候变暖的情况中, 随着大规模火山喷发平流层变暖、对流层冷却和地表冷却加剧, 喷发柱力学和SO2注入高度的变化促进了这种温度响应的放大(Aubry et al., 2021), 从而引发了“火山冬天效应”, 即在二叠纪末期古温度呈现出快速降温, 随后突然变暖(Campbell et al., 1992; Renne et al., 1995; He et al., 2014)。这种温度变化趋势又与同期海平面下降、随后立即发生海侵相一致(Shen S.Z et al., 2006; 沈树忠等, 2019; Zhang et al., 2021)。因此, 爆炸性火山活动致使的气候快速变化是导致环境退化的主要驱动因素之一。长兴期晚期研究区物源的构造背景复杂, 在Clarkina yini带Cu含量(样品Z2-69)极为丰富处(表 1)生物数量和多样性低(图 3), 且(Zr+Rb)/Sr值达到最高值(图 5), 与海平面下降相对应。由此推测, 此时可能正处于全球气候变凉时期。
笔者发现钙质藻类与有孔虫在生态学上具有一定程度的相关性, 钙质藻类随着有孔虫的大幅减少而同步消失, 但是生物并未伴随着碳同位素的负偏移而衰退(图 3)。例如, 在Clarkina changxingensis带和Clarkinayini带有孔虫锐减, 同期钙质藻类基本不可见, 生物生产力降低。而在Hindeodus changxingensis-Clarkina zhejiangensis带则出现了高度多样性的动物群, 直至生物突然衰退, 大多数类群消失。这一现象支持了钙质藻类在维持浅层底栖生态系统方面的重要作用, 但不支持
在Hindeodus changxingensis-Clarkina zhejiangensis带的下段, 生物多样性异常丰富, 可见钙质藻类、有孔虫、腹足类、海百合茎、海胆刺、腕足类和双壳类等(图 3)。其中, 裸海松藻科Permocalculus破碎的叶状体随处可见, 喜暖水的䗴类有孔虫物种增多, 见Nankinella和Reichelina, 且小壳体的Reichelina较为丰富。在Hindeodus changxingensis-Clarkina zhejiangensis带的上段(接近Hindeodus parvus带), 下段的生物突然消失, 留下了相对耐受的腕足类、双壳类和介形虫, 尤其是耐高温的介形虫集群出现, 支持了极端高温事件的出现(图 3)。此外, 一些学者认为䗴类有孔虫演化最明显的转折点与高环境应力时期(如全球海侵—海退循环)可能相关, 是其与盐度、温度、生存空间等各种多重因素相互作用的后续效应(Stanley and Yang, 1994; Isozaki and Aljinović , 2009; Hada et al., 2015)。文中数据也很好地证明了这一观点。
水城地区长兴组地层全岩和生物壳体微量元素在Hindeodus changxingensis-Clarkina zhejiangensis带变化剧烈, 呈现出水体盐度剧增(Sr/Ba和Sr/Ca值增加)、突然升温(Rb减少、Rb/Sr值减少和Mg/Ca值增加)以及从海退转入快速海侵(Zr/Rb和(Zr+Rb)/Sr值减少、Fe/Mn和87Sr/86Sr值增加)的一系列变化(图 5; 图 6)。值得关注的是, 壳体硼浓度在长兴期早期高达418 μg/g, 之后开始急剧下降, 仅有1次高于100 μg/g, 晚期又降低至26 μg/g(表 3), 而不同碳酸盐岩样品(壳体)中硼浓度的变化可能是钙化环境和
此外, 海退后的迅速海侵与温室气候相结合, 则有利于分层、缺氧海洋的形成, 而从深海到浅海广泛出现的缺氧事件被认为是二叠纪末生物大灭绝的主要原因(Wignall and Twitchett, 1996, 2002; Racki and Wignall, 2005; Song et al., 2014; Shen et al., 2016)。然而, 在水城地区长兴期V/(V+Ni)、Ni/Co和V/Cr值波动稳定(图 5), 以稳定的氧化环境特征为主, 可见该时期生物的锐减至消失并不是缺氧事件为主要诱因造成的。Sagasti 等(2000)曾率先进行过缺氧对表栖群落影响的研究, 结果表明表栖动物不一定比内栖动物更容易受到氧胁迫, 一些优势种似乎对缺氧存在很高的耐受性。同时, 表栖群落可能会保持相对不变的物种组成, 但随着氧浓度下降, 其丰度会发生一些变化。此外, 张立军等(2021)关于二叠纪—三叠纪之交深水相遗迹群落的研究也发现长兴期和印度期出现了高度多样性的动物群落, 似乎缺氧并未对生物造成极大的影响。由此推测, 缺氧可能并不是二叠纪末生物大灭绝的主要原因(Kakuwa, 1996; Haas et al., 2007), 生物自身也可能在不断增强对缺氧环境的适应能力和耐受能力。
黔西水城地区上二叠统长兴组海相碳酸盐岩发育较好, 运用系统的古生物学和地球化学分析对区内长兴组沉积末期的水介质条件、古气候和古海洋进行研究, 发现长兴期晚期古环境面貌以温暖湿润气候、海陆过渡相沉积环境和氧化环境为主, 温水类群钙质藻类和有孔虫较为丰富。然而, 在Hindeodus changxingensis-Clarkina zhejiangensis带古气候和生物演化迅速变化, 短时间内发生了生物锐减至消失、水体盐度剧增、突然升温以及从海退转入快速海侵的一系列变化。
推测短时期、快速的气候变化对浅海生物是一个更为沉重的打击, 因为一般在极短的时间尺度下, 大多数生物无法快速地应对极端气候而快速消亡, 只有极个别生物才能生存, 这也可能是二叠纪末期生物大灭绝的最重要原因之一。
(责任编辑 张西娟; 英文审校 刘贺娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|