


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
云南保山红岩剖面诺利期牙形石 Mockina slovakensis的发现及其对沉积环境的指示*
[郭豹1  , 杜怡星
, 杜怡星1, 2 , 时志强1, 2 , 金鑫1, 2 ]
, 杜怡星, 时志强|
|

第一作者简介 郭豹,男,1999年生,成都理工大学沉积地质研究院硕士研究生,主要研究方向为古生物学和沉积学。E-mail: 1539597169@qq.com。
Mockina slovakensis是晚三叠世诺利期一个重要的牙形石属种,除了具有重要的地层学作用之外,它还能够指示特殊的沉积环境。本次研究在云南保山红岩剖面上三叠统大水塘组下段灰岩中发现了大量牙形石M. slovakensis,其在地层中占据统治地位,据此将研究层位的时代限定为晚三叠世诺利期 Alaunian晚期。有记录表明,当M. slovakensis在地层中占据统治地位时,往往指示出一种较为局限的沉积环境。综合此次研究在保山红岩剖面发现的大量占统治地位的M. slovakensis牙形石以及前人对保山地块大水塘组沉积环境研究的结果,推测在诺利期 Alaunian晚期,保山地区应为拉张背景下形成的较为局限的沉积盆地。该研究成果有助于进一步了解晚三叠世特提斯东部的沉积环境演化。
About the first author GUO Bao,born in 1999,is a master degree student in Institute of Sedimentary Geology,Chengdu University of Technology. He is engaged in research on paleontology and sedimentology. E-mail: 1539597169@qq.com.
Mockina slovakensis is one of the important species of conodont in Norian period. It plays a significant role in stratigraphy and acts as an indication for certain distinctive sedimentary environments. In this study,a significant abundance of conodont species M. slovakensis was discovered in the limestone deposits located in the lower parts of the Dashuitang Formation in the Hongyan section of Baoshan,Yunnan. Based on the conodont biostratigraphy,the age of the sedimentary strata in this section has been constrained to the late Alaunian of Norian period. Its presence and abundance can provide valuable insights into the paleoenvironmental conditions and depositional settings. M. slovakensis is known for strong adaptability to a diverse living environments,including both shallow and deep oceanic environment,as well as restricted sea to open sea. However,various records indicate that M. slovakensis conodont indicates the restricted depositional environment. Considering the abundance of dominant M. slovakensis found in the Hongyan section of Baoshan,combined with previous studies on the sedimentary environment of the Dashuitang Formation in the Baoshan block,it is reasonable to conclude that the Baoshan area formed a restricted sedimentary basin during the Alaunian stage under a tectonic stretching setting. The results are helpful for understanding the evolution of the sedimentary environment during the Late Triassic period in eastern Tethys.
牙形石是一类已灭绝动物的口咽部位的磷酸盐岩硬体结构, 通常广泛分布于寒武纪至三叠纪的海相地层中, 极少数被发现于侏罗纪初期的地层中(Pá lfy et al., 2007; Du et al., 2020b, 2023)。牙形石因其演化迅速、易于获取、分布广泛等特点, 常被作为全球海相地层划分和对比的重要标志(张再天, 2016; 杨志华, 2016; Rigo et al., 2018; 吴强旺等, 2022)。尤其是在上三叠统地层划分中, 牙形石扮演了重要的角色, 比如卡尼阶和诺利阶、诺利阶和瑞替阶以及各亚阶之间界线的划分等, 大都采用牙形石来定义(Rigo et al., 2018)。值得注意的是, 一些牙形石不仅具有重要的地层学意义, 还可能具有指示特殊沉积环境的作用, 比如牙形石Mockina slovakensis(Kozur, 1972)。
Mockina slovakensis是晚三叠世诺利期重要的牙形石属种, 由Kozur在1972年建立。由于其易于识别, 且在这一时期的特提斯地区广泛分布, 因此Rigo等(2018)将M. slovakensis牙形石带作为特提斯地区诺利阶Alaunian亚阶的最后一个牙形石带, M. slovakensis的出现通常指示地层时代为诺利期Alaunian晚期至Sevatian晚期。除了重要的地层学作用之外, M. slovakensis还能够指示特殊的沉积环境。Du等(2021)在总结前人的研究后发现, 虽然M. slovakensis的分布环境较为多样, 从浅水碳酸盐台地到深海环境均有分布, 但其更加偏好一种较为局限的生存环境, 因为目前的研究已发现当M. slovakensis在地层中的牙形石群体中占据主导地位时, 其生存的沉积环境均为以台内盆地(intraplatform basin)为代表的局限环境(Roghi et al., 1995; Donofrio et al., 2003; Du et al., 2021)。
目前有关M. slovakensis的研究主要集中在西特提斯洋地区, 在东特提斯洋和泛大洋地区的研究还相对较少。在中国, 关于M. slovakensis牙形石的报道主要集中在东特提斯云南保山地区: 云南省地质局区域地质调查队在1980年的1︰20万保山幅区域地质调查报告中报道了牙形石Epigondolella cf. slovakensis①(①云南省地质局区域地质调查队. 1980. 保山幅G-47-28 1/20万区域地质调查报告.); 董致中和王伟(2006)在保山大堡子剖面发现了牙形石M. slovakensis; Jin等(2022)在保山红岩剖面也报道了牙形石M. slovakensis; 曾卫平(2022)在保山喜鹊林剖面也发现并报道了该牙形石属种, 并对其形态变异进行了讨论。前人的研究主要是针对M. slovakensis的生物地层学意义, 但对其沉积环境的指示意义尚未有足够的讨论和研究。因此, 此次研究除了对牙形石M. slovakensis的形态特征、物种的分布及其地层学意义进行讨论外, 还将结合前人研究对M. slovakensis蕴含的沉积环境指示意义进行探讨分析, 最终结合前人研究的文献资料, 对晚三叠世诺利期保山地块的沉积环境演化展开讨论。
保山地块位于云南省西部, 毗邻缅甸东北部, 处于欧亚板块和印度板块的交汇地带(图 1)。保山地块在大地构造上位于滇缅泰马地块(Sibumasu)北延位置(Li et al., 2004; Sone and Metcalfe, 2008; Metcalfe, 2009; 廖世勇等, 2013; 王小端等, 2019), 处于冈瓦纳大陆的边缘, 南侧与缅甸掸邦地块相连, 北侧由于澜沧江断裂和怒江断裂在碧江一带汇拢而消失(董美玲等, 2012), 西部以怒江— 龙陵— 潞西断裂为界与腾冲地块相连, 东部以澜沧江— 柯街— 南定柯街断裂为界与昌宁— 孟连结合带相接壤(Wang et al., 2001; Du et al., 2020a)。
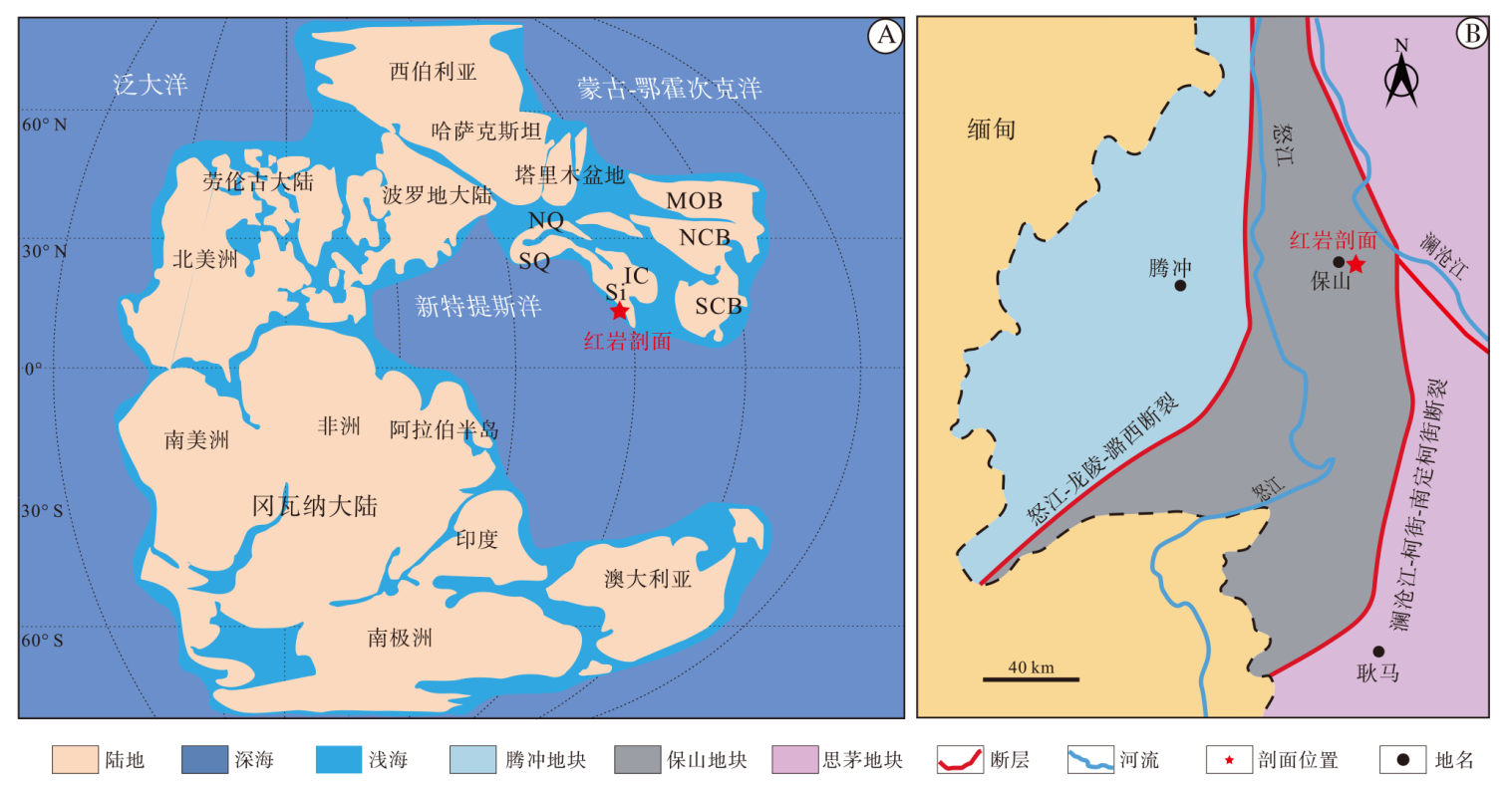 | 图 1 云南保山地区红岩剖面地理位置 A— 晚三叠世220 Ma全球古地理图(据Huang et al., 2018), 红星表示红岩剖面大致位置, Si: Sibumasu(滇缅泰马地块); IC: 中印地块; MOB: 蒙古板块; NCB: 华北板块; SCB: 华南板块; NQ: 北羌塘地块; SQ: 南羌塘地块。B— 红岩剖面地理位置(据Du et al., 2020a)Fig.1 Geographic location of Hongyan section in Baoshan area, Yunnan Province |
在晚古生代, 由于冈瓦纳大陆东北部的裂谷作用, 保山地块和其他Sibumasu-Lasa地体一起向北漂移, 古特提斯洋也在此时开始关闭, 直至晚三叠世与华南板块、华北板块等其他中国地块拼合在一起(图 1-A)(Li et al., 2015; Jin et al., 2022)。晚三叠世, 保山地区记录了2次显著的火山作用/岩浆作用事件, 一次是在早卡尼期(包佳凤等, 2020), 另一次则是在中诺利期(约216.8± 2.2 Ma, 廖世勇等, 2013)。
保山地块上三叠统由下至上可分为大水塘组、南梳坝组和弯甸坝组(Wang et al., 2001)。此次研究的保山红岩剖面位于云南省保山市以东9 km处(图 1), 由上三叠统大水塘组和南梳坝组组成, 其中大水塘组主要发育薄— 中层灰岩、生物碎屑灰岩、含硅质结核及条带灰岩, 含有少量浊积岩和泥页岩(图 2; 图 3), 与下伏河街湾组呈假整合接触①(①云南省地质局区域地质调查队. 1980. 保山幅G-47-28 1/20万区域地质调查报告.)(吴强旺等, 2020), 而南梳坝组下部主要由灰岩和泥岩组成, 上部主要由粉砂岩和砂岩组成, 仅有少量灰岩夹层。
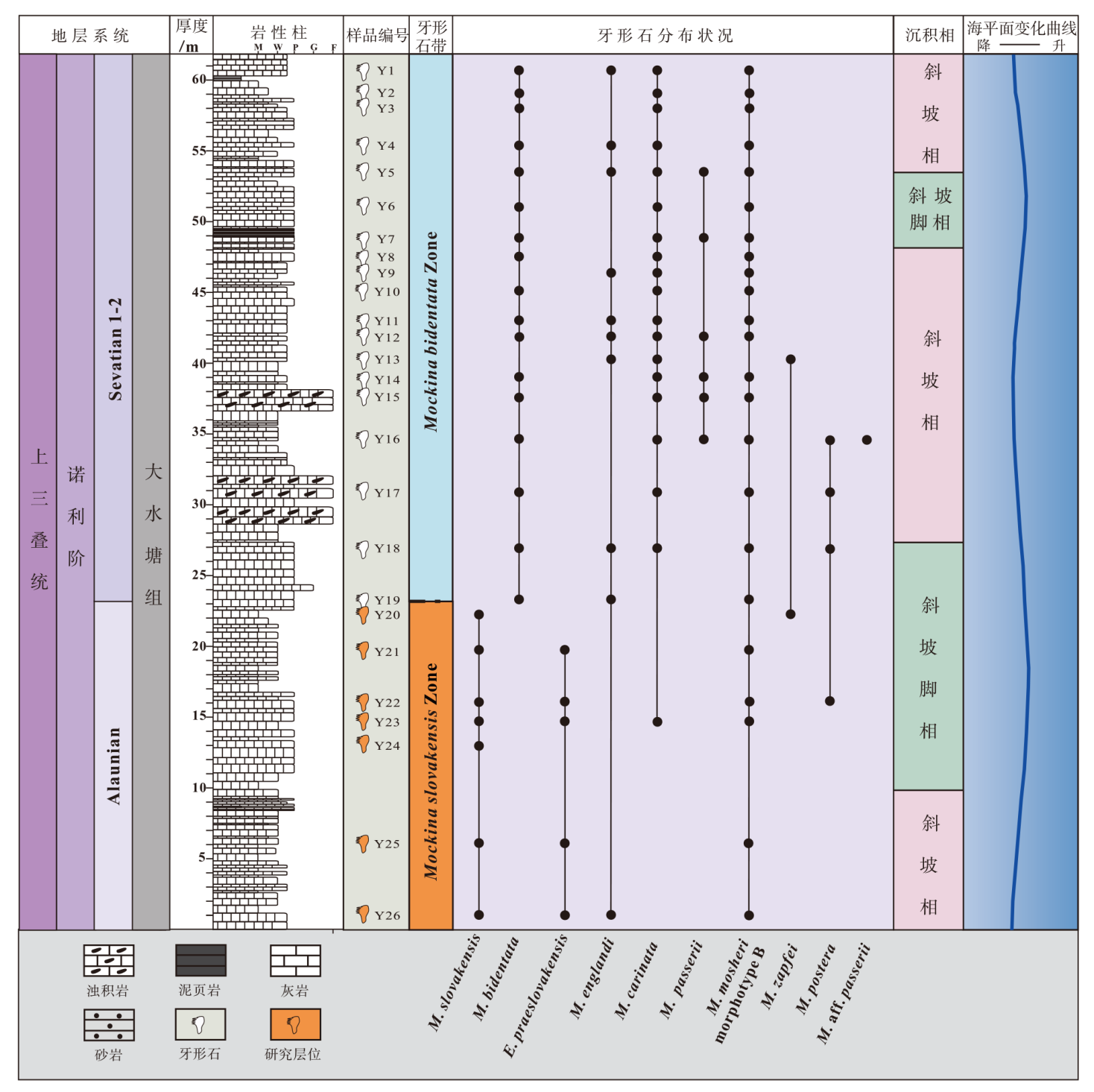 | 图 2 云南保山地区红岩剖面大水塘组综合地层柱状图 M: 泥晶灰岩; W: 粒泥灰岩; P: 泥粒灰岩; G: 颗粒灰岩; F:漂砾岩。岩性柱、牙形石带及部分牙形石分布状况据Jin 等(2022), 沉积相、海平面变化曲线据吴强旺等(2020)Fig.2 Comprehensive stratigraphic column of the Dashuitang Formation at Hongyan section of Baoshan area, Yunnan Province |
 | 图 3 云南保山地区红岩剖面野外露头照片 A— 红岩剖面厚度2 m处, 灰色薄— 中层状泥晶灰岩; B— 红岩剖面厚度13 m处, 为牙形石样品Y24取样位置, 浅灰色中— 厚层状泥粒灰岩、粒泥灰岩; C— 红岩剖面厚度20 m处, 浅灰色薄— 中层状泥晶灰岩、粒泥灰岩; D— 红岩剖面厚度32 m处, 浅灰色层状砾屑灰岩, 砾屑颗粒为垮塌角砾岩, 夹杂部分硅质团块; E— 红岩剖面厚度44 m处, 灰白色、灰色薄— 中层状粒泥灰岩, 含部分硅质条带, 水平层理发育; F— 红岩剖面厚度54 m处, 为牙形石样品Y5取样位置, 灰色、深灰色薄— 中层状泥粒灰岩Fig.3 Field outcrop photos of Hongyan section in Baoshan area, Yunnan Province |
吴强旺等(2020)对保山红岩剖面地层进行了研究, 共归纳出8种微相, 识别出斜坡相、斜坡脚相及深水陆棚相3种沉积相。微相类型表明, 该剖面自下而上为一个海平面相对上升的旋回(图 2), 对应云南保山地块晚三叠世的广泛海侵。Jin等(2022)对该剖面进行了生物地层学和稳定碳同位素的综合研究, 共识别出4次碳同位素负偏, 推测发生在中晚诺利期的2次碳同位素偏移(CIE 1和CIE 2)可能与大规模岩浆活动有关, 并建立了M. slovakensis和M. bidentata牙形石带(图 2)。
保山地块晚三叠世牙形石研究可追溯到20世纪70年代。1980年, 在1︰20万保山幅地质调查报告中首先报道了诺利期的牙形石①(① 云南省地质局区域地质调查队. 1980. 保山幅 G-47-28 1/20万区域地质调查报告。)。其后, 王志浩和董致中(1985)在保山地区开展了诺利期牙形石生物地层的专门研究, 在上三叠统大水塘组顶部和南梳坝组下部发现了牙形石“ Epigondolella” (=Mockina) postera (Kozur & Mostler, 1971)和“ Epigondolella” (=Mockina)bidentata(Mosher, 1968), 并建立了2个牙形石生物带, 将其时代分别划分为诺利中期和诺利晚期。董致中和王伟(2006)在云南东部和西部建立了2个不同的牙形石带, 在保山金鸡剖面和金鸡大平地剖面发现牙形石“ Epigondolella” bidentata, 在大堡子剖面发现Parvigondolella andrusovi(Kozur & Mock, 1972)和Misikella hernsteini (Mostler, 1967)。王小端等(2019)在云南保山大堡子地区大水塘组顶部灰岩中也报道了诺利期的牙形石。Du等(2020a)在保山红岩剖面报道了来自上三叠统南梳坝组的诺利期牙形石, 并首次在国内发现了牙形石Mockina englandi (Orchard, 1991)、Mockina carinata(Orchard, 1991)和Mockina mosheri morphotype B(Kozur & Mostler, 1971)等属种。Zeng等(2021)在保山地区南梳坝组灰岩中发现了P. andrusovi等多个属种的牙形石。Jin等(2022)在保山红岩剖面建立了牙形石M. bidentata带和M. slovakensis带(图 2)。曾卫平(2022)通过对保山地区3个剖面进行研究, 获得了丰富的晚三叠世诺利期牙形石自然群集, 重建了目前最年轻的牙形石多分子器官, 并尝试重建了相关牙形石演化谱系树。Zeng等(2023)对其先前研究的地区做了更加深入的研究, 并对其报道的牙形石形态物种进行了初步编绘。
总的来说, 前人对于保山地区上三叠统大水塘组生物地层的研究已经比较详细, 对于其时代业已比较清楚。根据前人的研究结果, 云南保山地区大水塘组的时代应为晚三叠世诺利期中— 晚期(图 2)(王小端等, 2019; 吴强旺等, 2020; Du et al., 2020a; Zeng et al., 2021, 2023; Jin et al., 2022; 曾卫平, 2022)。
依据Jin等(2022)在保山红岩剖面建立的M. bidentata和M. slovakensis牙形石带, 可以将红岩剖面分为上、下2个部分, 上部属于M. bidentata牙形石带, 时代为诺利期Sevatian早期, 而下部属于M. slovakensis牙形石带, 时代为诺利期Alaunian晚期。本次研究样品的采样位置为红岩剖面下部的大水塘组中— 薄层灰岩(图 2, 图 3, 从0到20 m), 该段地层中含有大量保存完好的牙形石。
将在野外采集的多个可能含有牙形石的灰岩样品(每个岩石样品重量约为2 kg)进行物理碎样, 然后将碎好的样品置于浓度约为10%的醋酸溶液中进行酸溶处理, 直到样品可溶部分完全溶解; 再将不溶残渣进行洗涤过筛, 将冲洗过筛后的残渣装入已编号的烧杯中, 残渣较多的样品需要进行进一步的重液分离; 之后将残渣放在约40 ℃的烘箱中进行干燥处理; 最后使用双目体式显微镜进行牙形石挑选和鉴定, 选取保存较好且具有代表性的牙形石样品进行扫描电镜(SEM)拍摄。上述实验过程均在成都理工大学完成。
本次研究在红岩剖面下段大水塘组(牙形石样品Y20-Y26)获得了大量的牙形石(图 4), 它们的色变指数CAI值为2~3。识别出的牙形石以Mockina属为主, 还有少量Epigondolella属的牙形石, 包括M. slovakensis、M. carinata、M. englandi、M. postera和E. praeslovakensis等。由于E. praeslovakensis和M. slovakensis在研究层位中同时出现(Moix et al., 2007), 且该地层中并未发现诺利晚期的标志性牙形石M. bidentata, 因此可以判断红岩剖面下部的大水塘组地层的时代为诺利期Alaunian晚期(图 2)。
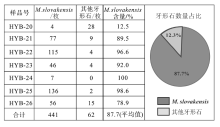
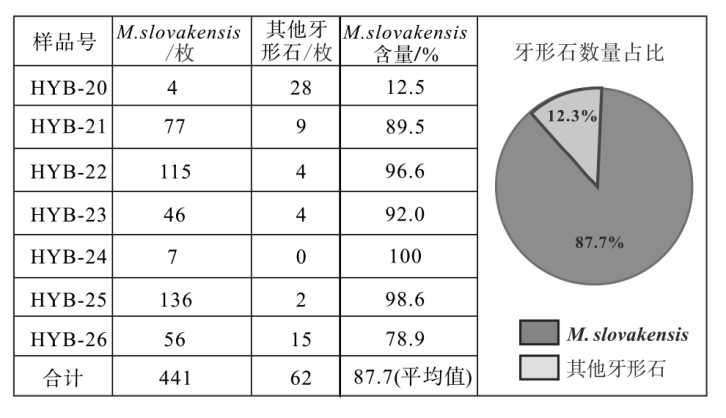 | 图 4 云南保山地区红岩剖面下段大水塘组牙形石产出数量及占比Fig.4 Quantity and proportion of conodont production in the lower part of Dashuitang Formation at Hongyan section, Baoshan area, Yunnan Province |
牙形石M. slovakensis在研究地层的牙形石生物群中占据统治地位。在样品HYB-20至HYB-26中, 共计产出牙形石503枚(不含枝型牙形石), 其中M. slovakensis占87.7%。尤其是在样品HYB-25、HYB-24、HYB-23、HYB-22中, M. slovakensis牙形石在生物群中占据绝对的优势地位。从样品HYB-21开始, M. slovakensis的占比开始下降, 在样品HYB-20中M. slovakensis已经明显失去优势地位(图 4; 图 5; 图 6)。
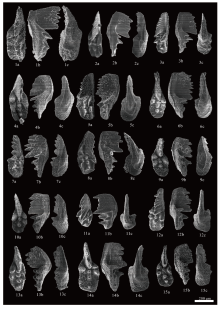
 | 图 5 云南保山红岩剖面大水塘组下段牙形石扫描电镜照片(一) 1-3— Mockina slovakensis, Y21; 4-11— M. slovakensis, Y22; 12-13— M. slovakensis, Y23; 14— Epigondolella praeslovakensis, Y23; 15— M. slovakensis, Y23。a: 口视图; b: 侧视图; c: 反口视图Fig.5 Scanning Electron Microscope(SEM)photomicrographs of conodonts from the lower part of Dashuitang Formation at Hongyan section, Baoshan area, Yunnan Province(Ⅰ ) |
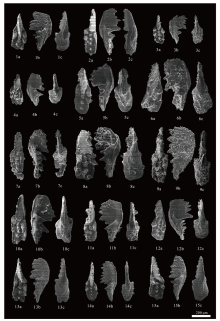
 | 图 6 云南保山红岩剖面大水塘组下段牙形石扫描电镜照片(二) 1-4— Mockina slovakensis, Y23; 5-7— M. slovakensis, Y25; 8— Epigondolella praeslovakensis, Y25; 9-11— M. slovakensis, Y25; 12-15— M. slovakensis, Y26。 a: 口视图; b: 侧视图; c: 反口视图Fig.6 Scanning Electron Microscope(SEM)photomicrographs of conodonts from the lower part of Dashuitang Formation at Hongyan section, Baoshan area, Yunnan Province(Ⅱ ) |
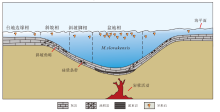
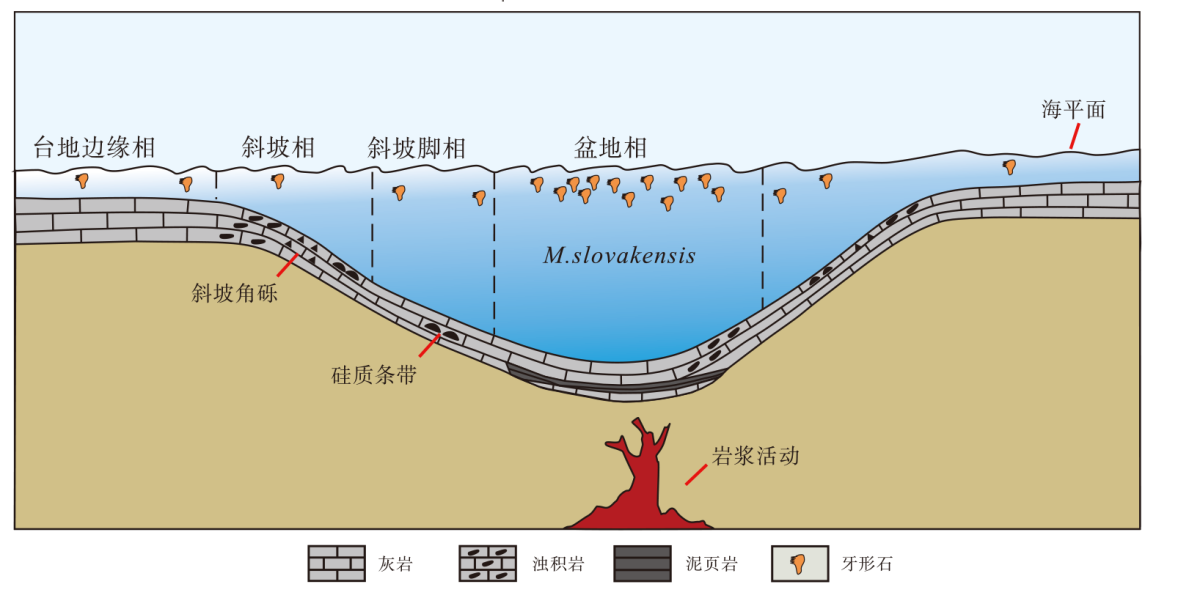 | 图 7 云南保山地区上三叠统大水塘组沉积模式Fig.7 Sedimentary model of the Upper Triassic Dashuitang Formation in Baoshan area, Yunnan Province |
由于牙形石Mockina slovakensis易于识别, 且在该时期的特提斯地区广泛分布, 因此被作为特提斯地区诺利期Alaunian晚期至Sevatian晚期的标志性属种。对于M. slovakensis的形态特征研究, 前人已经做过大量工作(e.g., Kozur, 1972; Budai and Ková cs, 1986; Ková cs and Nagy, 1989; Du et al., 2021; 曾卫平, 2022), 目前M. slovakensis的鉴定主要依靠2个最显著的形态特征: 齿片末端到齿脊呈现陡峭、突然的下降, 以及其龙脊末端从不分叉(图 5; 图 6)。其他形态特征的描述或分类还存在模糊性和争议性, 今后还需要进行更加详细的研究。
Mockina slovakensis具有重要的指示地层时代意义, 对地层划分亦具有重要的作用。该牙形石最初是由Kozur(1972)在上三叠统诺利阶Misikella hernsteini牙形石带中发现的, 因此, M. slovakensis被认为属于诺利期Sevatian晚期, 同时也可能延续至瑞替期(Moix et al., 2007; Kozur, 1972)。Budai和Ková cs(1986)报道了匈牙利Rezi Dolomite岩层中的M. slovakensis牙形石动物群, 认为M. slovakensis的时代应为诺利期Alaunian亚期。Ková cs和Nagy(1989)根据Budai和Ková cs(1986)的研究结果, 同样将M. slovakensis归为中诺利期, 但认为其也可能属于诺利期Sevatian早期, 其原因是M. slovakensis在地层中与菊石Rhabdoceras suessi一起出现(Moix et al., 2007)。Roghi等(1995)报道了意大利东北部Seazza Creek剖面的M. slovakensis, 并结合Budai和Ková cs(1986)、Ková cs和Nagy(1989)公布的数据, 将其归于诺利期Alaunian中晚期。Meç o(1999)报道了阿尔巴尼亚诺利期Sevatian晚期的M. slovakensis。Martini等(2000)报道了来自帝汶岛的M. slovakensis, 通过菊石证实其应属于诺利期Alaunian亚期(Martini et al., 2000; Moix et al., 2007)。Donofrio 等(2003)报道了在阿尔卑斯山Seefeld剖面台内盆地中的以M. slovakensis为主的牙形石动物群, 并将其时代归为诺利期Alaunian中期。Channell等(2003)报道了斯洛伐克Silická Brezová 的M. slovakensis, 并将其时代划为诺利期Alaunian晚期。Moix等(2007)提出当M. slovakensis与E. praeslovakensis一起出现时, 其代表的时代应为诺利期Alaunian晚期, 但当地层中只有M. slovakensis时, 其时代可能为诺利期Alaunian晚期至Sevatian晚期。Belvedere等(2008)报道了E. praeslovakensis和M. slovakensis, 并将其时代归为诺利期Alaunian晚期。Balini等(2010)在意大利西西里岛西部的Pizzo Mondello剖面的Sevatian地层中发现了“ Epigondolella” slovakensis, 随后Mazza 等(2012)也在同一地区的剖面中发现了M. slovakensis, 并认为其时代为诺利期Sevatian晚期。Rigo等(2012)在意大利拉各内格罗盆地Sasso di Castalda剖面发现了M. slovakensis, 将其时代归属于诺利期Alaunian 晚期至Sevatian 中期, 其后Rigo 等(2016)在拉戈内格罗盆地的Pignola Abriola剖面发现了M.slovakensis牙形石, 且将其时代归为诺利期Sevatian亚期。Yamashita等(2018)日本中部的犬山地区也发现了M. slovakensis牙形石, 同样将其时代归为诺利期Sevatian亚期。Du 等(2021)在Dolomia di Forni地区的Valmenone、Poschiedea Rovadia、Seazza和Forchiar Creek以及Rio Resartico等剖面均发现了M. slovakensis牙形石, 并认为其时代应为Alaunian晚期至Sevatian中期。
在云南保山地区, Zeng 等(2021)以及曾卫平(2022)在保山喜鹊林剖面发现了牙形石M. slovakensis, 但没有建立单独的M. slovakensis牙形石带。由于该剖面M. slovakensis出现在M. bidentata带和P. andrusovi带中, 因此根据M. bidentata带和P. andrusovi带可以推知喜鹊林剖面的M. slovakensis的时代归属应为诺利期Sevatian亚期。Jin 等(2022)报道了保山红岩剖面的M. slovakensis牙形石, 并建立了M. slovakensis牙形石带和M. bidentata牙形石带(图 2)。在M. slovakensis带中发现E. praeslovakensis和M. slovakensis共存, 但在M. bidentata牙形石带中并未发现M. slovakensis, 因此将M. slovakensis牙形石带的时代归为诺利期Aluanian晚期(Moix et al., 2007; Rigo et al., 2018; Jin et al., 2022)。Jin等(2022)在保山红岩剖面发现的M. slovakensis时代要早于Zeng 等(2021)以及曾卫平(2022)在保山喜鹊林剖面发现的M. slovakensis牙形石。但总的来看, 保山地区的M. slovakensis的时代分布范围在诺利期Alaunian晚期至Sevatian晚期之间, 与全球其他地区的研究结果相符。
综合前人所有关于M.slovakensis牙形石时代分布的研究结果, 可以得出M.slovakensis牙形石的时代分布范围大致为诺利期Alaunian晚期至Sevatian晚期, 当其与E.praeslovakensis同时出现时, 其指示的时代为诺利期Alaunian晚期。本次研究在红岩剖面下段的大水塘组中发现M.slovakensis和 E.praeslovakensis 共同出现, 且未发现牙形石M.bidentata, 因此认为研究层位的时代应为诺利期 Aluanian 晚期(图 2)。
在前人的研究中, M. slovakensis牙形石的生活环境被认为是多样的, 包括局限的盆地环境、浅水环境、较开阔或半开放远洋盆地环境(Du et al., 2021)。Trotter等(2015)分析了M. slovakensis的氧同位素组成, 认为该物种栖息在温暖的上层水体中。相对于开放的海洋环境(包括浅水和深水), 局限环境指的是与开放或广阔的海洋在连通性、海水的交换等方面均较弱的一种环境。最初, M. slovakensis经常被大量发现于上三叠统的台内盆地中, 如匈牙利Keszthely山脉的Rezi Dolomite组和Feketehegy组(Ková cs and Nagy, 1989; Budai and Ková cs, 1986)、奥地利南部阿尔卑斯山的Seefeld组(Donofrio et al. 2003)、安纳托利亚西北部的上三叠统(Kaya et al., 2001)以及意大利的Dolomia di Forni组(Roghi et al., 1995; Du et al., 2021)。近年来越来越多的报道表明, 较开阔或半开放远洋盆地环境也适合M. slovakensis的生存, 如在意大利拉戈内格罗盆地Sasso di Castalda剖面和Pignola Abriola剖面的Calcari con Selce组(Rigo et al., 2012, 2016)、阿尔巴尼亚(Meç o, 1999)、斯洛伐克(Channell et al., 2003)、意大利Mont Cocuzzo盆地(Mastandrea et al., 1997)、西西里岛Pizzo Mondello剖面的Scillato组(Mazza et al., 2012)以及日本犬山地区Kamiaso组地层中(Yamashita et al., 2018)都有记录和报道。此外, M. slovakensis也被发现生活在较浅的水环境中, 如以Dolomia Principale组为代表的碳酸盐台地(Belvedere et al., 2008), 不过在浅水环境中报道的次数较少。
总的来说, M. slovakensis分布的海洋环境较为多样, 从浅水到深水、从局限环境到开放环境均有分布, 但Du 等(2021)依据前人的研究结果总结出了一个很特别的现象: 当M. slovakensis在牙形石群体中占据主导地位(即M. slovakensis数量较多, 且为绝大多数)的时候, 其沉积环境均为与开放海水或海域连通性较差的局限环境, 尤其是局限的台内盆地(Roghi et al., 1995; Donofrio et al., 2003; Du et al., 2021); 在开放的浅水或深水环境中, 虽然M. slovakensis也能够生存, 但其并不会占据牙形石群体的统治地位, 而是和其他种属的牙形石平均分布, 有时甚至处于弱势地位(数量往往较少)。导致出现这种现象的原因目前仍然是一个谜, 但可能的原因为M. slovakensis的生存能力更强, 在生存条件有限的情况下(局限环境), 其在与其他同时期牙形石动物的竞争中处于优势地位。然而, 这却不能解释为何在开放环境(包括浅水和深水)中M. slovakensis没能占据统治地位, 有时候甚至处于弱势地位的问题。另一种更可能的原因是牙形石M. slovakensis的群体性很强, 更加适应局限的生存环境, 而在开放的环境中, 由于生存资源充足, M. slovakensis的群体性优势可能会被抵消, 但在局限环境中群体性的优势则能够使得M. slovakensis在生存竞争中胜出, 从而占据统治地位。在长期的演化过程中, 随着M. slovakensis不断地在局限环境的生存竞争中获胜, 其变得越来越适应局限的生存环境, 因此阻碍了M. slovakensis向不同地区的迁移。这也解释了为何M. slovakensis的报道几乎都集中在特提斯区域, 而在泛大洋只有零星的报道(Yamashita et al., 2018), 而在北美地区目前还没有关于M. slovakensis的报道。
正是由于M. slovakensis这种独特的生活习性(适应局限的生存环境), 且根据前人总结的经验性规律, 可以推测当M. slovakensis在牙形石群体中占据统治地位时, 其往往指示出一种较为局限的沉积环境。
保山地块晚三叠世经历了复杂多变的沉积过程: 在晚三叠世卡尼期, 保山地块发生了一次广泛的海侵, 海平面随之上升, 此时的保山形成一片广阔的海洋; 在诺利期海水持续侵进, 沉积范围不断扩大; 直至晚三叠世瑞替期, 印支运动到达鼎盛时期, 地壳持续上升, 海水随之全部退出并结束了海相沉积的历史(胡友恒, 1990; 包佳凤等, 2020)。晚三叠世保山地块海相沉积较为发育, 但对于该地区沉积环境的研究程度不足并且存在一定的争议。云南省地质矿产局(1990)和云南省地质局区域地质调查队(1980)都认为保山地块晚三叠世为滨海— 浅海环境①。张远志等(1996)认为保山地块在早— 中三叠世大多数为滨海— 浅海环境, 在晚三叠世除部分地区有中酸性— 酸性火山喷发外, 主要为浅海台地— 滨海潮坪— 近海河湖沉积环境。
对于大水塘组, 郝子文和饶荣标(1999)认为云南保山大水塘组的灰岩主要是浅海相的, 属于碳酸盐岩台地沉积。赵云江等(2012)认为保山地块大水塘组主体沉积了一套钙质浊积岩, 并推断其沉积环境为斜坡— 盆地相, 而包佳凤等(2012, 2020)报道了保山金鸡地区大水塘组的震积岩, 也提出其沉积环境为斜坡— 盆地环境, 并认为该震积岩是在强烈拉张背景下形成的斜坡— 盆地的钙质浊积岩的基础上改造形成的。在晚三叠世, 云南保山地块处于拉张裂陷背景, 火山活动和构造运动较为发育, 事实上保山地块是大型断陷盆地的一部分, 其上三叠统大水塘组沉积环境是在强烈拉张的背景下形成的裂谷, 在这个裂谷中发育有良好的古斜坡和盆地(马杏垣, 1982; 燕守勋等, 1996; 包佳凤等, 2012, 2020; 赵云江等, 2012)。彭成龙等(2014)在保山地块姚关地区上三叠统大水塘组发现钙屑浊积岩, 推断其发育于裂陷槽盆环境。王小端等(2019)在对云南保山大堡子地区上三叠统牙形石和沉积环境的研究中, 根据微相推断大水塘组顶部灰岩为具有一定坡度的陆棚环境沉积。吴强旺等(2020)采用微相分析的方法对保山红岩剖面大水塘组碳酸盐岩进行研究, 归纳细分了8种微相, 识别出斜坡相、斜坡脚相及深水陆棚相3种沉积相类型(图 2), 并指出在大水塘组沉积时期该地区是一个深水斜坡, 其形成于低能的相对深水环境。曾卫平(2022)在云南保山地区坡头剖面中报道了与台内局限区和开阔海相似的碳酸盐岩微相, 并揭示了坡头剖面大水塘组和南梳坝组为低能水流波动的台内局限缓坡环境。
综合前人的研究成果, 保山地区在诺利期Alaunian晚期可能是一个形成于拉张环境的沉积盆地, 红岩剖面所在的地区正好位于该沉积盆地的斜坡— 斜坡脚(吴强旺等, 2020)。前人研究发现保山地块在晚三叠世卡尼期时有一套基性+酸性+基性火山岩喷发, 即牛喝塘组, 其具有大陆裂谷的演化特征, 保山地块西缘形成于中生代裂陷张裂较强的环境(王义昭等, 2000), 但牛喝塘组在保山红岩剖面附近的区域并不发育。廖世勇等(2013)对云南保山地块东缘存在的岩浆岩进行研究, 发现其时代约为216.8± 2.2 Ma, 这一年龄与M. slovakensis指示的时代Aluanian晚期很接近。这表明从卡尼期到诺利期, 保山地区周围一直存在着活跃的岩浆或构造活动, 这也解释了保山地区大水塘组震积岩发育以及红岩剖面下端大水塘组浊积岩以及滑塌构造发育的原因。
总的来说, 前人多数的观点认为中诺利晚期保山地块大水塘组沉积应该发育于一个构造运动形成的沉积盆地(马杏垣等, 1982; 燕守勋等, 1996; 包佳凤等, 2012, 2020; 赵云江等, 2012)。结合本次研究在红岩剖面发现了大量占据统治地位的中诺利晚期牙形石M. slovakensis(图 4), 可以推测该盆地的沉积环境应该较为局限, 与外界的沟通较少, 使得M. slovakensis得以迅速在该环境中占据统治地位, 从而挤占了其他牙形石动物的生存空间。在晚诺利初期(M. bidentata牙形石带), 研究区的构造活动一直很活跃(发育浊积岩和滑塌构造), 且伴随着海水的不断加深(图 2), 该局限盆地和外界产生了更多的沟通, 因此环境中生存的牙形石种类也变得更加多样, M. slovakensis失去统治地位并消失。最后, 随着诺利晚期大量陆源碎屑的输入, 牙形石在保山地区的踪迹也彻底消失。
由牙形石M. slovakensis指示出的保山地区的这一系列沉积环境演化, 很可能与特提斯洋古板块的逐次拼合、运动存在联系, 故研究结果也有利于了解晚三叠世特提斯东部的沉积环境演化。
1)在云南保山红岩剖面下部大水塘组发现了大量的牙形石(样品HYB-20至HYB-26), 以Mockina属为主, 含有少量Epigondolella属, 其中M. slovakensis在地层中占据了统治地位。
2)依据红岩剖面下部大水塘组中M. slovakensis和E. praeslovakensis共同出现, 且未发现M. bidentata, 推断该段地层的时代为诺利期Alaunian晚期。
3)M. slovakensis不仅能够适应从浅水到深海的环境, 也能够适应从局限环境到开放环境, 但当地层中M. slovakensis占据统治地位时, 其往往指示一种较为局限的沉积环境。综合M. slovakensis牙形石的生活习性以及前人对保山地块大水塘组沉积环境的研究成果, 认为保山地区在诺利期Aluanian晚期可能是一个构造运动形成的较为局限的沉积盆地。
致谢 感谢两位审稿专家提出的宝贵的意见和建议, 使文章质量得到显著提升!
(责任编辑 张西娟; 英文审校 刘贺娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|