




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
豹皮(斑)状碳酸盐岩分类与成因初探: 以鄂尔多斯盆地马家沟组四段为例*
[魏柳斌1, 2  , 王宗延
, 王宗延3 , 李漪3 , 庞志超3 , 王前平1, 2 , 包洪平1, 2 , 杨琦琦3 , 王振3 , 苏中堂4 , 张立军3 ]
, 王宗延]
|
|

第一作者简介 魏柳斌,男,1987年生,工程师,主要从事碳酸盐岩油气地质研究工作。 E-mail: weilb_cq@petroChina.com.cn。
豹皮(斑)灰岩是华北寒武系—奥陶系广泛发育的一种特殊形貌的海相碳酸盐岩,因具有显著的识别特征和成因机制而备受关注。鄂尔多斯盆地中奥陶统马家沟组四段发育丰富的豹皮(斑)碳酸盐岩,目前已在其中识别出遗迹化石 7属 8种,分别是Thalssinoides network,Thalassinoides boxwork,Planolites isp.,Palaeophycus tubularis,Phycodes isp.,Chondrites isp.,Skolithos isp.,?Zoophycos isp.,其以Thalassinoides的 2种形态类型(水平迷宫形 network和三维箱状结构 boxwork)占主导,属于固底底质条件的 Glossifungites遗迹相。通过野外露头、岩心的宏观和微观观察,系统调研前人关于生物扰动成因碳酸盐岩的类型,依据生物扰动、生物潜穴形态特征和方解石 /白云石相对含量,笔者提出了生物扰动成因豹皮(斑)状碳酸盐岩的分类方案,将其主要分为豹皮灰岩和豹斑云岩 2个大类,分别对应Thalassinoides network(水平二维迷宫)和Thalassinoides boxwork(三维箱状结构) 2种形态类型。鄂尔多斯盆地马家沟组四段的豹皮(斑)状碳酸盐岩主要受生物扰动和高频海平面升降下的早期成岩作用耦合控制,表现为在固底底质条件下,生物潜穴为富 Mg2+卤水的白云石化流体提供了良好的运移通道,使得潜穴内部优先发生白云石化作用,而不同类型的生物潜穴及准同生和埋藏白云石化作用,最终形成了豹皮灰岩和豹斑云岩这 2种类型的生物扰动成因豹皮(斑)状碳酸盐岩。
, WANG Zongyan
About the first author WEI Liubin,born in 1987,is an engineer of PetroChina Changqing Oilfield Company. He is mainly engaged in research on carbonate reservoir. E-mail: weilb_cq@petroChina.com.cn.
Leopard skin (spot) limestone, a type of marine carbonate rock with unique morphology, is widely distributed in the Cambrian-Ordovician strata of North China. It has attracted significant attention due to its distinct characteristics and formation mechanism. Abundant leopard-skin (spot) carbonate rocks developed in Member 4 of the Middle Ordovician Majiagou Formation in the Ordos Basin. Eight species of 7 genera have been identified, including Thalssinoides network, Thalassinoides boxwork, Planolites isp., Palaeophycus tubularis, Phycodes isp., Chondrites isp., Skolithos isp.,? Zoophycos isp. Two morphologic types of Thalassinoides (horizontal enigmatic network and three-dimensional boxwork) are predominant, belonging to the Glossifungites ichnofacies under firm ground condition. Based on detailed macroscopic and microscopic characteristics of outcrops and drilling cores, this paper systematically summarizes the types of carbonate rocks caused by bioturbation. By examining the morphological characteristics of bioturbation and burrow, as well as the calcite/dolomite content, this paper proposes a classification for leopard-skin (spot) carbonate rocks formed by bioturbation. These rocks are categorized into leopard-skin limestone and leopard-spot dolomite, corresponding to the Thalassinoides network (two-dimensional horizontal mystery palace) and the Thalassinoides boxwork (three-dimensional box-like structure), respectively. The leopard-skin (spot) carbonate rocks in Member 4 of the Majiagou Formation in the Ordos Basin are primarily controlled by the interaction of bioturbation and early diagenesis during high-frequency sea level fluctuations. The bioturbated burrows in the firm substrates creat pathways for Mg2+ rich brines, promoting dolomitization within the burrows. Two distinct types of leopard-skin (spot) carbonate rocks, induced by bioturbation, were generated under varying bioturbation types, parasyngenesis, and buried dolomitization.
豹皮灰岩, 是一种具有特殊形貌的碳酸盐岩, 在中国华北寒武纪— 奥陶纪海相碳酸盐岩地层、四川盆地栖霞组和塔里木盆地鹰山组中广泛分布(翟淳, 1961; 张俊明等, 1981; 陈战杰和张镔, 1991; 武永强和吴卓丹, 1995; 李定龙等, 1999; 陈云峰等, 2007; 龙刚等, 2013; 王起琮等, 2016; Zhang et al., 2017; 吴灿灿等, 2018; 刘梦瑶等, 2020; 许杰等, 2022; 周进高等, 2022)。“ 豹皮灰岩” 最早由赵亚曾在1929年描述结核灰岩中提出, 1945年庄司诚一等称之为“ 云状灰岩” , 1956年陈世波等命名为“ 黄花灰岩” , 而中国地质大学一直以“ 豹皮灰岩” 来描述华北寒武纪— 奥陶纪地层中的斑状灰岩(翟淳, 1961; 陈战杰和张镔, 1991)。近30多年以来, 随着海相碳酸盐岩油气资源勘探的深入, “ 豹斑状云质灰岩” 、“ 斑状云岩” 、“ 豹斑灰岩” 、“ 云斑灰岩” 、“ 斑点状灰岩” 、“ 蠕虫状灰岩” 等术语被先后提出(贾振远和马淑媛, 1984; 龙刚等, 2013; 董小波和牛永斌, 2015; 张天付等, 2018; 刘梦瑶等, 2020; 许杰等, 2022; 周进高等, 2022), 也有学者用 “ 豹斑构造” 、“ 花斑状构造” 来描述此类灰岩(冯增昭等, 1989), 造成同物异名现象, 不具有普识性和规律性, 因此很有必要将术语进行统一。
豹皮灰岩的形成机制主要分为无机和有机无机混合2个类型, 前者认为其形成原因是以岩溶和机械重力溶蚀为主(翟淳, 1961; 李定龙等, 1999), 后者则认为生物扰动和白云岩化流体的盐度是控制豹皮灰岩形成的主要因素(Gingras et al., 2004, 2012; Jin et al., 2012; 刘梦瑶等, 2020; Niu et al., 2022; 许杰等, 2022)。
鄂尔多斯盆地中东部奥陶系马家沟组海相碳酸盐岩一直是中国石油长庆油田分公司的主力勘探层位之一, 近年来, 该公司加大了盐下深层碳酸盐岩(马四段— 马一段)的勘探强度, 并在2021年MT1井的马四段获得了高产工业气流的重大突破(付金华等, 2022)。前人对于鄂尔多斯盆地奥陶系马家沟组四段的研究主要集中于岩相古地理、白云岩储集层特征及控制因素等方面(如吴东旭等, 2021; 许杰等, 2022; 周进高等, 2022; 于洲等, 2023a, 2023b; 牟春国等, 2023), 对于马四段豹皮灰岩和斑状白云岩的成因及储集层物性等认识还存在很大分歧, 尚不统一。近年来, 笔者在鄂尔多斯盆地奥陶系马家沟组四段中识别出丰富的豹皮(斑)状碳酸盐岩, 通过详细的野外宏观特征观察和镜下微观特征分析, 基于宏观形貌特征和方解石/白云石相对含量, 提出了豹皮(斑)状碳酸盐岩的分类方案, 并将之对应于相应的遗迹组构类型, 以期为今后的生物扰动成因碳酸盐岩研究及相关储集层预测提供新的参考和借鉴。
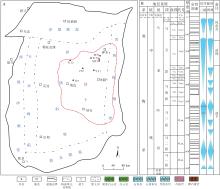
鄂尔多斯地块位于华北地台西部, 可划分为晋西挠褶带和伊陕斜坡在内的6个构造单元(图 1-A), 其中伊陕斜坡为鄂尔多斯盆地最主要的油气富集区域(付金华等, 2022)。华北地台奥陶系为一套浅水碳酸盐岩台地沉积, 发育多期次海侵与海退旋回。鄂尔多斯盆地中东部下奥陶统由冶里组和亮甲山组构成, 冶里组以砂砾屑的竹叶状灰岩为主, 亮甲山组以富含燧石条带及团块的中厚层白云岩为主; 中奥陶统由马家沟组构成, 马家沟组发育碳酸盐岩— 膏盐岩共生体系沉积, 纵向上具有明显的旋回性, 共分为6个岩性段, 其中马一段、马三段和马五段为海退沉积旋回, 主要发育蒸发台地和局限台地背景下的膏盐岩及白云岩, 马二段、马四段和马六段为海侵沉积旋回, 主要发育开阔台地背景下的石灰岩。
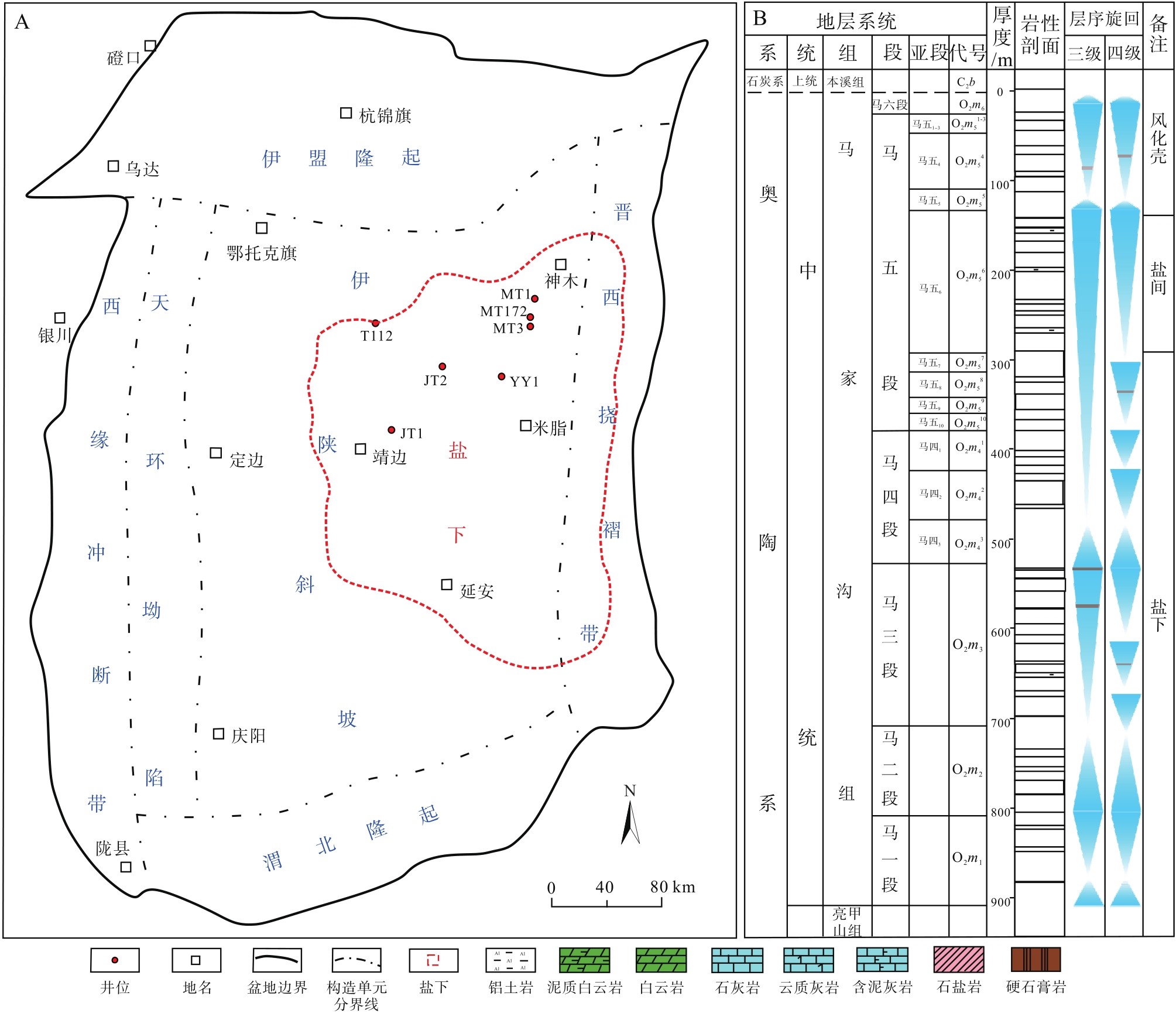 | 图 1 鄂尔多斯盆地构造纲要图(A)和中奥陶统综合柱状图(B)(据付金华等, 2022; 周进高等, 2022; 有修改)Fig.1 Simplified structural map(A)and stragiraphic column of the Middle Odovician(B) in Ordos Basin(modified from Fu et al., 2022; Zhou et al., 2022) |
鄂尔多斯盆地中东部盐下马四段厚度相对稳定, 约150 m, 进一步细分为3个亚段, 由上而下依次为马四1、马四2和马四3亚段。马四3亚段为海侵期沉积, 主要岩性为泥晶石灰岩、云质石灰岩和灰质白云岩, 厚50~60 m; 马四2亚段为海退早期沉积, 主要岩性为泥晶石灰岩、云质石灰岩和颗粒白云岩, 厚50~60 m; 马四1亚段为海退中期沉积, 主要岩性为白云岩和硬石膏岩互层, 厚40 m左右(图1-B)。笔者在马家沟组四段中识别出了丰富的遗迹化石, 主要包括Thalassinoides, Planolites, Palaeophycus, Skolithos, 它们从属于固底底质条件的Glossifungites遗迹相, 主要分布于潮坪或潟湖沉积环境。
笔者实测了2条野外剖面(山西柳林剖面和偏关闫贵家嘴剖面), 并对7口全取心井进行实际观察和描述, 总计岩心长763.2 m; 共拍摄岩心和野外相片3000余张, 鉴定岩石薄片1000余个。野外实测评估生物扰动强度时使用国际通用的生物扰动指数(Bioturbation Index=BI), 其由小到大、由无到有、由弱到强, 不同的比例对应不同等级的生物扰动强度(BI=0~6)(Taylor and Goldring, 1993)。
岩石薄片和光面形态的宏观和微观特征分别利用偏光显微镜(德国卡尔蔡司Axio Imager M2 Pol, 配备506 color CCD摄像头, 河南省生物遗迹与成矿过程国际联合实验室, 河南理工大学, 焦作)观察。扫描电镜样品制备时, 先将切割好的岩石标本用蒸馏水清洗烘干后用导电胶固定在样品台上(样品台直径约为1 cm), 放入金属离子溅射仪中进行喷金处理, 喷金时间60~80 s; 之后使用场发射扫描电镜对其进行相对应的超微形态特征观察。场发射环境扫描电镜(ESEM)型号为FEI Quanta 250 FEG, 布鲁克能谱仪(EDS)型号为Bruker Quantax 200 XFlash 6|30), 实验地点为河南省生物遗迹与成矿过程国际联合实验室(河南理工大学, 焦作)。
中奥陶统马家沟组四段主要发育遗迹化石7属8种, 分别是Thalssinoides network, Thalassinoides boxwork, Planolites isp., Palaeophycus tubularis, Phycodes isp., Chondrites isp., Skolithos isp., ?Zoophycos isp., 从属于固结底质底质的Glossifungites(舌菌迹)遗迹相。
Palaeophycus tubularis呈不分枝的柱形水平潜穴, 潜穴直径约1.0 cm, 表面光滑, 具有清楚的衬壁, 主要分布于马四3亚段的豹皮灰岩中(图 2-A至2-D)。Planolites isp. 主要表现为平行层面的柱形潜穴, 不分枝, 表面较粗糙, 无衬壁, 主要分布于马四段(图 2-E)。Phycodes isp. 主要沿层面分布, 呈水平束状潜穴, 由1条主潜穴向外延伸呈手掌状分枝, 各分枝互不相交, 整体呈簇状扇形, 主潜穴宽约3.2 mm, 分枝潜穴直径1.0~1.2 mm, 主要分布于马四3亚段(图 2-F)。Chondrites isp. 表现为树枝形细小分枝潜穴系统, 主枝短粗, 分枝较短但不密集, 可见2次分枝, 潜穴直径1.5~2.0 mm, 主要见于马四3亚段和马四2亚段(图 2-G, 2-H)。Thalassinoides network主要为三维空间分支的潜穴系统, 水平面上常呈Y形、T形分枝, 分枝处膨大略粗, 纵切面上潜穴之间互不连通, 潜穴直径4.5~10.8 mm, 主要分布于马四3亚段(图 3-D至3-H)。Thalassinoides boxwork主要为相互连通、交切连成网格状的复杂潜穴系统, 常见Y形、T形或V形分枝, 分枝处膨大略粗, 多级潜穴互相覆盖, 潜穴直径9.2~12.8 mm, 主要分布于马四1亚段和马四2亚段(图 3-I至3-M)。?Zoophycos isp.主要表现为螺旋形蹼层, 可见边缘管, 在纵切面上可以看到交互出现的深色和浅色小蹼层, 边缘管直径约5.2 mm, 主要分布于马四1亚段(图 2-I)。Skolithos isp. 主要表现为与层面垂直呈高角度倾斜的针形潜穴, 不分枝, 颜色较围岩深, 内部为被动充填, 多期次的殖居窗口可反映底质的变化, 主要分布于马四1亚段(图 4-E)。
 | 图 2 鄂尔多斯盆地中奥陶统马家沟组四段典型遗迹化石 A, B— 山西柳林三川河剖面, 马四3亚段, 含云石灰岩表面发育的Palaeophycus tubularis; C, D— M172井, 泥晶灰岩中见到的Palaeophycus的横切面, 衬壁为厚层的白云石; E— JT2井, 3399.32 m, 马四2亚段, 发育Planolites; F— 山西柳林三川河剖面, 马四3亚段, 云质石灰岩表面发育的掌状Phycodes isp.; G, H— 山西柳林三川河剖面, 马四3亚段, 云质石灰岩表面发育的树枝形Chondrites isp.; I— JT1井, 3762.21 m, 马四1亚段, 发育疑似 ?Zoophycos isp., 可见边缘管及浅色和深色的小蹼纹组成的蹼层Fig.2 Trace fossils from the Member 4 of Middle Ordovician Majiagou Formation in Ordos Basin |
 | 图 3 鄂尔多斯盆地中奥陶统马家沟组四段中发育的Thalassinoides A— Thalassinoides的主要形态结构示意图; B— Thalassinoides network(二维迷宫结构)示意图; C— Thalassinoides boxwork(三维箱状结构)示意图; D, E— 山西柳林三川河剖面, 马四3亚段, 豹皮灰岩纵切面和上层面中发育的Thalassinoides network; F— M172井, 2854.41 m, 马四2亚段, Thalassinoides network, 潜穴中充填为白云石, 基质为泥晶, 缺少相互交切关系; G— 山西柳林三川河剖面, 马四3亚段, 斑状白云质石灰岩中Thalassinoides network的显微特征, 潜穴中充填物的为白云岩, 基质为泥晶; H— M172井, 2859.16 m, 马四3亚段, 斑状白云质石灰岩中Thalassinoides network的显微特征, 潜穴中充填为晶粒白云石, 发育晶间孔, 基质为泥晶; I, J— 山西柳林三川河剖面, 马四3亚段, 豹斑云岩中Thalassinoides boxwork的宏观特征, 潜穴相互切, 呈厚层斑状, 见Y形分枝和膨大的转折端; K— M172井, 2875.01 m, 马四2亚段, 斑状灰质白云岩中的Thalassinoides boxwork显微特征, 潜穴中为镶嵌状白云石, 基质中白云石呈零散状, 由潜穴向基质扩散; L— M172井, 2877.16 m, 马四2亚段, 斑状白云岩中的Thalassinoides boxwork, 主要显示了多期次的生物扰动, 最后1次的生物扰动中潜穴充填的白云石颗粒较大, 早期的生物扰动已成为基质, 白云石颗粒较小; M— M172井, 2835.84 m, 马四2亚段, Thalassinoides boxwork的宏观特征, 潜穴多期次相互交切, 形成三维箱状结构Fig.3 Abundant Thalassinoides from the Member 4 of Middle Ordovician Majiagou Formation in Ordos Basin |
 | 图 4 鄂尔多斯盆地中奥陶统马家沟组四段遗迹化石反映的沉积底质条件特征 A— 遗迹化石形态特征反映的不同底质条件变化(据Ekdale et al., 1984, 有修改); B— JT1井, 3835.10 m, 马四3亚段, 软底向固底的变化; C— YY1井, 3131.98 m, 马四2亚段, 复合遗迹组构包括2个明显的生物扰动阶段, 侵蚀前发育的Palaeophycus(Pa)圆柱形潜穴为软底, 而后形成的Thalassinoides(Th)发育在固底并向上进行挖掘; D— YY1井, 3153.51 m, 马四3亚段, 识别出的由固底至软底至固硬底的变化; E— JT2井, 3317.63 m, 马四1亚段固底中不同遗迹化石反映的殖居窗口的长短变化特征, 遗迹化石深度越大, 代表殖居窗口时间越长, Sk全写为Skolithos isp.Fig.4 Characteristics of substrate reflected by trace fossils from the Member 4 of Middle Ordovician Majiagou Formation in Ordos Basin |
遗迹化石的形态特征可以很好地指示沉积物的底质条件类型(图 4)。固底(Firmground)是底质脱水压实形成的产物, 主要特点为上层被侵蚀到较深的层次后暴露在海底表面, 遗迹化石的潜穴边界明显、突出、清晰(Ekdale et al., 1984; Buatois and Má ngano, 2011)。硬底(Hardground)是由海底液化产生的, 其长期暴露在海底动荡的水域, 内部水循环导致孔隙已被胶结物充填, 生物扰动作用停止, 取而代之的是生物侵蚀作用、生物钻孔发育(Ekdale et al., 1984; Buatois and Má ngano, 2011)。
马四段的遗迹化石很好地反映了底质的变化。如JT1井马四3亚段3835.10 m处(图 4-B), 下部生物潜穴边界较模糊, 体现出部分压实作用, 反映了软底的底质条件; 而上部生物潜穴边界突出明显, 基本未被压实, 指示固底的底质条件。JD2井马四1亚段固底中不同遗迹化石反映的殖居窗口的变化特征(图 4-D), 表明遗迹化石深度越大, 造迹生物殖居窗口的时间越长。
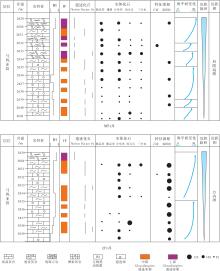
马四段的遗迹化石总体上表现为固底— 半固结底质的Glossifungites遗迹相。根据遗迹组构类型和遗迹化石组合特征, 将Glossifungites遗迹相进一步划分为上部、中部和下部Glossifungites遗迹亚相, 分别指示不同的水动力条件和沉积环境(图 5)。上部Glossifungities遗迹亚相主要以垂直的Skolithos和三维箱状结构的Thalassinoides boxwork为主, 多为居住迹和逃逸迹, 生物扰动指数4~6级, 生物扰动作用较强, 指示潮间带上部和潮上带的沉积环境; 中部Glossifungites遗迹亚相以发育水平的二维迷宫形的Thalassiniodes network和水平具衬壁的Palaeophycus为主, 生物扰动指数2~3级, 指示潮间带下部的沉积环境; 下部Glossifungites遗迹亚相主要以弱生物扰动的Thlassinoides network和部分Chondrites、Phycodes等为主, 生物扰动指数1~3级, 指示潮下带的沉积环境。
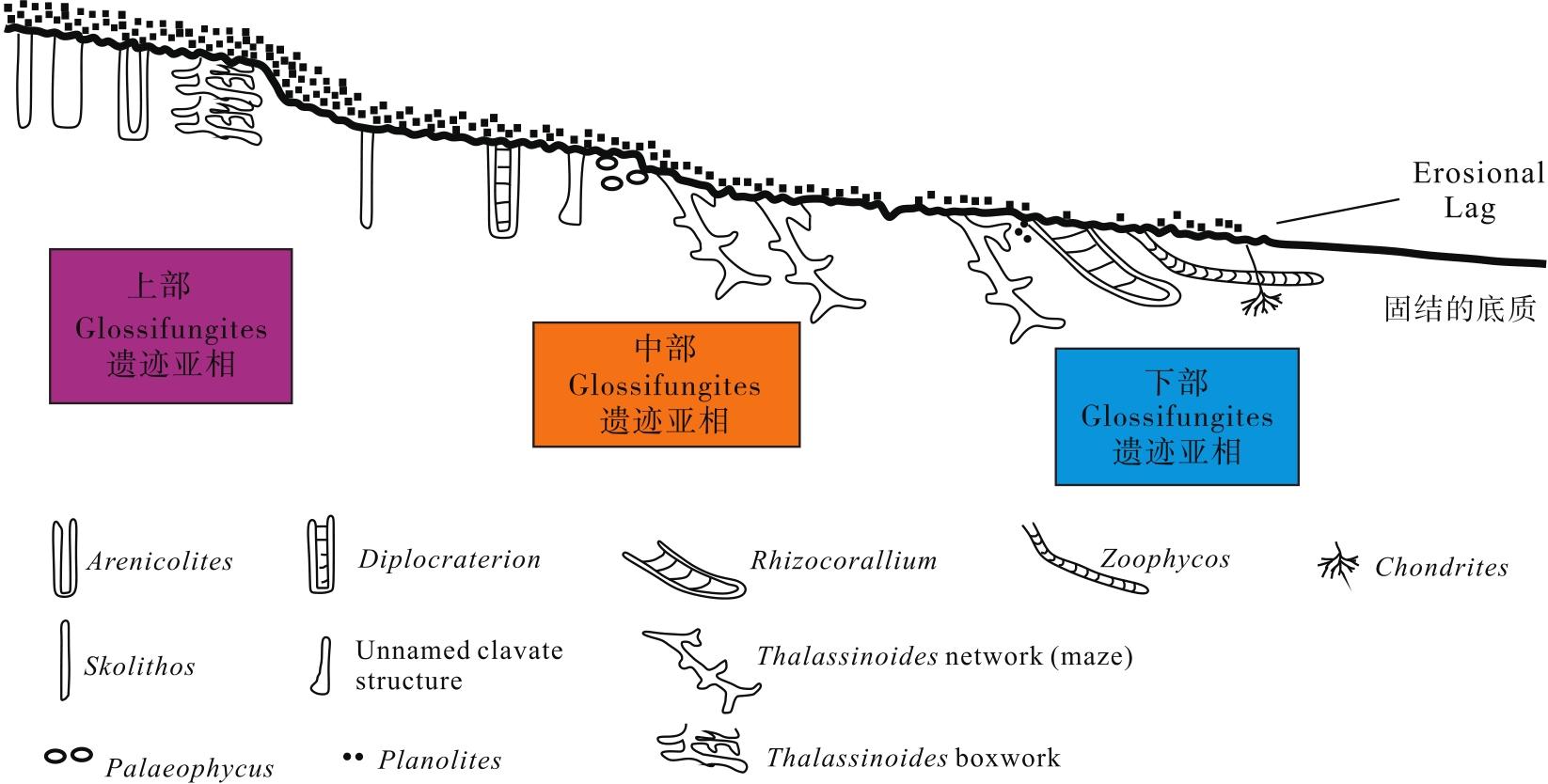 | 图 5 鄂尔多斯盆地中奥陶统马家沟组四段Glossifungites遗迹相示意图Fig.5 Sketch map of Glossifungites Ichnofacies in the Member 4 of Middle Ordovician Majiagou Formation in Ordos Basin |
笔者主要依据生物扰动强度、生物潜穴形态特征和方解石/白云石相对含量, 将豹皮(斑)状碳酸盐岩划分为豹皮灰岩和豹斑云岩2个类型, 分别对应Thalassinoides network(水平迷宫形)和Thalassinoides boxwork(三维箱状结构)2种形态类型。
4.1.1 Thalassinoides network与豹皮灰岩
鄂尔多斯盆地马家沟组马四3亚段中下部和马四2亚段中部条带状云质石灰岩(豹皮灰岩)非常发育, 主要富集Thalassinoides network。该生物潜穴呈水平方向延伸的网格状, 纵向上不连通, 风化面上潜穴边界相对围岩较为清楚, 局部较模糊, 保存为软底到固底。潜穴内部颜色较围岩部分略浅, 其中潜穴内部颜色多为灰白色和灰黄色, 围岩多为深灰色。水平层面上呈T形或Y形分枝, 分枝处略膨大, 垂直层面方向的分枝潜穴较短, 不与相邻的潜穴相互连通, 生物扰动指数为1~3级(图 3-D, 3-E)。
偏光显微镜下, 生物潜穴内部充填粗— 粉晶、半自形— 自形的白云石, 点— 面接触, 生物潜穴边界清晰(图 3-G, 3-H)。生物潜穴内部的白云石晶间孔非常发育, 在部分生物潜穴的边界还可以见腕足类或三叶虫的生物碎屑。围岩主要为泥晶方解石, 偶见孤立散落的、自形程度高的白云石。整体上, 豹皮灰岩中白云石含量小于50%, 白云石主要保存在生物潜穴中, 基质以泥晶方解石为主体。
4.1.2 Thalassinoides boxwork与豹斑云岩
鄂尔多斯盆地马家沟组马四3亚段上部、马四2亚段上部和马四1亚段斑状灰质白云岩和斑状白云岩(豹斑灰岩)中非常发育相互连通的三维箱状结构的Thalassinoides boxwork。生物潜穴在垂向上相互连通, 构成蜂窝型潜穴系统, 反映了多期次的生物扰动, 生物扰动指数为4~6级(图 3-I, 3-J, 3-M)。在水面层面上可以看到多期次相互交切的T形和Y形分枝, 分枝处膨大。斑状灰质白云岩中生物潜穴颜色较围岩略浅, 多为土黄色和灰白色, 斑状白云岩中生物潜穴多为土黄色。
在偏光显微镜下, 生物潜穴内部充填物以粗— 粉晶、半自形和他形白云石为主, 生物潜穴边界主要为扩散型, 零散的白云石由潜穴内部向围岩扩散(图 3-K, 3-L)。在生物扰动指数5级以上的斑状白云岩中, 基质见残余阴影结构。晶间孔在斑状灰质白云岩中发育(图 3-K)。整体上, 豹斑云岩中白云石含量大于50%, 白云石主要发育在生物潜穴中, 基质为泥晶方解石或粉晶白云石, 白云石充填的生物潜穴占主导。
4.1.3 分类
按着传统的岩石学分类与命名, Thalassinoides network赋存的豹皮石灰岩, 其主体是泥晶石灰岩或泥晶生物碎屑石灰岩; 依据生物潜穴发育的程度和白云石/方解石相对含量, 可以进一步划分为条带状含云质石灰岩或白云质石灰岩。Thalassinoides boxwork赋存的豹斑云岩, 其主体是斑状白云质灰岩和斑状白云岩, 反映的是4~6级强生物扰动作用形成的三维箱状结构(豹斑状结构)。如果不结合三维结构的宏观和微观形态特征, 很有可能将上述生物扰动成因的碳酸盐岩, 直接根据方解石和白云石含量进行命名, 从而忽略生物扰动作用对于此类岩石形成过程的重要影响, 因此从岩石成因的角度, 非常有必要对此类岩石进行重新厘定。依据鄂尔多斯盆地中奥陶统马家沟组四段中大量的岩心、岩石薄片和野外露头的豹皮灰岩和豹斑云岩的宏观和微观特征, 笔者提出了基于生物扰动成因的豹皮(斑)状碳酸盐岩分类(表 1)。
| 表 1 生物扰动成因的豹皮(斑)状碳酸盐岩分类 Table1 Classification of bioturbated lepord skin(spot)carbonate rocks |
这一分类体系, 主要依据生物扰动强度、生物潜穴形态特征和方解石/白云石相对含量划分为2个类型, 即豹皮灰岩和豹斑云岩。豹皮石灰岩(豹皮灰岩)主要发育水平展布的二维迷宫型Thalassinoides network, 生物潜穴内部充填物为白云石, 基质为泥晶方解石, 基质占主导。豹斑白云岩(豹斑云岩)按照生物扰动强度、白云石/方解石相对含量进一步划分为2个小类, 其中一类是完全生物扰动(BI=6)后形成的斑状白云岩, 基质为早期由于生物扰动残留的白云石, 潜穴中充填物为白云石, 总的白云石含量大于90%; 另一类是中— 强生物扰动作用(BI=4~5)形成的斑状灰质白云岩, 基质为颗粒— 泥晶方解石, 潜穴充填物为白云石, 潜穴占主导, 白云石含量50%~90%。
鄂尔多斯盆地奥陶系马四段豹皮灰岩主要发育于向上变浅序列的中下部, 豹斑云岩主要发育于向上变浅序列的上部(图 6)。马四段有2种类型的向上变浅序列(图7)。 第1种主要分布于马四3亚段, 以台内滩、生屑滩和潟湖组合向上变浅为特征。底部为泥晶灰岩, 发育水平泥纹层; 向上生屑和团粒增多, 云质含量明显增加, 发育水平展布的Thalassinoides network; 顶部发育三维箱状结构的Thalassinoides boxwork, 白云化程度进一步增强, 是灰云质潟湖的产物, 孔隙度和渗透率好, 为优质的储集层。第2种类型以潟湖和台坪向上变浅序列为特征。底部以泥晶灰岩为主; 中部云质含量增多, 发育有水平迷宫形Thalassinoides network, 水体盐度变化较大, 进入灰云质或云灰质潟湖沉积阶段; 顶部发育三维箱状结构的Thalassinoides boxwork, 伴随强烈的蒸发作用, 白云石化作用进一步增强, 转为云坪沉积阶段。 该序列发育于马四2和马四1亚段, 其中马四1亚段受到后期岩溶作用改造, 发育以晶间孔为主的颗粒白云岩, 是马四段最优质的储集层(牟春国等, 2023)。
 | 图 6 鄂尔多斯盆地中奥陶统马家沟组四段典型沉积旋回及遗迹相特征Fig.6 Typical sedimentary cycles and ichnofacies in the Member 4 of Middle Ordovician Majiagou Formation in Ordos Basin |
目前, 部分学者认为豹皮灰岩和豹斑云岩的形成与生物潜穴密切相关(刘梦瑶等, 2020; Niu et al., 2022; 许杰等, 2022)。鄂尔多斯盆地马四段豹皮灰岩和豹斑云岩中(图 7), 豹斑形白云石斑块主要沿潜穴分布, 并没有分布于基质之中, 且白云石斑块很好地保存了生物潜穴的原始形态特征, 从侧面说明生物潜穴相对于基质优先发生白云岩化作用。因此, 生物扰动作用是控制豹皮灰岩和豹斑云岩形成的主要因素。奥陶纪Thalassinoides可能是三叶虫类、甲壳类或蠕虫类所形成的居住迹和觅食迹(Myrow, 1995; Cherns et al., 2006; Jin et al., 2012)。生物潜穴的形态主要取决于水体能量、含氧量和营养物质等, 一般情况下, 潮下带水体能量较弱、营养物质丰富, 形成的生物潜穴大多为水平展布的觅食迹, 而在潮间带和潮上带, 形成的大多为垂直或倾斜方向的居住迹和逃逸迹(Ekdale et al., 1984; Bromley, 1996; Buatois and Má ngano, 2011)。鄂尔多斯盆地马四段的Thalassinoides为开放式的居住潜穴, 属于固底底质条件的Glossifungites遗迹相。固结的底底质条件为生物潜穴起到了支撑和保护作用, 一般情况下, 生物潜穴内部会形成还原性微环境(Baniak et al., 2013), 有利于白云石化作用。此外, 固底底质条件下Thalassinoides的生物潜穴为白云石化流体提供了快速、便捷的运移通道, 潜穴内部充填物在高镁卤水的回流渗透作用下优先发生白云石化作用, 进而形成 “ 砂糖状” 粉晶白云石(冯增昭等, 1989), 而后沿着潜穴边界向围岩渗透, 形成了扩散型边界(图 3-K, 3-L)。
 | 图 7 鄂尔多斯盆地中奥陶统马家沟组四段豹皮灰岩与豹斑云岩的微观特征 A— JT2井, 3374.58 m, 马四2亚段, 豹斑云岩, 白云石因镶嵌而呈浑圆形; B— JT2井, 3398.74 m, 马四2亚段, 豹斑云岩, 白云石的晶间孔非常发育, 靠近晶间孔一侧白云石生长迅速; C— JT1井, 3793.19 m, 马四2亚段, 豹斑云岩, 白云石雾心结构; D— M172井, 2875.01 m, 马四2亚段, 晶间孔与白云石的镶嵌结构; E, F— 山西柳林三川河剖面, 马四3亚段, 豹皮灰岩, 见白云石的微孔; G— MT1井, 2637.68 m, 马四2亚段, 豹皮灰岩, 潜穴中见腕足碎片, 白云石见雾心结构, 基质为泥晶; H— JT2井, 3403.08 m, 马四3亚段, 豹皮灰岩, 发育少许晶间孔; I— JT1井, 3830.38 m, 马四3亚段, 豹皮灰岩, 白云石因镶嵌结构而呈现浑圆形Fig.7 Microcharacteristics of leopard skin limestone and leopard spot dolostone in the Member 4 of Middle Ordovician Majiagou Formation in Ordos Basin |
鄂尔多斯盆地关家崖剖面马家沟组四段豹皮状碳酸盐岩的碳氧同位素分析表明, 生物潜穴中的δ 13C和 δ 18O 较同期的围岩略负偏, 其中马四段顶部的负偏程度较强, 向下负偏程度减弱, 说明生物潜穴中白云石的形成受到了大气淡水与海水的混合影响(许杰等, 2022)。颗粒白云岩和生物潜穴内部的细— 粉晶白云岩的U-Pb定年数据分别为342± 34 Ma和458± 28 Ma, 说明生物潜穴内部的白云石形成时间早, 为准同生白云化作用所形成, 而晶粒化的颗粒白云岩则经历了埋藏白云石化作用(于洲等, 2023b)。此外, 有研究指出, 与正常海水成因的泥晶灰岩相比, 马四段豹皮灰岩的云斑中Sr含量相对较低, 相对富集Fe、Mn; 与蒸发海水成因的细— 粉晶白云岩相比, 云斑中Mg、Mn和Fe含量相对较低, 表明云斑形成于埋藏期海水的缓慢白云石化过程(王起琮等, 2016)。综合上述分析, 马四段的豹皮状碳酸盐岩先后经历了准同生白云岩化和埋藏白云岩化过程。
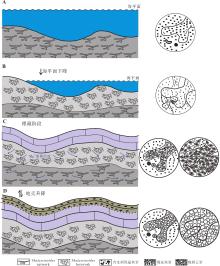
马四段向上变浅的岩性序列由下向上表现为从豹皮灰岩到豹斑云岩, 反映了由水平方向的潜穴过渡至三维展布的垂直网络空间(图 8), 至马四段顶部豹斑云岩与石膏岩伴生在一起, 推测沉积水体向上暴露形成局限咸化环境, 受到早成岩期大气淡水的岩溶改造, 早期形成的生物潜穴具有优势运移通道, 优先形成了一系列沿生物潜穴的不规则岩溶组构(图 8)。在准同生期, 富Mg2+卤水优先渗入生物潜穴或岩溶裂隙发育的地区, 发生回流渗透白云石化作用, 形成的白云石晶粒逐渐增生形成了白云石的“ 雾心亮边” 结构(图 8)。在回流渗透作用下, 粉晶白云石“ 雾心亮边” 结构的出现标志着流体能量上不足以支撑白云石晶粒的继续增长, 成为白云石通常生长到粉晶级别即停止的原因之一(董小波和牛永斌, 2015)。
 | 图 8 鄂尔多斯盆地中奥陶统马家沟组四段豹皮灰岩与豹斑云岩的成因演化(由A至D)示意图Fig.8 Skethc map showing evolution model of leopard skin limestone and leopard spot dolostone of the Member 4 of Middle Ordovician Majiagou Formation in Ordos Basin |
在后期深埋过程中, 随着地温和压力的升高, 白云石晶粒发生重结晶增大, 尤其是靠近晶间孔或次生溶孔一侧的白云石晶面生长最为迅速, 因争抢空间使得白云石晶粒呈镶嵌式的接触, 白云石呈“ 浑圆形” 的细晶白云石(图 7-A; 图 8), 这些浑圆形的晶粒白云石更多地保存于Thalassinoides boxwork的生物潜穴中。相较于Thalassinoides network, Thalassinoides boxwork的孔隙更为发育, 流体的渗透性更高, 导致白云石增大效果更为明显(图 7; 图 8)。
综上所述, 鄂尔多斯奥陶系马家沟组四段的豹皮灰岩和豹斑云岩主要受到了固底底质条件下生物扰动、高频海平面升降和早期成岩作用的耦合控制。马四段由下向上的变浅序列、生物扰动由弱到强、Thalassinoides形态由水平迷宫向三维箱状结构变化相耦合, 潜穴内部的白云石形态特征先后经历了准同生和埋藏白云石化作用的叠加影响, 导致白云石晶体形态特征的异同。
1)鄂尔多斯盆地中奥陶统马家沟组四段共识别出遗迹化石7属8种, 分别是Thalssinoides network, Thalassinoides boxwork, Planolites isp., Palaeophycus tubularis, Phycodes isp., Chondrites isp., Skolithos isp., ?Zoophycos isp., 主要属于固结底质底质的Glossifungites(舌菌迹)遗迹相, 进一步划分为上部、中部和下部Glossifungites遗迹亚相。
2)依据生物扰动和生物潜穴形态特征和方解石/白云石相对含量, 提出了生物扰动成因的豹皮(斑)状碳酸盐岩分类, 主要分为豹皮灰岩和豹斑云岩, 分别对应Thalassinoides network(水平二维迷宫)和Thalassinoides boxwork(三维箱状结构)。
3)豹皮(斑)状碳酸盐岩发育受到生物扰动、高频层序旋回下的早期成岩作用的共同控制。固底底质条件下的Thalassinoides生物潜穴为富Mg2+卤水的白云石化流体提供了良好的运移通道, 使得潜穴内部优先白云石化作用; 不同类型的生物潜穴在准同生和埋藏白云岩化的作用下, 最终分别形成了豹皮灰岩和豹斑云岩。
(责任编辑 张西娟; 英文审校 刘贺娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|