



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
湖北三峡地区黄陵隆起北缘灯影组微生物碳酸盐岩类型及古环境演化*
[范承蒙1  , 杨海林
, 杨海林1 , 杨洋2 , 温汉捷2 , 樊海峰2 , 张洪杰2 , 李方会3 , 肖蛟龙3 , 周煜4 ]
, 杨海林, 杨洋, 温汉捷|
|

第一作者简介 范承蒙,男,1995年生,硕士,主要从事古生物学与地层学研究工作。E-mail: 1468536140@qq.com。
黄陵隆起北缘震旦系灯影组不仅记录了震旦纪晚期环境与生命的协同演化,而且大量发育微生物碳酸盐岩,成为研究新元古代—古生代转折期古环境的重要对象。文中通过对黄陵隆起北缘震旦系灯影组开展详细的野外勘查及室内岩相分析解释,识别出云质微生物岩、云质微礁岩、云质微屑岩等的9个岩石微类,以云质微生物岩为主。根据微生物碳酸盐岩类型及其组合关系分析,认为发育多种向上变浅的潮下带—潮间带—潮上带沉积序列以及若干颗粒滩、微生物丘构成的米级沉积旋回,并以此为基础详细刻画出4次海侵—海退次级旋回, 建立了黄陵隆起北缘震旦纪晚期以潮坪相、丘滩体、颗粒滩为典型沉积特征的浅水碳酸盐岩台地模式。该发育模式与现代微生物碳酸盐岩沉积模式相似,明显特征为凝块石形成于正常浪基面以下的潮下带低能环境,较深于云质层纹石和云质叠层石。综合微生物碳酸盐岩沉积特征、沉积组合特征及沉积相演化规律,认为该地区古环境演化主要受次级海平面波动影响。推测微生物碳酸盐岩的发育程度可能为新元古代氧化事件之后氧含量增高以及微生物大量发育的海洋岩石形成的响应,同时也可能与石板滩生物群的出现和以微生物为食的后生动物演化导致微生物含量减少密切相关。分析黄陵隆起北缘震旦系灯影组微生物碳酸盐岩发育类型,不仅可以约束震旦纪晚期全球海平面变化、古海洋环境演化,而且对于研究早期生命演化具有重要意义,同时可为下一步油气储集层的探索提供有力的依据。
, YANG Hailin, YANG Yang, WEN HanjieAbout the first author FAN Chengmeng,born in 1995,is mainly engaged in palaeontology and stratigraphy. E-mail: 1468536140@qq.com.
The Sinian Dengying Formation at the northern margin of the Huangling Uplift not only records the co-evolution of the environment and life during the late Sinian,but also contains a large number of calcimicroliths,which serve as important indicators for studying the paleoenvironment of the Neoproterozoic-Paleozoic transition. In this paper,based on the detailed field investigation and indoor lithofacies analysis and interpretation of the Sinian Dengying Formation along the northern margin of Huangling Uplift,nine types of sedimentary microfacies are identified,including dolomitic microbialite,dolomitic microlithite and dolomitic microallolite,with dolomitic microbialite being the most dominant type. Based on the analysis of calcimicrolith lithology types and their combinatory relationships,it is proposed that a variety of subtidal-intertidal-supertidal stratigraphic sequences have developed,characterized by upward stacking and meter-scale stratigraphic cycles composed of multiple grain beaches and microbial mounds. On this basis,four transgression-regression secondary cycles are described in detail. A shallow water carbonate platform model,exhibiting typical lithofacies characteristics of tidal flat facies,mound beach bodies,and grain beaches during the late Sinian period at the northern margin of the Huangling Uplift,has been established. This carbonate sedimentary model is similar to that of modern microbial carbonate,with a notable characteristic being that the thrombolites are formed in a low-energy subtidal environment,situated below the normal wave base and deeper than dolomitic laminated stone and dolomitic stromatolites. Based on the lithofacies characteristics,lithofacies assemblage,and the evolution of sedimentary facies of calcimicrolith,it is inferred that the palaeoenvironmental evolution in this area is primarily influenced by secondary fluctuations in sea level. The development of calcimicrolith in the Sinian Dengying Formation,located at the northern margin of the Huangling uplift,may respond to the increased oxygen content and the formation of marine rocks abundant in microorganisms following the Neoproterozoic oxidation event. Additionally,it may be closely associated with the emergence of the Shibantan biota and the decrease of microbial content due to the evolution of metazoans. Analyzing the types of calcimicrolith development in the Sinian Dengying Formation at the northern margin of Huangling uplift not only constrains global sea-level changes and the evolution of paleo-ocean environment during the late Sinian,but also holds significant implications for studying the early life evolution. The Dengying Formation is characterized by a high abundance of calcimicrolith,which may provide a substantial basis for future exploration of oil and gas reservoirs.
最早识别出来的微生物岩类型是叠层石(Kalkowsky, 1908), 而微生物岩的第2种类型— — 凝块石, 直到1967年才被识别出来(Aitken, 1967)。“ 微生物岩” 这一术语是由Burne和Moore(1987)提出的, 用以指底栖微生物群落主导形成的碳酸盐沉积, 包括叠层石、凝块石、核形石、隐微生物岩和球形微生物岩。Riding(2000), Riding等(2011)将微生物碳酸盐岩分为叠层石、凝块石、树形石和均一石。和微生物相关的岩石是一个较为庞大的岩石群体, 除了微生物岩之外, 一些由微生物形成的岩石尚未被包括到现有的生物礁岩分类系统或微生物岩分类系统中(Folk, 1959, 1962; Dunham, 1962; Embry and Klovan, 1971; Wright, 1992; Riding, 2000; 梅冥相; 2007; 吴亚生, 2023)。Wu(2022a)将生物成因的碳酸盐岩以及生物成因的其他岩类命名为生物岩, 建立了包括所有生物岩的分类系统, 首次对微生物岩与微生物岩石这2个术语明确加以区分, 将微生物碳酸盐岩分为微生物岩、微礁岩和微屑岩, 其中微礁岩和微屑岩2个术语均为首次定义, 并将微生物岩分为叠层石、层纹石、包壳石、核形石和凝块石(Wu, 2022b; 吴亚生, 2023)。元古宙为微生物碳酸盐岩发育的鼎盛时期(Riding, 2000, 2006), 分析微生物碳酸盐岩可揭示该时期海洋环境和生物随时间发生的主要变化(McMenamin, 2004; Allwood et al., 2007)。微生物碳酸盐岩在指示元古代微生物群落构成、形成环境、古海水化学性质等方面具有重要作用, 是研究古生态和古环境的重要材料(Riding, 2000; Webb and Kamber, 2000; Dupraz et al., 2009)。震旦系灯影组不仅是新元古代氧化事件之后全球氧含量增高的海洋岩石方面的响应, 而且记录了早寒武世骨骼动物大爆发前夕地球早期动物的演化过程(Allwood et al., 2007; Zhu et al., 2007; Zhang et al., 2008; Xiao et al., 2016), 同时大量发育的微生物碳酸盐岩也为古环境演化提供了难得的证据(吴亚生等, 2007, 2020; Lee et al., 2015)。
针对微生物碳酸盐岩的研究目前主要集中于四川盆地和塔里木盆地, 前人对这2个地区的微生物碳酸盐岩岩石学特征及成因(王自强等, 2002; 陈寿铭等, 2013; 林孝先等, 2018)、微相类型和古地理演化(孙海涛等, 2018; 李新晨, 2020; 汪泽成等, 2020)等均开展了较为深入的研究, 对其剖面岩石序列特征及形成环境演化亦有报道(郭成贤和胡明毅, 1999; 李忠雄等, 2004; 周雁等, 2004; Zhu et al., 2007; Jiang et al., 2011; 汪泽成等, 2020), 这为分析其古环境演化奠定了良好的基础。但由于各个地区岩石特征、形成环境存在差异以及研究方法、研究程度的不同, 因此有关微生物碳酸盐岩的微观特征识别及命名还存在较大争议, 尚未达成共识; 况且已有的研究区过于集中, 而黄陵隆起北缘微生物碳酸盐岩研究相对滞后, 缺乏对该地区微生物碳酸盐岩形成环境演化的细致分析和海平面变化的精细刻画。
文中以湖北三峡地区黄陵隆起北缘保康县竹园沟ZK4803钻孔为研究对象。该钻孔震旦系灯影组地层连续, 界限清楚, 厚度可达520 m, 且地层记录完整丰富, 微生物碳酸盐岩特征明显, 是研究该时期古环境演化的难得材料。通过对该钻孔灯影组详细的室内岩相分析和解释, 首次从微生物碳酸盐岩角度刻画了其详细的古环境演化以及海平面变化历程, 并建立了相模式。该研究不仅对深入探索震旦纪晚期微生物岩成因具有一定的理论意义, 而且对进一步明确震旦纪灯影组沉积时期的古环境演化提供了有力的依据。
中扬子区北邻秦岭造山带, 南依江南造山带, 西接四川盆地, 向东收敛于湖北武汉以东(胡军等, 2021)。晋宁— 四堡造山运动形成扬子克拉通统基底(张国伟等, 2013)。在Rodinia超大陆裂解的全球构造背景下, 以扬子克拉通东南缘为中心形成南华裂谷盆地, 伴有裂谷型岩浆活动, 发育一套火山岩层系(王剑, 2000)。南华冰期阶段(635~720 Ma), 发育持续伸展构造背景下的寒冷气候地层, 这也是扬子新元古代— 早古生代海相盆地被动陆缘阶段早期的碎屑岩陆架建设过程(汪正江等, 2015)。震旦纪以来, 扬子地块进入台地稳定发展期, 受超大陆裂解-聚合旋回影响, 表现为克拉通内部稳定、外缘活动的特点(童崇光, 1992; Wang and Li, 2003; 魏国齐等, 2015; 李智武等, 2019)。陡山沱期, 气候转暖, 扬子克拉通经历快速海侵, 岩石类型以黑色页岩夹碳酸盐岩、硅质岩和磷块岩为主(Jiang et al., 2011)。灯影组是继陡山沱组之后发育的一套完整的海侵退积式地层序列, 是在黄陵隆起北缘分布广泛的古老碳酸盐岩层系, 具有良好的油气地质条件(杜金虎等, 2015)。
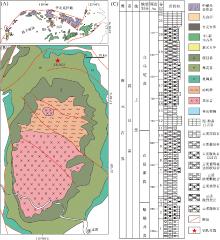
黄陵隆起北缘位于中扬子板块北部区域(图 1-A, 1-B)。与上扬子板块一样, 震旦系灯影组均为浅水碳酸盐岩台地相地层, 不同的是中扬子板块以开阔台地— 局限台地沉积为普遍特征, 而上扬子板块大部分地区为潮坪— 潟湖沉积, 因此, 中扬子板块震旦纪灯影期沉积水体较深(马永生等, 2009)。受全球海平面变化的影响, 两者均可识别出4个三级层序(周雁等, 2004)。在地层发育特征上, 黄陵隆起北缘震旦系灯影组自下而上为蛤蟆井段、石板滩段和白马沱段, 地层厚度约520 m。蛤蟆井段主要为一套云质砂屑颗粒岩, 为海侵序列沉积; 石板滩段沉积期, 海水后退, 以发育大套云质微生物岩地层为主; 白马沱段古环境较为复杂, 发育有海退初期的云质凝块石和云质微骨架岩, 海退之后可见以云质层纹石和泥-粉晶云岩为主的地层(图 1-C)。综上所述, 黄陵隆起北缘震旦系灯影组是发育在陡山沱组之上的一套以微生物碳酸盐岩为主的浅水碳酸盐岩台地沉积。
 | 图 1 黄陵隆起北缘地质图及湖北保康县竹园沟ZK4803钻孔柱状图 A, B— 黄陵隆起北缘地质图(据Zhao et al., 2012; Wang et al., 2021); C— 湖北保康县竹园沟ZK4803钻孔柱状图Fig.1 Geological map of northern margin of Huangling Uplift and histogram of ZK4803 borehole in Zhuyuangou, Baokang County of Hubei Province |
对于微生物碳酸盐岩分类, 学者们已提出了多种分类方案(Burne and Moore 1987; Riding, 2000; 梅冥相, 2007), 但由于分类依据不同, 造成微生物碳酸盐岩的分类方案具有较大分歧。Wu(2022b)建立一个包括所有生物岩的较为完善的新分类系统, 其不仅包括了传统的生物礁岩和微生物岩类型, 并且对前人分类系统中尚未包括的生物岩进行定义和命名。新分类系统可以避免以往一种岩石被不同学者归入不同类型的问题, 从而使今后的生物岩研究有了更科学的依据。该分类系统首次定义的微礁岩和微屑岩在本次研究中均有发现。文中在Wu(2022b)分类方案和命名的基础上, 结合黄陵隆起北缘保康县竹园沟ZK4803钻孔震旦系灯影组岩石的微观特征, 共识别出岩石类型6大类, 并细分为10种微类(表 1)。
| 表 1 黄陵隆起北缘灯影组主要岩石类型 Table1 Main rock types of the Dengying Formation in northern margin of Huangling Uplift |
1)云质叠层石。叠层石是被最早识别出来的微生物岩类型, 也是最容易识别的微生物岩类型(Kalkowsky, 1908)。研究区叠层石以微生物席成因的纹层结构为特征, 纹层明暗相间呈近平直状, 延伸距离较短(图 2-A, 2-B)。云质叠层石在灯影组中发育较少, 仅在白马沱段顶部有不足10 m厚的出露。由于重结晶作用, 纹层结构已遭受到很大程度的破坏。
 | 图 2 黄陵隆起北缘灯影组微生物碳酸盐岩显微特征(一) A— 白马沱段, 云质丘状叠层石; B— A中黄框放大2倍, 可隐约看到向上叠覆的纹层结构; C— 白马沱段, 云质层纹石, 泥晶纹层(la)与亮晶纹层(gl)组成的纹层结构; D— 白马沱段, 云质层纹石, 泥晶纹层(la)与亮晶纹层(gl)组成的纹层结构; E— 蛤蟆井段, 云质层纹石, 相互缠绕的泥晶纹层(la)与微亮晶纹层(mgl)组成的纹层结构; F— 白马沱段, 云质斑点状凝块石, 暗色斑点状凝块(sd)分布于微亮晶沉积物(mgl)中, 粒间孔可见粉晶白云石(po)填充; G— 白马沱段, 云质网状凝块石, 网线由暗色泥晶白云石组成, 网眼为粉晶白云石充填; H— 石板滩段, 云质网状凝块石, 微生物诱导矿化形成的致密泥晶, 泥晶中可见微生物留下的模孔被亮晶充填(红色虚线), 泥晶周围为亮晶充填体(黄色虚线); I— 石板滩段, 云质网状凝块石, H中黄框放大2倍, 可见由致密黑色泥晶(mu)组成的凝块结构; J— 白马沱段, 云质凝块石— 层纹石, 泥晶纹层(la)、网状凝块石(net)组成的纹层结构; K— 白马沱段, 云质凝块石— 层纹石, H中黄框放大2倍, 泥晶纹层(la)、网状凝块石(net)组成的纹层结构; L— 白马沱段, 云质凝块石— 层纹石, 泥晶纹层(la)、网状凝块石 (net)、斑点状凝块石(sd)组成的纹层结构。所有照片均为单偏光Fig.2 Microscopic characteristics of calcimicrolith of the Dengying Formation in northern margin of Huangling Uplift(Ⅰ) |
2)云质层纹石。指微生物席形成的层状碳酸盐岩(Riding, 2000; 梅冥相, 2007; 吴亚生等, 2018), 表现为微生物席形成的暗色致密泥晶纹层与其他成分组成的纹层交替出现。在显微镜下, 研究区云质层纹石的暗色致密泥晶纹层一般呈黑色, 厚度变化较大, 最小厚度为数微米, 最大厚度可达0.5 mm左右。其他成分的纹层颜色较浅, 矿物组成变化较大, 可为较大的白云石颗粒形成的亮晶或微亮晶纹层、凝块石纹层、颗粒纹层或其他沉积物形成的纹层, 甚至可见2种或2种以上沉积物组成的复合纹层(图 2-C, 2-D, 2-E)。层纹石与叠层石的区别在于前者呈层状, 后者呈柱状、丘状、分枝状等形状, 且前者的纹层延伸距离较长。
3)云质凝块石。凝块石被定义为具有凝块结构的微生物岩(Aitken, 1967; Riding et al., 2011; 吴亚生等, 2018)。显微镜下, 研究区云质凝块石为呈网状(图 2-G, 2-H, 2-I)、斑点状(图 2-F)等形态的暗色泥晶凝块。网状泥晶凝块中分布有近圆形的亮晶充填体, 具有明显的泥晶边缘, 推测其可能为微生物死亡之后被降解留下的模孔(图 2-H, 2-I)又被亮晶所胶结, 泥晶边缘为微生物矿化形成的矿物壳, 泥晶凝块之间为成岩作用过程中形成的亮晶胶结物。斑点状凝块石由小于1 μ m的斑点状泥晶凝块和周围的微亮晶组成。
4)云质凝块石— 层纹石。是由凝块石纹层和微生物席形成的泥晶纹层共同组成的一种纹层结构, 同时具有凝块石和层纹石的特征。显微镜下, 可见层纹石中形成的泥晶纹层以及泥晶纹层之间的凝块石纹层, 其中凝块石纹层由网状凝块或斑点状凝块组成, 凝块之间的矿物多为微亮晶(图 2-J, 2-K, 2-L)。
5)云质葡萄花边状胶结岩。岩石后期遭受溶蚀作用形成较多孔洞, 孔洞中充填胶结物, 当胶结物的体积超过岩石体积50%, 称为胶结岩(Wright, 1992)。研究区该类岩石的胶结物以葡萄花边状构造为特征(图 3-A), 原岩为云质凝块石。葡萄花边状构造包括核心和包壳两部分(图 3-B, 3-C), 前者即凝块石, 主要为微生物作用形成的碳酸盐矿物, 后者主要是在生长— 同生阶段由海水无机结晶作用形成(林孝先等, 2018)。
 | 图 3 黄陵隆起北缘灯影组微生物碳酸盐岩显微特征(二) A— 石板滩段, 云质葡萄花边状胶结岩, 核心为云质网状凝块石(co), 包壳(cla)为多期次胶结作用形成的同心纹层; B— 石板滩段, 云质葡萄花边状胶结岩, 核心为黑色泥晶白云石(co), 包壳(cla)为胶结作用形成的同心纹层; C— 石板滩段, 云质葡萄花边状胶结岩, 核心为黑色泥晶白云石(co), 包壳(cla)为胶结作用形成的同心纹层; D— 石板滩段, 云质砂屑颗粒岩, 砂屑颗粒(sc)内部重结晶作用明显, 仅留下外围的泥晶边缘; E— 石板滩段, 云质砂屑颗粒岩, 成岩作用导致砂屑颗粒(sc)呈扁平状, 顺层分布, 颗粒内部重结晶作用明显, 仅留下外围的泥晶边缘; F— 蛤蟆井段, 云质砂屑颗粒岩, 砂屑颗粒(sc)由于重结晶作用形成粉晶白云石; G, H, I— 蛤蟆井段, 云质核形石, 核心为泥晶、亮晶或碎屑颗粒组成, 包壳为暗色泥晶和亮晶交替组成的同心纹层; J— 白马沱段, 云质附枝菌微骨架岩, 疑似附枝菌(Epip.)原地钙化形成的黑色泥晶白云石; K— 白马沱段, 云质肾形菌微骨架岩, 疑似肾形菌(Rena.)原位钙化形成的黑色泥晶白云石, 周围为微生物作用形成的组构; L— 白马沱段, 云质肾形菌微骨架岩, K中黄框放大2倍, 疑似肾形菌(Rena.)原位钙化形成的黑色泥晶白云石。所有照片均为单偏光Fig.3 Microscopic characteristics of calcimicrolith of the Dengying Formation in northern margin of Huangling Uplift(Ⅱ) |
6)云质砂屑颗粒岩。研究区的云质砂屑颗粒岩中砂屑颗粒形态清晰, 由暗色泥粉晶白云石构成的椭圆形、近圆形颗粒组成, 粒径在0.1~1.2 mm之间。颗粒由波浪作用冲刷而成, 分选性和磨圆度均较好, 具有明显光滑的颗粒边缘, 呈孤立状或线接触。由于重结晶作用而呈残余的砂屑结构, 大部分颗粒内部重结晶作用后变成粉晶白云石, 仅剩黑色泥晶轮廓的残余, 颗粒间主要为亮晶胶结物(图 3-D, 3-E, 3-F)。
7)云质核形石。具有微观同心纹层的球形结构(Burne and Moore, 1987; 吴亚生等, 2018), 是由微生物形成的泥晶纹层以同心纹层状包裹在某种核心之上(如岩石碎屑、半固结沉积物的碎屑、生物骨骼的碎屑等)而形成的。黄陵隆起北缘灯影组核形石以球形为主, 少数呈椭球形、纺锤形等不规则形态, 直径0.1~5 mm, 少数直径可达20 mm, 多集中于0.2~1 mm之间。核形石颗粒间为亮晶胶结, 颗粒内部可能遭受重结晶作用, 部分同心纹层受到破坏(图 3-G, 3-H, 3-I)。
8)云质微骨架岩。是具有底栖微生物的原地骨骼构成的骨架(Wu, 2022b)。研究区微骨架岩受成岩作用影响, 微生物化石形态保存不是太好, 已识别出疑似附枝菌、肾形菌化石。构成此处岩石骨架的附枝菌化石由树枝状、短放射状的泥晶白云石构成(图 3-J), 肾形菌化石为由一系列泥晶白云石构成的中空球状体— 肾状体紧密排列组成的分枝状结构, 内部的花朵状空腔被粗晶白云石所充填(图 3-K, 3-L)。云质微骨架岩中微体化石间多为微亮晶充填, 少见亮晶胶结物。云质微骨架岩发育于白马沱段中部。
微生物碳酸盐岩主要分布于海洋环境中。自~3400 Ma的叠层石开始发育(Wacey, 2010), 其一直持续至今, 可作为探索地球上古老生命活动的直接证据(Riding, 2000, 2006), 被广泛用于追踪海洋环境生命活动、水化学性质和水动力条件等古环境因素, 以及研究地质历史时期生物与环境的协同演化(Allwood et al., 2007; Lee et al., 2015)。通过对黄陵隆起北缘ZK4803钻孔微生物碳酸盐岩类型划分以及显微特征的观察, 识别出各类岩石发育的沉积环境, 以此分析纵向上沉积相、典型沉积序列及旋回的变化特征, 建立沉积演化和海平面变化趋势(图 4)。
 | 图 4 黄陵隆起北缘ZK4803钻孔灯影组沉积演化综合地层柱状图Fig.4 Comprehensive stratigraphic histogram of the Dengying Formation at ZK4803 borehole in northern margin of Huangling Uplift |
通过对黄陵隆起北缘震旦系灯影组垂向上高精度的岩石显微图像分析, 基于微生物碳酸盐岩对古环境的响应, 并结合微生物碳酸盐岩中胶结物的类型, 识别出各类岩石形成的沉积环境(表 2)。
| 表 2 黄陵隆起北缘ZK4803钻孔灯影组各类岩石沉积环境 Table2 Sedimentary environment of various rocks in the Dengying Formation at ZK4803 borehole in northern margin of Huangling Uplift |
1)云质凝块石。由于纹层缺失等原因, 因此认为其发育的环境不应局限于潮间带, 而是自潮间带下部到深处或更深处, 水动力由强到弱, 均可能有分布(Aitken, 1967)。研究区云质凝块石内部矿物成分分布均匀, 应形成于潮下带低能环境。关于网状凝块石中模孔以及泥晶凝块之间的亮晶充填体, 可能为成岩作用过程中流体由下往上运移过程中结晶的产物。斑点状凝块石中的微亮晶可能为泥晶重结晶形成(吴亚生等, 2018)。
2)云质微骨架岩。作为微生物群落原地钙化的产物, 微生物化石周围被微亮晶填充, 表明其形成于水动力较强的环境。因上、下层位发育有云质凝块石, 故认为它们形成于正常浪基面以上、水体能量较高的潮下带弱动荡环境。
3)云质砂屑颗粒岩。发育在蛤蟆井段底部和石板滩段中部, 颗粒的成因暂未确定, 共同特征为颗粒外围均见泥晶边缘。推测砂屑颗粒为云质微生物岩破碎、受波浪作用冲刷形成。云质砂屑颗粒岩通常形成于浪基面以上、强水动力条件的潮下带中等动荡环境。
4)云质核形石。该类岩石的形态和结构在一定程度上取决于其生长过程中的水流能量(Dahanayake, 1997; 张园园等, 2009; 齐永安等, 2016)。研究表明, 海平面、水动力条件等变化会在核形石包壳和核心有不同的显示, 故可以通过核形石核心、包壳、形状以及胶结物类型等的差异来判断其形成的沉积环境(Vé drine et al., 2007)。黄陵隆起北缘灯影组核形石形态各异, 核心多为泥晶矿物, 包壳厚度较小, 核形石颗粒之间主要为亮晶胶结, 可见其是经过水流作用的冲洗将颗粒间的物质带走, 再经化学沉淀形成, 说明核形石形成于动荡或连续水流、水体能量较高的潮下带强动荡环境。
5)云质葡萄花边状胶结岩。仅在蛤蟆井段顶部和石板滩段顶部发育, 代表微生物丘向上变浅暴露发生准同生期大气淡水淋滤改造, 溶蚀孔洞被大气淡水和海水的多期胶结物充填。可能形成于潮间带暴露环境。
6)云质凝块石— 层纹石。与云质层纹石和云质凝块石不同, 其可能形成于不同的古环境条件中, 是海平面频繁升降的产物。为了刻画黄陵隆起北缘灯影期精细的古环境演化及海平面变化, 笔者将云质凝块石— 层纹石作为一种单独的岩石类型, 认为其通常形成于潮间带环境。
7)云质叠层石。该类岩石的形态结构与古环境的水动力条件密切相关(Logan et al., 1964; 钱迈平等, 2002; 范正秀等, 2018)。研究区以小型平直状叠层石为特征, 纹层形态较为均一, 规模不大, 指示水体循环受限、相对低能潮间带局限环境。
8)云质层纹石。可能形成于一种低能的环境(Wright and Barnett, 2015)。研究区组成层纹石的矿物成分较为复杂, 一般由泥晶纹层、亮晶纹层和微亮晶纹层构成, 在潮坪环境中均有分布。
9)泥— 粉晶白云岩。以发育鸟眼孔和裂缝为特征, 作为暴露于大气环境的标志, 形成于潮上带蒸发环境。
通过对黄陵隆起北缘震旦系灯影组垂向上岩石发育沉积环境的分析, 认为其相类型主要以微生物丘、颗粒滩、潮下带以及潮坪沉积为主。
微生物丘纵向上可分为丘基、丘核、丘坪3种沉积微相。丘基位于微生物丘底部, 是微生物发展的初期和拓殖阶段, 微生物较缺乏, 通常代表相对海平面较低、水动力较小的环境, 以发育云质层纹石为主, 是初始海平面扰动较小背景下的产物; 丘核为平均海平面以下的浅水潮下带沉积, 主要由高能潮下带发育的云质砂屑颗粒岩或低能潮下带发育的云质凝块石组成, 以垂向加积为主, 进入微生物发展的泛殖阶段; 丘坪为微生物丘快速“ 生长” 、海平面相对下降背景下暴露于水面之上的产物, 由于准同生期大气淡水淋滤改造, 溶蚀孔洞被成岩环境下大气淡水和海水的多期胶结物充填形成云质胶结岩。
颗粒滩主要由高能潮下带沉积产物组成, 沉积梯度在正常浪基面与平均低潮面之间, 水体持续动荡。颗粒滩可细分为由云质砂屑颗粒岩组成的砂屑滩沉积微相和云质核形石组成的核形石滩沉积微相, 砂屑颗粒和核形石颗粒之间均为亮晶胶结物, 受波浪作用影响颗粒分选性和磨圆度较好, 显示出高能、动荡的环境。
潮下带可细分为高能潮下带和低能潮下带2种沉积微相。高能潮下带除发育颗粒滩外, 还发育微生物体相互缠绕形成的具有一定抗浪能力的微骨架岩。低能潮下带水体循环良好、能量较低、盐度正常, 通常为底栖微生物群落大量发育的区域, 以形成云质凝块石为主要特征。
潮坪相可进一步细分为潮间带暴露环境、潮间带局限环境和潮上带蒸发环境3种沉积微相。潮坪相作为藻席生态系统的聚集地, 以形成具有纹层结构微生物碳酸盐岩为主要特征, 包括云质层纹石和云质叠层石等。泥— 粉晶白云岩中发育鸟眼孔及裂缝, 指示潮上带蒸发环境。
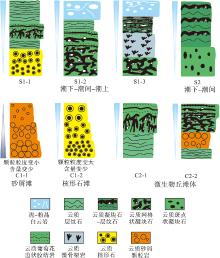
研究发现, 黄陵隆起北缘震旦纪灯影期发育若干向上变浅的沉积序列以及米级沉积旋回, 其中沉积序列表现为相对海平面的逐渐降低, 其形成可能是由于海平面下降或沉积速率大大超过沉降速率所致, 而沉积旋回反映水体深度波动较小。沉积序列和沉积旋回共有4种主要类型: 序列类型Ⅰ (S1), 潮下带— 潮间带— 潮上带序列; 序列类型Ⅱ (S2), 潮下带— 潮间带序列(图 5); 旋回类型Ⅰ (C1), 为潮下带颗粒滩旋回; 旋回类型Ⅱ (C2), 为潮间带微生物丘旋回(图 5)。
 | 图 5 黄陵隆起北缘ZK4803钻孔灯影组典型沉积序列及旋回演化示意图Fig.5 Typical sedimentary sequence and cycle evolution diagram of the Dengying Formation at ZK4803 borehole in northern margin of Huangling Uplift |
黄陵隆起北缘震旦系灯影组岩石类型受潮汐、波浪等作用强烈控制, 古环境演化受全球海平面波动、区域构造活动以及碳酸盐岩形成速率综合影响。该时期研究区共经历了4次海侵— 海退次级旋回(图 4)。
蛤蟆井段主要发育云质核形石和云质砂屑颗粒岩, 指示水动力较高的潮下颗粒滩环境, 作为灯影组最底部地层, 继承了陡山沱末期海侵相序的特征。石板滩段由2期连续的微生物丘滩体构成, 以微生物丘内发育颗粒滩微相形成的云质砂屑颗粒岩为主要特征:第1期微生物丘沉积厚度较小, 仅20 m左右, 底部丘基主要为云质层纹石, 丘核主要为云质凝块石, 顶部丘坪主要为古岩溶系统改造形成的云质葡萄花边状胶结岩; 第2期微生物丘滩体沉积厚度较大, 约150 m, 发育模式与第1期相似, 仅有的区别为丘核中发育1层颗粒滩云质砂屑颗粒岩, 指示了由微生物丘滩体主导的潮间带— 潮下带环境。白马沱段岩性较为复杂, 除云质砂屑颗粒岩和云质核形石以外, 其他的岩石类型在该段均有分布, 指示了水体动荡、海平面变化相对较大的沉积环境。
次级旋回Ⅰ 由整个蛤蟆井段和石板滩段底部构成, 厚度约150 m, 岩相序列为向上颗粒粒度变大的颗粒滩(海侵)、微生物丘(海退)、潮上带云质层纹石(海退)。次级旋回Ⅱ 由石板滩段顶部和白马沱段底部构成, 厚度约150 m, 岩相序列为低能潮下带云质凝块石(海侵)、微生物丘(海退)、潮上带蒸发环境泥— 粉晶白云岩(海退)。次级旋回Ⅲ 由白马沱段中部构成, 厚度约50 m, 岩相序列为潮下低能带云质凝块石(海侵)、潮间带云质凝块石— 层纹石(海退)。次级旋回Ⅳ 由白马沱段顶部构成, 厚度约150 m, 岩相序列为潮下低能带云质凝块石(海侵)、潮间带云质凝块石— 层纹石(海退)、潮上带蒸发环境泥— 粉晶白云岩(海退)。
灯影组为元古代— 古生代转折期的地层, 体现了后生动物即将出现之前以微生物主导的海洋生态系统中的碳酸盐岩岩相特征(Riding, 2006; Riding and Virgone, 2020), 该时期海洋以潮汐作用和波浪作用为主。根据对黄陵隆起北缘震旦系灯影组微生物碳酸盐岩特征、岩相演化规律的研究以及在前人相关研究成果的基础上, 建立了黄陵隆起北缘震旦系灯影组的浅水碳酸盐岩台地沉积模式。该模式依据海水深度以及波浪、潮汐作用影响的范围, 基于微生物碳酸盐岩对古环境的响应, 将其形成环境划分为潮下带、潮间频繁暴露环境、潮间局限环境(弱暴露)以及潮上带蒸发环境(图 6)。
 | 图 6 黄陵隆起北缘灯影组沉积模式Fig.6 Sedimentary model of the Dengying Formation in northern margin of Huangling Uplift |
针对微生物碳酸盐岩的沉积模式, 国内外学者已经开展过大量研究(朱士兴, 1993; Jahnert and Collins, 2012), 部分学者认为现代微生物碳酸盐岩发育环境与元古宙有较大的差别(Lyons et al., 2009), 故认为用“ 将今论古” 的思维将现代沉积模式沿用于前寒武纪古老深层微生物碳酸盐岩似乎并不合适。但是笔者通过对黄陵隆起北缘ZK4803钻孔震旦系灯影组岩石显微镜下观察, 认为晚震旦世微生物碳酸盐岩的沉积模式与现代基本吻合, 尤其是云质叠层石、云质层纹石和云质凝块石的发育环境, 凝块石主要形成于潮下带低能环境(Aitken, 1967), 形成时水体深度较云质叠层石和云质层纹石大。因此, 笔者认为古代与现代微生物碳酸盐岩沉积模式的对比不能简单地从单一变量入手, 应综合考虑大气环境、海水水化学条件、生物等相关因素, 建立全面的沉积模式。
关于黄陵隆起北缘震旦纪末期古地理特征, 笔者从微生物碳酸盐岩对古环境响应的角度, 认为震旦系灯影组在黄陵隆起北缘表现为以丘滩体、颗粒滩、潮坪相为典型岩相的受次级海平面波动影响的浅水微生物碳酸盐岩台地, 构成4次由快速的海侵相序到缓慢的海退相序的次级旋回。岩石类型受潮汐作用和波浪作用的控制, 以大规模发育微生物碳酸盐岩为特征。微生物碳酸盐岩作为一类重要的储集层, 已被国内外油气勘探实践证实, 黄陵隆起北缘震旦系灯影组含有大量的微生物碳酸盐岩, 对其进行研究可能为下一步油气储集层的探索提供有力的依据。
微生物碳酸盐岩作为微生物生命活动的产物, 其形成直接与微生物在地质历史时期的丰度紧密相关。蛤蟆井段、石板滩段以及白马沱段底部以发育大量的微生物碳酸盐岩为特征, 表明该时期海洋环境适宜于微生物的生长发育并且达到了较高的丰度, 可为晚新元古代的海洋含氧量的演化提供信息。石板滩段较灯影组其他地层微生物碳酸盐岩发育程度高, 可能与石板滩生物群的出现有密切的关系。石板滩生物群以典型的埃迪卡拉软躯体化石和管状化石为代表(Chen et al., 2014; Wang et al., 2020a; Xiao et al., 2021), 具起皱的微纹层是存在微生物席的证据(Chen et al., 2013; Meyer et al., 2014), 而微生物席被认为是埃迪卡拉类软躯体化石的重要保存因素和诱导碳酸盐岩矿物沉淀的主控因素之一(Callow and Brasier, 2009; Wang et al., 2020b), 因此, 石板滩生物群的出现可能为碳酸盐矿物沉淀提供了动力学基础。白马沱段顶部未见微生物碳酸盐岩发育, 其作为灯影期沉积的最后一个阶段, 是否是因为早期生命演化出现以微生物为食的后生动物, 从而导致微生物丰度的急剧减少, 还需要作进一步研究。总之, 通过对新元古代— 古生代转折期微生物碳酸盐岩的研究, 可以更加准确地推断地质历史时期的古环境信息, 为大气圈、水圈以及生命演化提供有力的信息。
1)根据不同微生物碳酸盐岩类型的组构特征差异以及分类原则, 将黄陵隆起北缘震旦系灯影组划分为9种岩石类型: 云质层纹石、云质叠层石、云质凝块石— 层纹石、云质葡萄花边状胶结岩、云质核形石、云质砂屑颗粒岩、云质微骨架岩、云质凝块石和泥— 粉晶白云岩。
2)基于岩石特征解释和纵向组合关系, 发现黄陵隆起北缘震旦系灯影组存在若干向上变浅的沉积序列及米级沉积旋回, 包括S1潮下带— 潮间带— 潮上带沉积序列、S2潮下带— 潮间带沉积序列以及C1颗粒滩沉积旋回、C2微生物丘沉积旋回等。
3)古环境演化研究表明, 灯影组经历了4次海侵— 海退旋回, 次级旋回Ⅰ 由整个蛤蟆井段和石板滩段底部构成, 次级旋回Ⅱ 由石板滩段顶部和白马沱段底部构成, 次级旋回Ⅲ 由白马沱段中部构成, 次级旋回Ⅳ 由白马沱段顶部构成。
4)灯影组形成环境受全球海平面波动、区域构造活动以及碳酸盐岩形成速率综合影响, 岩石类型由潮汐作用和波浪作用控制, 反映了一个以丘滩体、颗粒滩、潮坪相为典型沉积特征的浅水碳酸盐岩台地沉积模式。微生物碳酸盐岩发育模式与现代相似, 明显特征为凝块石形成于正常浪基面以下的潮下带低能环境, 对比云质叠层石和云质层纹石, 云质凝块石形成时的水体较深。
5)黄陵隆起北缘震旦系灯影组微生物碳酸盐岩大量发育, 可为下一步油气储集层的探索提供有力的依据。石板滩段微生物碳酸盐岩发育达到鼎盛, 可能与新元古代氧化事件、石板滩生物群以及早期后生动物演化密切相关。
(责任编辑 张西娟; 英文审校 刘贺娟)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|